
Khoikhoicetus kergueleni, Lambert & Muizon & Duhamel & Plicht, 2018
|
publication ID |
https://doi.org/ 10.5252/geodiversitas2018v40a6 |
|
publication LSID |
urn:lsid:zoobank.org:pub:06EB756D-EE16-4B28-A09C-EA983B758397 |
|
persistent identifier |
https://treatment.plazi.org/id/2543332B-3F91-4890-8A85-93EB8A3B11E0 |
|
taxon LSID |
lsid:zoobank.org:act:2543332B-3F91-4890-8A85-93EB8A3B11E0 |
|
treatment provided by |
Marcus |
|
scientific name |
Khoikhoicetus kergueleni |
| status |
sp. nov. |
Khoikhoicetus kergueleni n. sp.
urn:lsid:zoobank.org:act:2543332B-3F91-4890-8A85-93EB8A3B11E0
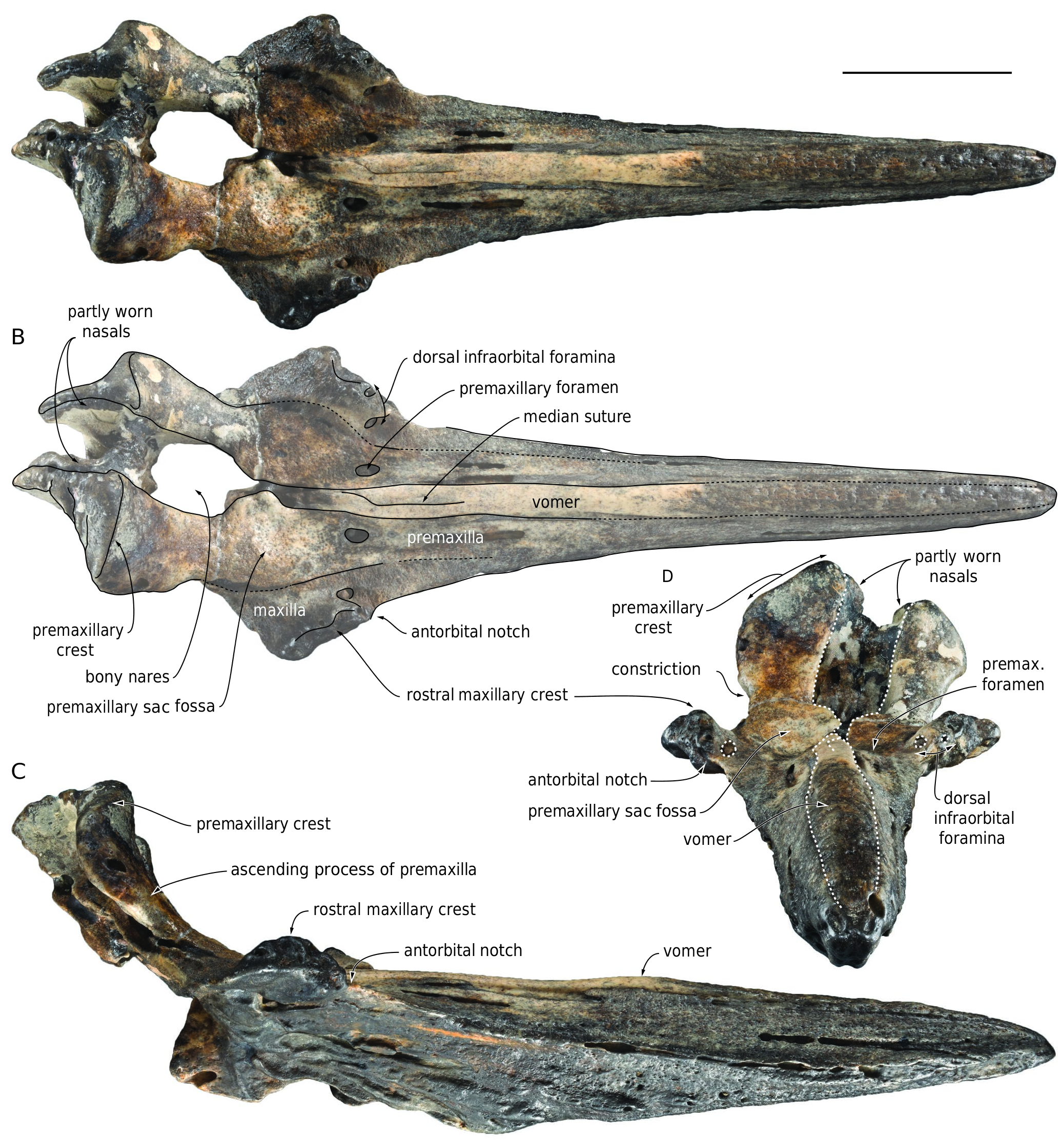
HOLOTYPE. — MNHN.F.COI1, partial cranium including the complete rostrum the anterior part of the cranium with a large part of the face, premaxillary sac fossae, the bony nares, part of both supraorbital regions, and the vertex ( Fig. 2 View FIG ).
TYPE LOCALITY. — The holotype has been fished on Skiff Bank , 370 km SWW to Kerguelen Islands at a depth of 885 m ; geographic coordinates 49°49’17.4”S, 63°37’33.6”E ( Fig. 1 View FIG ).
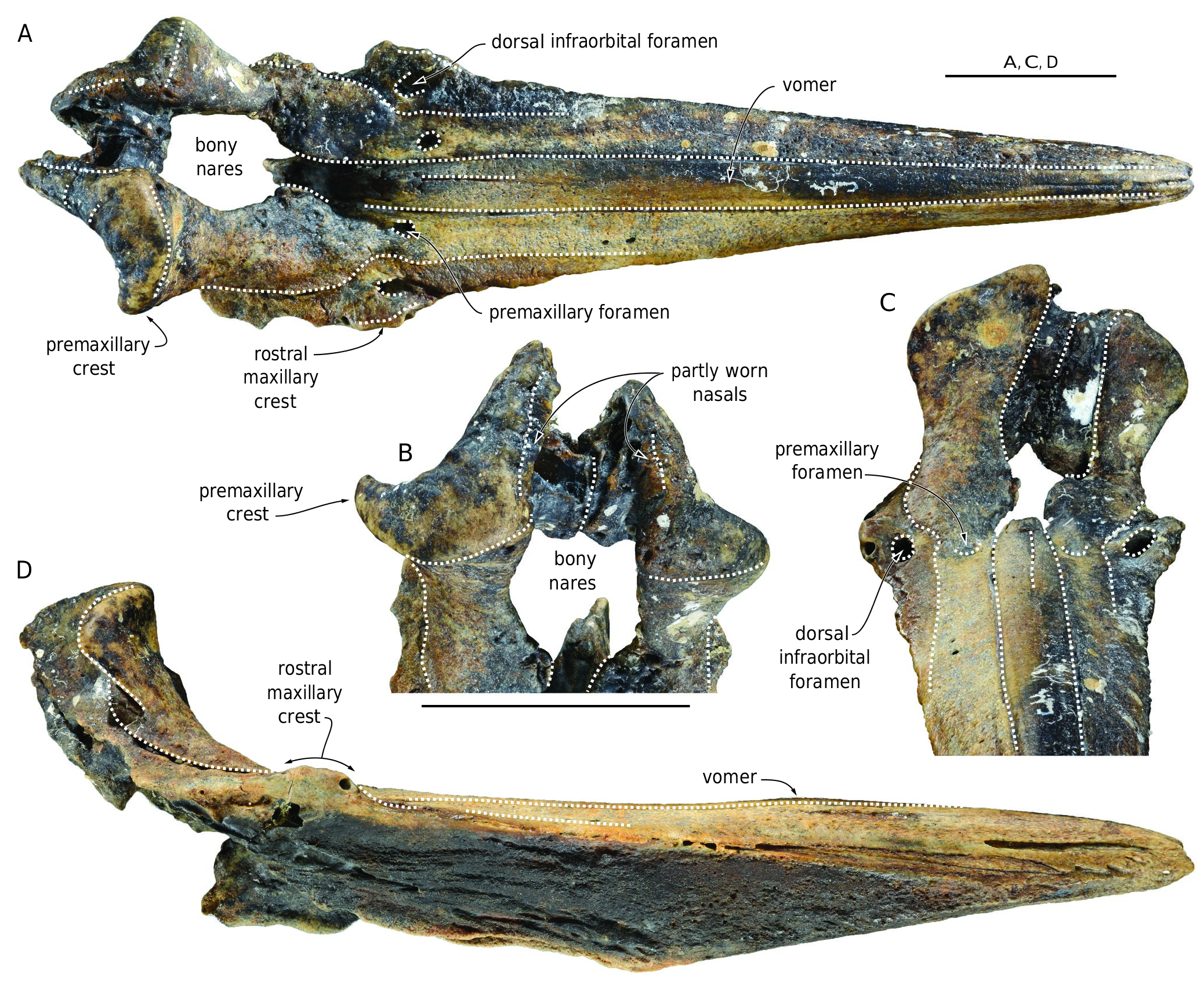
REFERRED SPECIMEN AND LOCALITY. — Partial cranium MNHN.F.COI13 including rostrum, premaxillary sac fossae, part of both supraorbital regions, and vertex ( Fig. 3 View FIG ); off Kerguelen Islands, no indication about the precise locality is available.
ETYMOLOGY. — In memory of Yves Joseph de Kerguelen de Trémarec (1734-1797), rear admiral of the French Royal Navy, who took possession of the “îles de la Désolation” (later designated by James Cook “Kerguelen Archipelago”) for the crown of France in 1772.
DIAGNOSIS. — Khoikhoicetus kergueleni n. sp. differs from Khoikhoicetus agulhasis in the following features: larger size; outline of the rostrum in dorsal view distinctly triangular with straight lateral edges; dorsal infraorbital foramen notably distant from the maxillapremaxilla suture (from 10 to 20 mm); premaxillary sac fossae signifi-
A
cantly proportionally narrower; bony nares relatively narrower and roughly oval-shaped; premaxillary crests anteroposteriorly thicker and wider (in anterior view) than the premaxillary sac fossae; and premaxillary crests located posterior to the premaxillary sac fossae (in lateral view), and not overhanging them.
BRIEF DESCRIPTION AND COMPARISON
Completely preserved, the rostrum of both specimens is higher than wide for most of its length. The lateral edges of the rostrum
are roughly straight in dorsal view and regularly diverge posteriorly until the antorbital notches.The rostrum of MNHN.F.COI13 is more massive and widens posteriorly more abruptly than on COI1. The mesorostral groove is completely filled by the vomer, which is dorsally higher than the adjoining premaxilla for its whole length. This condition is more accentuated in COI1, in which the vomer forms a small dome in the mid-length region of the rostrum. A median suture marks the posterior portion of the vomer. The rostral maxillary crest forms an elevated, but relatively short dome in the antorbital region. Located medial to the crest, the medium-sized main dorsal infraorbital foramen is opening anteriorly to anteromedially, roughly at the same anteroposterior level as the premaxillary foramen, but higher dorsally than the latter (best seen in anterior view of MNHN.F.COI1). In COI13 the main dorsal infraorbital foramen is two to three times larger than on the other specimen and opens slightly posterior to the premaxillary foramen. The premaxillary foramen is large, especially on COI13, in which it opens posteriorly and extends as a wide and deep groove running toward the anteromedial area of the premaxillary sac fossa. It is slightly posterior to the antorbital notch on both specimens. Anteriorly defining the bony nares, the medialmost angle of the right premaxillary sac fossa projects medially. A similar medial projection is observed on the left side, but it is less protruding and more rounded. The lateral margin of the ascending process of the premaxilla (sensu Bianucci et al. 2007) displays a marked constriction in anterior view. The medial margins of the right and left ascending processes converge slightly dorsally, and the posterodorsal part of the anterior surface is subvertical, but does not overhang the more ventral part of the ascending process. The markedly asymmetric premaxillary crests are directed slightly posterolaterally, and the dorsal margin of each crest slopes abruptly ventrolaterally in anterior view. Although a large part of each nasal is lost, the preserved lateralmost portions indicate transversely wide and anteroposteriorly long bones in dorsal view, but narrow in anterior view. Based on the preserved parts, a medial depression most likely excavated the anterodorsal surface of the joined nasals. Each nasal barely takes part to the corresponding premaxillary crest.
These specimens share many similarities with the holotype and only known specimen of Khoikhoicetus agulhasis SAM PQ 2678 ; the most salient are the rostral maxillary crest being dome-like, the medialmost angle of the right premaxillary sac fossa projecting medially towards the rounded medial edge of the left fossa, the medial margins of the right and left ascending processes converging slightly posterodorsally, the dorsal margin of each premaxillary crest sloping markedly ventrolaterally, and the large nasal on the vertex in dorsal view.
However, several differences are observed. In addition to their larger size ( Table 1), the two Kerguelen specimens differ from SAM PQ 2678 in the following features:
1) The rostrum is distinctly triangular in dorsal view, with straight lateral edges; it is wider at its base and does not widen at mid-length as in SAM PQ 2678.
2) In anterior view, the premaxillary crests are distinctly wider than the premaxillary sac fossae, whereas they are significantly narrower in SAM PQ 2678. The condition on the right premaxillary crest of SAM PQ 2678 may be accentuated by wear and/or break of the lateralmost part; indeed, in anterior view and, as compared to the left side, the right crest seems truncated. Nevertheless, the difference is still valid for the left premaxillary crest, which is apparently unworn in SAM PQ 2678.
3) In lateral view, the premaxillary crests are located posterior to the premaxillary sac fossae, whereas in SAM PQ 2678 they are anteriorly projected and slightly overhang the premaxillary sac fossae.
4) In dorsal view, the bony nares are proportionally narrower and roughly oval-shaped, whereas they are wider and roughly circular in SAM PQ 2678. Because the three specimens of Khoikhoicetus are of different size, three measurements have been selected to establish comparable ratios for the relative width of the bony nares: the width between the premaxillary foramina (Wpf), the width between the main dorsal infraorbital foramina (Wdiof), and the length of the skull from the apex of rostrum to the posteriormost edge of the nasals (Pona). To establish the first two measurements, widths at lateral and medial edges of foramina have been measured, and the mean calculated has been used for Wpf and Wdiof. To establish the third measurement, we had to estimate the length of the rostrum of SAM PQ 2678, which is incomplete. The apex of the rostrum was reconstructed extending the lateral edges anteriorly, with a rounded end as in the two Kerguelen specimens ( Fig. 4). This reconstruction suggests that about 30 mm are missing anteriorly. Therefore, the Pona of SAM PQ 2678 is the measurement taken from the specimen as preserved, to which we added 30 mm. Table 2 clearly establishes that the two specimens from Kerguelen Islands have relatively narrower bony nares compared to the holotype of Khoikhoicetus agulhasis and resemble each other in this respect more than they do the South African specimen.
5) The premaxillary sac fossae are proportionally significantly narrower than those of the holotype of Khoikhoicetus agulhasis . As for the relative width of the bony nares, the measurements of the width of the premaxillary sac fossae (Wpsf) of the three specimens have been compared on the basis of the three measurements mentioned above, Wpf, Wdiof, and Pona. Table 3 clearly establishes that the two specimens from Kerguelen Islands have relatively narrower premaxillary sac fossae than the holotype of Khoikhoicetus agulhasis and resemble each other in this respect more than they do the South African specimen.
and 6) The main dorsal infraorbital foramen of MNHN.F.COI. 1 and 13 is notably distant from the maxilla-premaxilla suture (from 10 to 20 mm), whereas it almost contacts this suture in SAM PQ 2678.
It may be tempting to interpret these differences as related to significant individual variation (ontogenetic and/or related to sexual dimorphism, as commonly observed in extant ziphiids). In order to evaluate if the disparity between the Kerguelen and South African specimens could be related to individual variation, we have referred to the variation observed in 10 specimens of the extant Mesoplodon layardii (Gray, 1865) and eight specimens of the extant Mesoplodon densirostris (Blainville, 1817) ( Ross 1984: tables 8 and 25). We excluded from our sample the specimen of M. layardii mentioned by Ross (1984: table 8) as “ Falkland Islands, Turner, 1880 ”, which, given its size, is likely to be a juvenile, and the specimen of M.densirostris PEM1518 /84 ( Ross 1984: table 25), which is a foetus. We used two measurements provided by Ross (31 and 37), which correspond respectively to our width of the premaxillary sac fossae (Wpsf) and width of the bony nares (Wn). It is noteworthy that measurements of Ross (1984: tables 8 and 25) are expressed as percentages of the condylobasal length, a measurement not available for our specimens and replaced here by the length of the skull from the apex of rostrum to the posteriormost edge of nasals (Pona). However, this difference is likely to be of little incidence on our evaluation of the individual variation and on the comparison with our fossil sample. For the two extant species of Mesoplodon Gervais, 1850 , we calculated the difference between the maximum and minimum value for each measurement and calculated the percentage of this difference to the minimum value. As observed in Table 4, the variation observed in our sample of Khoikhoicetus Bianucci, Lambert & Post, 2007 is much greater that in the two Mesoplodon species , especially in the case of the width of the premaxillary sac fossae, the disparity being less extreme in the case of the width of the bony nares. Because the variation observed in the three Khoikhoicetus specimens exceeds significantly that observed in two species of extant Mesoplodon , it is regarded here as related to interspecific rather that intraspecific variation.Clearly resembling each other more than they do the South African holotype of Khoikhoicetus agulhasis , the specimens from Kerguelen Islands are therefore referred to a new species of Khoikhoicetus , K. kergueleni n. sp.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |