
Lonicera
|
publication ID |
https://doi.org/ 10.1016/j.phytochem.2018.07.012 |
|
DOI |
https://doi.org/10.5281/zenodo.10514461 |
|
persistent identifier |
https://treatment.plazi.org/id/2B1887CC-FFAA-1F03-FF99-814CFBFEFDF0 |
|
treatment provided by |
Felipe |
|
scientific name |
Lonicera |
| status |
|
2.1. Genomic diversity and phylogeny among Lonicera View in CoL species
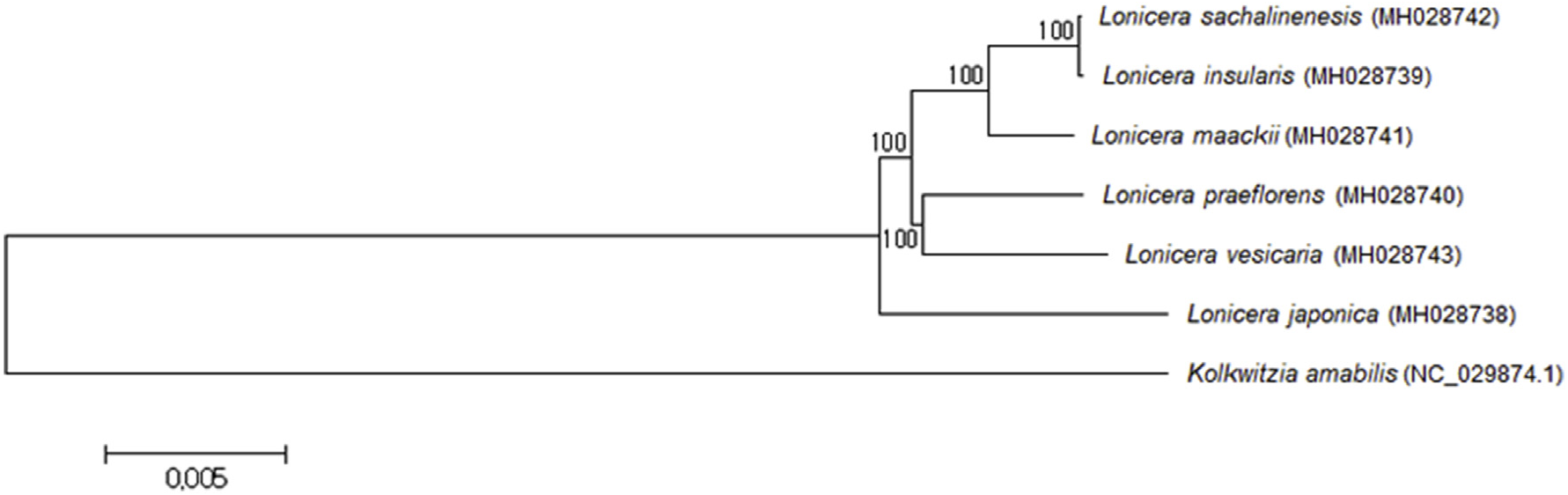
The complete chloroplast genome sequences of six Lonicera species were obtained by assembly of approximately 1 Gbp of whole genome sequences for each species. The completely assembled sequences were ranged from 154,892 to 155,318 bp (GenBank nos. MH028738, Lj; MH028739, Li; MH028740, Lp; MH028741, Lm; MH028742, Ls; MH028743, Lv) ( Table 1 View Table 1 ). Diverse polymorphism among these plants was revealed by the comparative analysis. We identified 17–2261 SNPs and 5–278 InDels between species. The lowest numbers of SNPs and InDels (17 and 5) were identified between L. insularis and L. sachalinensis a; meanwhile, the highest numbers of SNPs (2,261) were identified between L. vesicaria and L. japonica and the highest numbers of InDels (278) were identified between L. insularis and L. japonica ( Table 2 View Table 2 ). The phylogenetic tree revealed that L. japonica is most diverse and grouped into an independent group ( Fig. 1 View Fig ). L. insularis and L. sachalinensis were the closest, and they belonged to the same subgroup as L. maackii . L. praeflorens and L. vesicaria were classified into another subgroup.
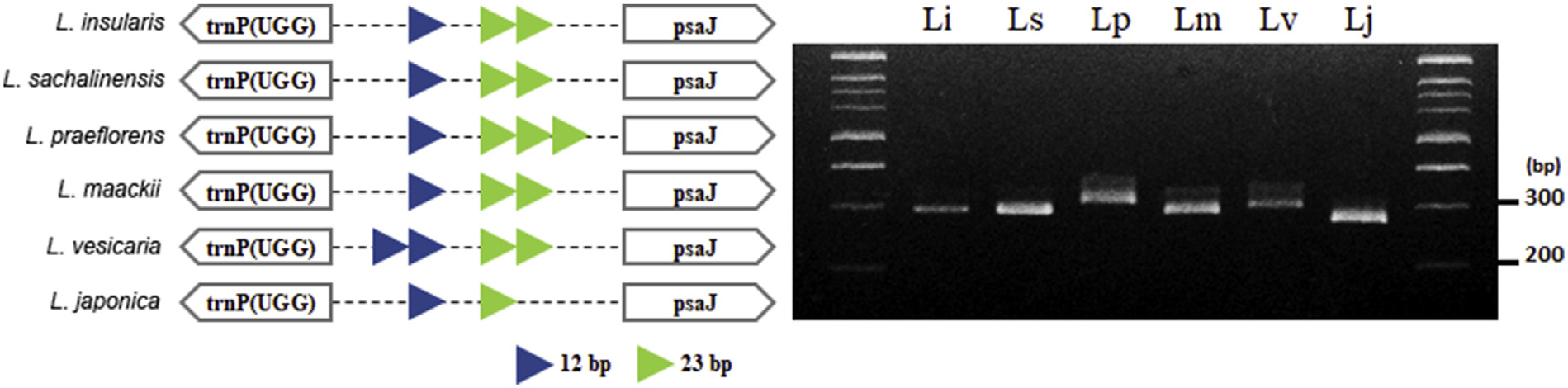
2.2. Development of DNA marker to authenticate the Lonicera species
We developed a DNA marker, named as Lo_i_04, to validate the chloroplast genome sequence assembly and to serve a further application in the authentication of each species. PCR primers were developed for identifying the Lonicera species based on the copy number variation (CNV) of the tandem repeat units in the chloroplast genomes. The CNVbased InDel variation was estimated, and the PCR result coincided with the sequence-based estimation ( Fig. 2 View Fig ). Using this DNA marker, the genomic diversity of L. insularis , L. sachalinensis , and L. maackii to other related species could be authenticated. These three species cannot be distinguished by this DNA marker alone; however, as mentioned above, many more SNPs and InDels between Lonicera species were characterized ( Table 2 View Table 2 ). Thus, we expect that we could develop more DNA markers for establishing a practical authentication system for Lonicera species. Because of the increasing demands for L. japonica in the medicinal herb market, quality control has been an important issue for this species. Thus, an authentic DNA marker for identifying these species can be utilized to prevent adulteration or misuses of other Lonicera species as L. japonica .
2.3. Tentative identification of metabolites
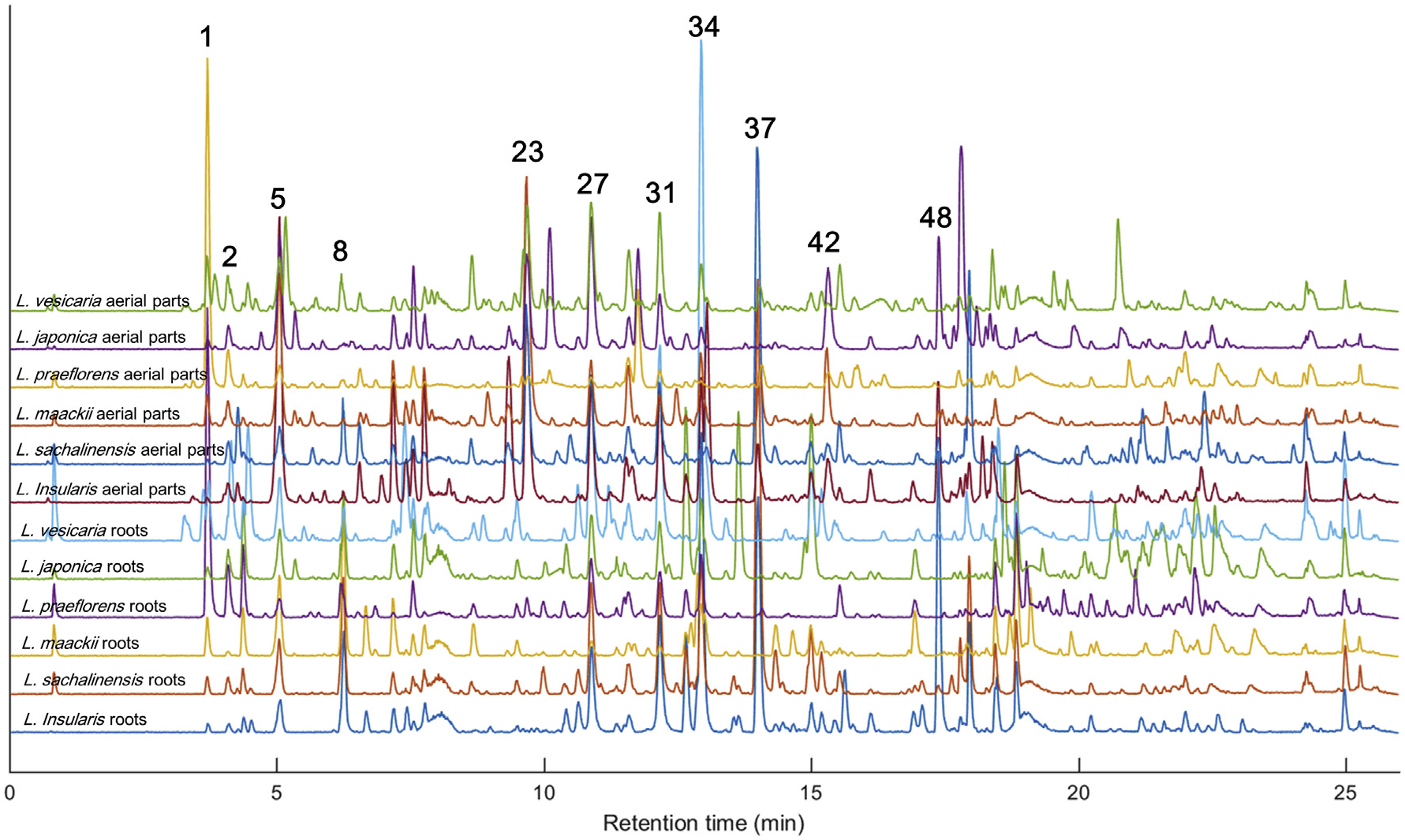
The UHPLC–Q/TOF–MS analysis of the aerial parts and root extracts of six Lonicera species exhibited base peak ion (BPI) chromatograms as shown in Fig. 3 View Fig . The MS E method ( Plumb et al., 2006) allowed us to acquire high-energy collision-induced dissociation (CID) MS data for tentative identification of the major chromatographic peaks. Flavonoids, phenolic acids, iridoids, and their glycosides have been closely investigated for their MS/MS fragmentation ( Es-Safi et al., 2007; Fabre et al., 2001; Jaiswal et al., 2014; March et al., 2006); hence, many peaks could be tentatively identified based on their high-energy CID MS spectra as shown in Table 3 View Table 3 . In our previous study, 13 iridoids and secoiridoids were isolated and identified from the roots of the Korean endemic species L. insularis ( Kang et al., 2018) ; thus, these isolated compounds were also injected to confirm the identification of peaks 1, 5, 8, 9, 14, 15, 20, 33, 37, and 48. Details on the tentative identification are described in the Supplementary Data ( Figs. S1–S View Fig 23).
2.4. Chemotaxonomy among Lonicera species samples
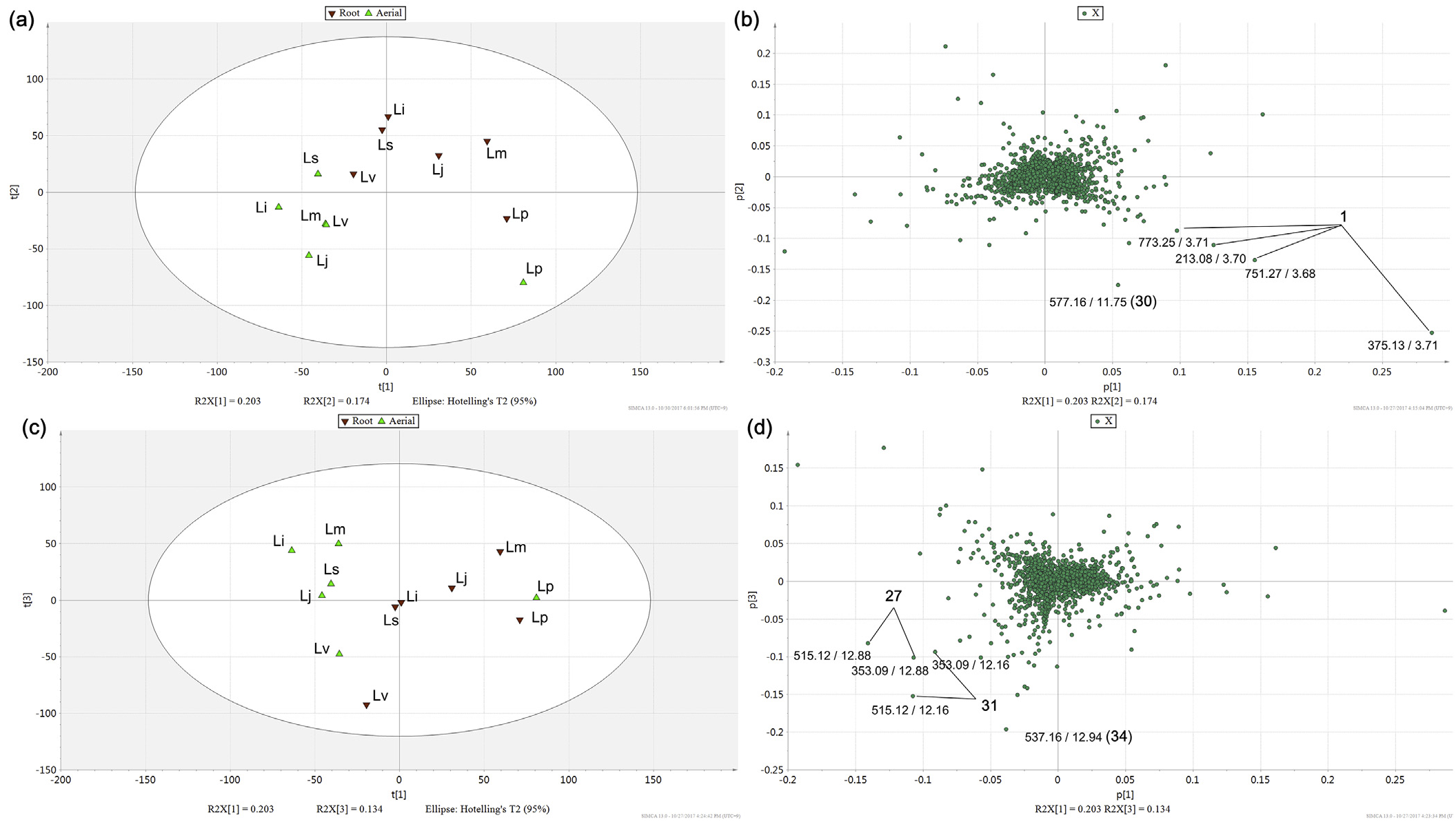
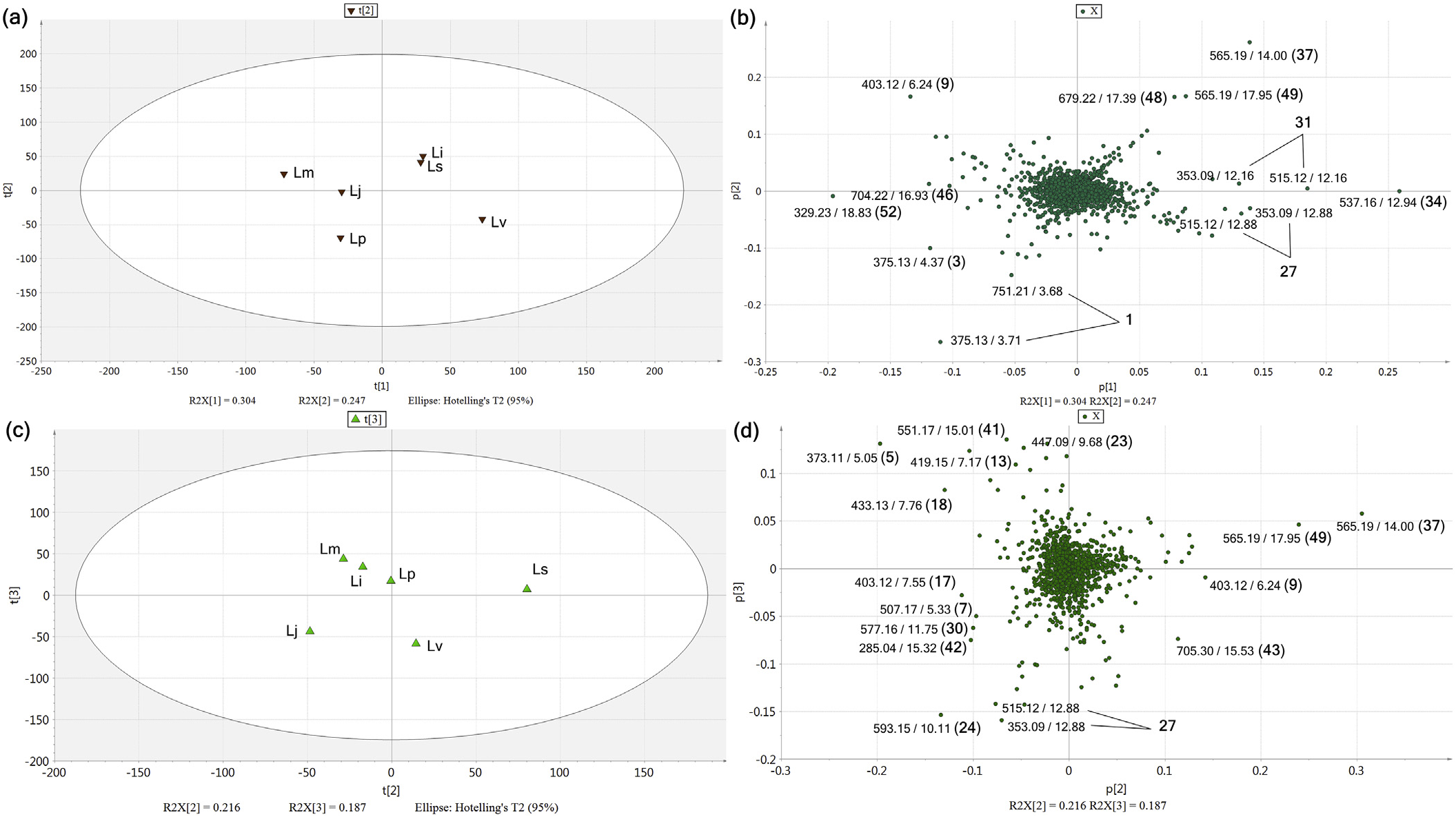
1471 MS ion markers were extracted from the LC–MS dataset, and a principal component analysis (PCA) was performed with them to analyze the chemodiversity among samples. A PCA model with three principle components (PC) was established in which PC1, PC2, and PC3 accounted for 20.3%, 17.4%, and 13.4% of the total variance, respectively (Supplementary Data, Fig. S24a). The PC1-PC2 score plot ( Fig. 4a View Fig ) showed that every sample was distributed in the Hotelling's T 2 95% confidence ellipse, which means the analysis did not contain any outlier. The aerial parts and roots of L. praeflorens were separately grouped from the other species, with positive PC1 and negative PC2 values. The PC1-PC2 loading plot ( Fig. 4b View Fig ) revealed that this separation was caused by the relatively high content of loganic acid (1) in L. praeflorens . This could also be ascertained in the BPI chromatograms ( Fig. 3 View Fig ) and the MarkerLynx ion marker table in which the ion intensities of loganic acid in the L. praeflorens samples were more than five times higher than those of the other species. Iridoid glycosides are well-known as plant derived defense metabolites against herbivores or pathogen ( Dobler et al., 2011). Whitehead and Bowers revealed that in Lonicera plants, iridoid glycosides show significantly higher concentrations in fruits than in leaves, which was suggested to defend fruits against antagonistic seed predators and fruit pathogens ( Whitehead and Bowers, 2013). L. praeflorens bear fruits between May and June while most of other Lonicera species bear fruits between July and August ( “ praeflorens ” means flowering early). Plant samples used in this study were harvested in early July, so it could be suggested that L. praeflorens biosynthesized significantly higher amount of iridoid glycosides, especially loganic acid, to defend fruits. In the PC1-PC3 score plot ( Fig. 4c View Fig ), the roots and aerial parts of L. vesicaria were separated from the other species by their PC3 values. L. vesicaria showed relatively high contents of dicaffeoylquinic acids (27 and 31) and grandifloroside (34), which was suggested by the PC1-PC3 loading plot ( Fig. 4d View Fig ). The BPI chromatogram of the L. vesicaria roots supported this, showing especially high intensity of peak 34. For the other samples, the roots and aerial parts tended to be separated in the scatter plots ( Fig. 4a and c View Fig ).
For further investigation of the chemical diversity among the Lonicera species, additional PCAs were performed within the aerial parts and roots separately. In the PCA model within six Lonicera roots ( Fig. 5a and b View Fig ), in which PC1, PC2, and PC3 accounted for 30.4%, 24.7%, and 19.1% of the total variance respectively (Supplementary Data, Fig. S24b), the samples showed a different distribution of iridoid and secoiridoid derivatives. L. praeflorens and L. vesicaria showed similar patterns to the first PCA result, showing significantly high contents of loganic acid (1) (Lp), and dicaffeoylquinic acids (27 and 31) and grandifloroside (34) (Lv). 7-Desmethylsecologanol (3) was also abundant in L. praeflorens . L. insularis and L. sachalinensis exhibited very similar metabolite profiles, in which periclymenoside (37), kinginoside (48), and methylgrandifloroside methyl ester (49) showed high ion intensities. These three compounds and grandifloroside (34) share a structural trait; they commonly contain a feruloyl moiety in their structures. Periclymenoside and kinginoside have been reported from only a small number of Lonicera species, L. periclymenum ( Calis et al., 1984) , L. morrowii ( Aimi et al., 1993) , and L. insularis ( Kang et al., 2018) . From these, it could be proposed that the biosynthetic ability for feruloyl iridoid derivatives recently appeared during the speciation of these species. L. maackii showed a relative abundance of sweroside (9). PC3 did not show a significant difference between species (Supplementary Data, Figs. S25a and S25b).
Another PCA model was established within the LC–MS dataset from the aerial parts of five Lonicera species ( Fig. 5c and d View Fig ). In this mode, PC1, PC2, and PC3 accounted for 34.2%, 21.6%, and 18.7% of the total variance (Supplementary Data, Fig. S24c). However, PC1 majorly shows the variance between L. praeflorens and other species which were already investigated in Fig. 4 View Fig (Supplementary Data, Figs. S25c and S25d); thus, further variance between other five species were visualized using PC2-PC3 plots. The aerial parts of L. sachalinensis showed a significant abundance of periclymenoside (37) and methylgrandifloroside methyl ester (49) as similar to the root sample, whereas the aerial parts of L. insularis exhibited a different chemical profile. L. insularis and L. mackii showed similar chemical profiles which were relatively abundant in secologanic acid (5) and unidentified iridoid derivatives (13 and 18). L. japonica showed relatively high contents of flavonoids and phenolic acids, such as luteolin 7- O -(6- O -rhamnosylhexoside) (24), 3,4- di- O -caffeoylquinic acid (27), and apigenin 7- O -(6- O -glucosylrhamnoside) (30). Sweroside (9) was identified as a chemical marker for L. maackii in the root sample based the PCA model, but in the aerial parts it was accumulated abundantly in L. sachalinensis and L. vesicaria .
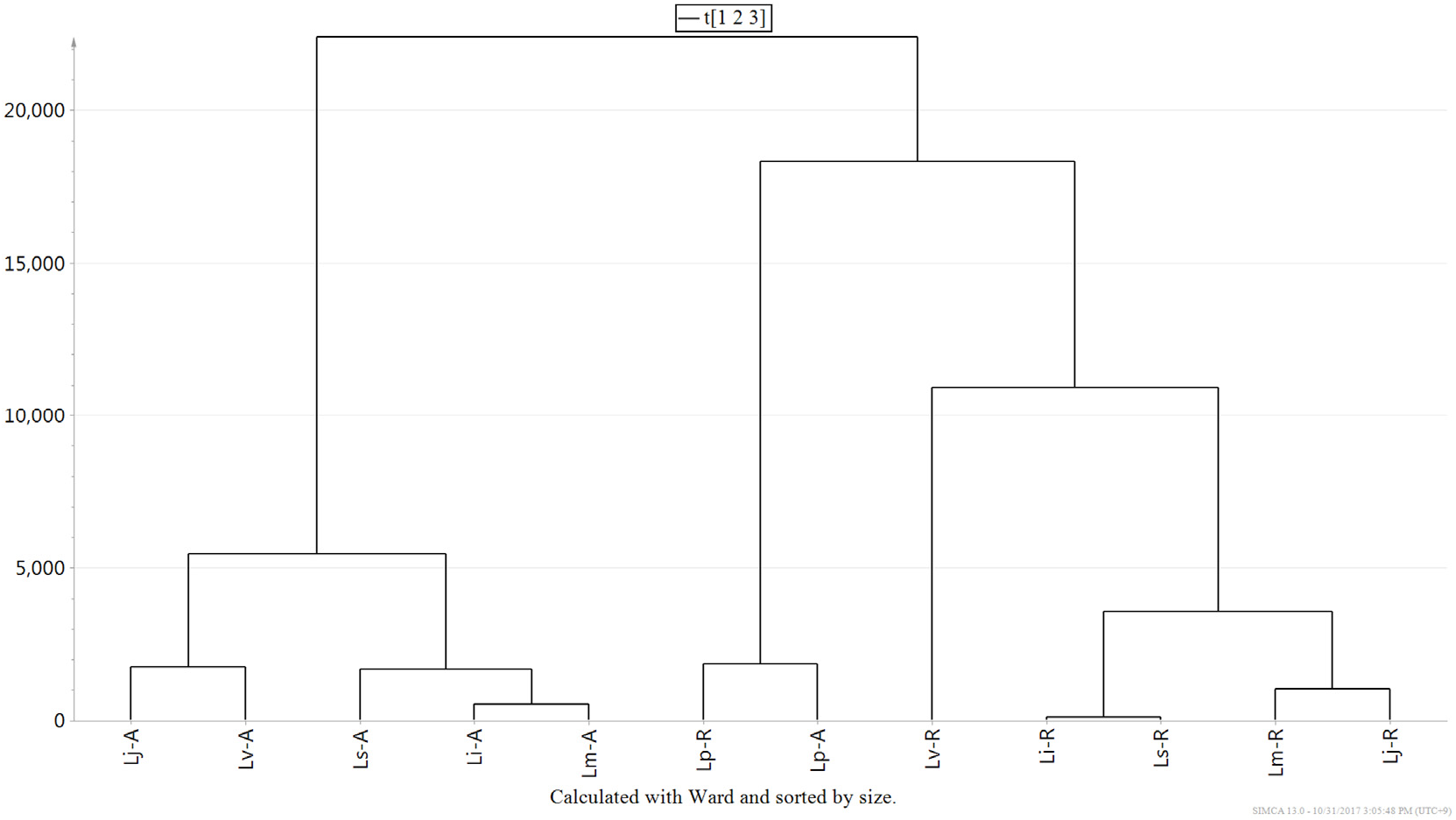
The chemotaxonomic relationship among Lonicera species was established by a hierarchical clustering analysis (HCA) among 12 samples ( Fig. 6 View Fig ). The distances between samples were calculated using Ward's method and Euclidian metrics, and the tree was sorted by size. The samples were divided into three groups: the roots and aerial parts of L. praeflorens , the aerial parts of the other species, and the roots of the other species. As shown in the PC1-PC2 score plot, the chemical contents of L. praeflorens were quite different from that of the other five species. The chemophylogeny among the roots did not correspond to one between the aerial parts or to the phylogenetic tree based on the chloroplast DNA sequences. This type of discordance between genotypes and chemotypes is common because plant specialized metabolite phenotypes are determined by the complex contribution and interaction of genotype and environmental effects ( Chen et al., 2015; Hamilton et al., 2001). Nevertheless, some partial consistency can still be found in the phylogenetic and phylochemical trees; for example, the roots of L. insularis and L. sachalinensis exhibited similar chemical profiles. L. insularis , L. sachalinensis , and L. maackii formed a cluster for their aerial part metabolites, as they did in the chloroplast genome-based phylogenetic tree.
2.5. Metabolite localization in Lonicera species
To further investigate the chemical diversity among the Lonicera samples, additional multivariate analysis models were established. At first, based on the PCA and HCA results, an orthogonal projections to latent structures discriminant analysis (OPLS-DA) model was built with ten samples except L. praeflorens to examine the metabolic difference between the aerial parts and roots of Lonicera plants. The OPLS-DA model exhibited an acceptable predictability, showing R 2 and Q 2 values of 0.989 and 0.755, respectively. The OPLS-DA score plot and the S -plot were used for visualization of the ion markers that influenced the model ( Fig. 7 View Fig ). Several iridoid derivatives showed a significantly higher content in the roots, such as 7-desmethylsecologanol (3), sweroside (9), (E)-aldosecologanin (33), and grandifloroside (34), whereas the flavonoids (23 and 29) tended to subsist in the aerial parts. Interestingly, secologanin (5) is a secoiridoid derivative, but it showed a high content in the aerial parts of Lonicera species rather than in their roots. We could set two hypothesis for this characteristic localization of secologanin. As mentioned above, iridoid glycosides are well-known as plant derived defense metabolites. Peñuelas and coworkers reported that eggs of the herbivore Euphydryas aurinia significantly increased the concentration of iridoid glycosides in leaves of Lonicera implexa , and secologanic acid was the most abundant derivative ( Peñuelas et al., 2006). Based on these facts, we could hypothesize that secologanin was localized at leaves, because it might have specific selectivity against herbivores than other iridoid glycosides. Another hypothesis was related to the biosynthetic pathway of secoiridoids; secologanin is known to be formed by the oxidative cleavage of loganin, which is catalyzed by secologanin synthase (SLS) ( Irmler et al., 2000). Despite early studies on the secoiridoid biosynthesis pathway, especially the identification of SLS performed with suspension cultured cells of L. japonica (Yamamoto et al., 1999, 2000), little is known about the gene expression, protein localization, and metabolite accumulation involved with secoiridoid biosynthesis in Lonicera plants. Y. Liu and coworkers showed that the expression of the SLS2 gene is higher in the stems, leaves, and flowers than in the roots in the case of the Tibetan medicinal plant Swertia mussotii ( Liu et al., 2017) . This was contrary to the high expression of SLS1 and SLS 2 in Catharanthus roseus roots ( de Bernonville et al., 2015), which suggests that the localization of SLS can differ across plant taxa. A. Rai and coworkers reported a significantly high expression of SLS in the young leaf tissues of L. japonica , but they did not use the root tissue in the study ( Rai et al., 2017). Thus, it could be hypothesized that in the case of Lonicera plants, SLS is localized in the leaves while other iridoid biosynthetic cascades are localized in the roots. However, further investigation is required to confirm these hypotheses on localization of secologanin in Lonicera .

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |