
Necremnus aenigmaticus, Gebiola & Bernardo & Ribes & Gibson, 2015
|
publication ID |
https://doi.org/ 10.1111/zoj.12210 |
|
persistent identifier |
https://treatment.plazi.org/id/293AD62E-FFBC-341A-1097-F977FEF3FE27 |
|
treatment provided by |
Felipe |
|
scientific name |
Necremnus aenigmaticus |
| status |
SP. NOV. |
NECREMNUS AENIGMATICUS GIBSON SP. NOV.
( FIGS 92–102 View Figures 92–97 View Figures 98–102 )
Etymology
The species name is derived from the Latin aenigma, ‘something obscure, a mystery’, in reference to its molecular distinctiveness but otherwise yet question-
able morphological differentiation from other similar species as N. duplicatus and N. hippia .
Type material
Holotype ♀ (CNC). GERMANY: Schleswig-Holstein, Bellin, 4.vii.2010, ex. Ceutorhynchus typhae on Capsella bursa-pastoris in canola field, T. Haye (pointmounted, critical-point dried, entire). Allotype ♂ (CNC). Same data and condition as holotype. Paratypes (61♀, 15♂): DENMARK: Jylland, Bov, 24/9 1955, K.-J. Hedqvist (2♀ BMNH). Jylland, Klitmølle, 22/9 1955, K.-J. Hedqvist (1♀ BMNH). ENGLAND: B. Cooke Coll. 84-52, Hippis W (1♀ BMNH). CB, Manea, A.L. Winfield, B.M. 1959- 582, ex. pod of Brassica juncea var. Trowse, opened 24.vii.1959, pupa within em. 6.viii.1959 (1♀ BMNH). Harpenden dist., 11.IX.1943 (4♀, BMNH). FRANCE: Moulinet, sweeping herbaceous layer, NT165 (1♀). GERMANY: 21♀ (including NT90, NT92, NT93) and 14♂ (including NT67, NT91) same data as holotype. Bonn, IX.36, Weiss-Wichert, f. Turnip shoots (1♀, 1♂ BMNH). Nahkamp, Ostholstein, collected 14.VI.2000, ref. no.: #2 (1♀ NMBE), ref. no.: (1♀ NMBE), Capsella bursa-pastoris ex. Ceutorhynchus floralis . Scharnhagen nearby Kiel, S-H [or] Schleswig-Holstein, Coll: 16.06.01, Em: 28 (1♀), 29(1♀) 0.06.01, 02 (1♀), 03 (1♀), 04 (4♀), 05 (1♀), 06(1♀), 09(2♀) 0.07.01, Beate Klander, Scharnhagen/SCH, GPS latitude N54°26.972, GPS longitude E10°06.306, Ex: Ce. uto. floralis, In : C. bursa-pastoris , Leg. Klander/CABI-CH (all NMBE). SWEDEN: Sk., Gislöv, 8/9 1955, K.-J. Hedqvist (7♀ BMNH). Sk., Åhus, 3/8 1956, K.-J. Hedqvist (1♀ BMNH). Sk., Karpalund, 6/8 1967, K.-J. Hedqvist (1♀ BMNH). Sm., Visingö, 19/10 1955, K.-J. Hedqvist (4♀ BMNH). Upl., Vallentuna, 1/6 1956, K.-J. Hedqvist (1♀ BMNH). UNKNOWN LOCALITY: three handwritten labels with Ptilius / Necremnus / Ptilius (1♀ BMNH).
Additional material (2♀): ENGLAND: Little Cheyne, Kent, 1975, ex. Ceutorhynchus sp. in B. napus pods, leg. J. Dale (1♀, coll. R. R. Askew). SPAIN: Cabrils, Barcelona, 31.iii.2005, on Sinapis alba flowers, IRTA (1♀, coll. A. Ribes); Sarroca, Lleida, ex. Diplotaxis erucoides , col. 11.iv.2014, em. 21–30.iv.2014 (1 ♀ 2 ♂, coll. A. Ribes).
Description
Female (holotype, Fig. 93 View Figures 92–97 )
Body 1.8 mm in length. Head primarily green, but scrobal depression dark and under some angles of light parascrobal region, gena and lower face with slight coppery lustre, and vertex slightly more bluish-green ( Fig. 100 View Figures 98–102 ). Antenna ( Fig. 101 View Figures 98–102 ) with scape similarly dark as flagellum; length of flagellum + pedicel about 1.1 × width of head; flagellum with length of F1 + anelli almost 2.1 × as long as wide and about 1.3 × length of pedicel; F2 about 1.9 ×, F3 about 1.8 ×, and clava about 3 × as long as wide. Mesosoma green with slight coppery lustre, most distinctly on scutellum ( Figs 92, 96 View Figures 92–97 ); tegula pale along inner margin and basally but brown apically ( Fig. 94 View Figures 92–97 ). Mesonotum with mesoscutum reticulate; scutellum longitudinally reticulate-imbricate on either side of midline ( Fig. 96 View Figures 92–97 ). Fore wing ( Fig. 97 View Figures 92–97 ) hyaline; speculum broadly bare dorsally; basal cell and speculum posteriorly delineated by complete rows of setae, and mediocubital fold with one seta basal to basal fold; subcubital setal line composed of one row of setae along most of length; basal cell ventrally with two or three spots or minute setae mesally near submarginal vein; postmarginal vein 1.6 × length of stigmal vein. Legs ( Figs 92, 93 View Figures 92–97 ) dark except protibia dorsolongitudinally, knees narrowly, mesotibia narrowly apically, and basal tarsomeres of meso- and metatarsi pale, with second tarsomeres more brownishwhite and apical two tarsomeres brown. Metanotum with dorsellum mesh-like coriaceous ( Fig. 96 View Figures 92–97 ). Propodeum ( Fig. 96 View Figures 92–97 ) with complete median carina; more finely mesh-like coriaceous to alutaceous than dorsellum; spiracle obliquely oval, the rim anteriorly distinctly overlapped by metanotum. Gaster ( Figs 92, 93 View Figures 92–97 ) similarly green as mesosoma except posterior margins of basal four tergites broadly brown; about 1.5 × as long as wide (inflated) and as long as combined length of head + mesosoma; syntergum short.
Male (allotype, Fig. 98 View Figures 98–102 )
Body almost 1.5 mm in length. Similar to female except fore wings with three or five setae on mediocubital fold basal to basal fold and postmarginal vein about 1.7 × as long as stigmal vein. Antenna with scape about 3 × as long as wide; length of funiculars and clava about 1.0, 1.4, 1.7, 2.25, and 2.75 × length of pedicel, with F4 about 3.9 × and clava about 3.3 × as long as wide; rami with long, hair-like setae, elongate-slender with R1 about 2.7 × as long as scape, and all rami lacking mps ( Fig. 99 View Figures 98–102 ).
Variation
Female body length varies from about 1.7–2.3 mm; variably bright green and sometimes without or with only very limited, obscure coppery lustre; tegula sometimes entirely yellow ( Fig. 95 View Figures 92–97 ); mesotibia often entirely dark except basally; and gaster sometimes with up to basal four tergites entirely or almost entirely brown dorsally. Flagellum with length of first funicular + anelli about 1.3–1.5 × length of pedicel; F2 and F3 usually comparatively shorter, and clava about 2.4–3.2 × as long as wide. Fore wing with one or two setae beyond basal fold; basal cell often lacking minute spots mesally near submarginal vein. Metanotum and propodeum often more strongly sculptured, the dorsellum sometimes slightly reticulate-imbricate and propodeum sometimes distinctly coriaceous-alutaceous. Propodeum sometimes with anterior margin of spiracle visible under posterior margin of metanotum, but then rim sinuate or depressed. Gaster sometimes up to about 2.2 × as long as wide in air-dried females.
Male body length varies from about 1.25–1.5 mm in length; otherwise similar to allotype except sometimes with only one seta on mediocubital fold basal to basal fold. A single paratype from which the antennae were slide mounted had a row of eight or nine sensory pores within the apical half of the scape ( Fig. 102 View Figures 98–102 ).
Hosts
Ceutorhynchus typhae (= C. floralis ) (Herbst) on Cap. bursa-pastoris , Ceutorhynchus sp. on rape ( B. napus ) and probably some weevils associated with turnip ( Brassica rapa L. var. rapa ), Indian mustard [ B. juncea (L.) var. Trowse] and white wall-rocket [ Diplotaxis erucoides (L.)].
Discussion
We sequenced five individuals from one site in Germany and one female from France. Our inclusion of other BMNH specimens in the type series is based on the females having an entirely or partly yellow tegula ( Figs 94, 95 View Figures 92–97 ) (differentiating N. aenigmaticus from N. hippia and N. duplicatus ), a broad fore wing speculum ( Fig. 97 View Figures 92–97 ) (differentiating N. aenigmaticus from N. tidius ), and partly overlapped propodeal spiracles ( Fig. 96 View Figures 92–97 ) (differentiating N. aenigmaticus from N. leucarthros and N. tidius ). The reared males and the single male associated with the female from turnip also have these features as well as the complete absence of mps from all three rami. Additional sequencing of specimens from throughout Europe is advisable to more confidently establish whether a partly to entirely pale tegula is a valid differentiating feature for both sexes of N. aenigmaticus because this feature appears to be variable for N. tidius . Individuals of N. metalarus also have the tegula bicoloured similar to N. aenigmaticus , but females differ by having a much more strongly reticulate propodeum and the propodeal spiracles distinctly separated from the metanotum ( Fig. 37 View Figures 32–39 ), and males have mps on the basal ramus. The different position of the sensory pores on the scape will probably help differentiate E. aenigmaticus males from those N. tidius males lacking evident mps on the rami. Although the position of the pores within the apical half of the scape was determined for only a single male of N. aenigmaticus ( Fig. 102 View Figures 98–102 ), this is the same position as for N. duplicatus and therefore probably also characteristic for N. aenigmaticus . The more mesal position of the pores on the scape for N. tidius males ( Fig. 171 View Figures 165–171 ) was determined from numerous specimens.
Individuals of N. aenigmaticus were reared from Ce. typhae on Cap. bursa-pastoris in Europe and those of N. duplicatus from the same host in North America. We did not sequence or see any specimens of N. aenigmaticus from North America, but because of the host association it may be discovered there in the future. If tegular colour pattern is indeed a valid differential feature, this should be sufficient to distinguish any N. aenigmaticus ultimately discovered in North America.
NECREMNUS DUPLICATUS GAHAN REVISED STATUS
( FIGS 103–116 View Figures 103–110 View Figures 111–116 )
Necremnus duplicatus Gahan, 1941: 201–203 View in CoL . ♀ holotype (USNM, examined). Tentative synonymy under N. tidius View in CoL by Boucˇek (1959: 152), and formal synonymy under N. tidius View in CoL by Gibson et al. (2005: 384).
Type material
The holotype female of N. duplicatus is point mounted; it lacks its head but otherwise is entire (see http:// www.usnmhymtypes.com/default.asp?Action=Show _Types&Single_Type=True&TypeID=7003).
Description
Female
Body 1.4–2.3 mm in length. Head sometimes mostly brown in smaller individuals, but usually with variably distinct green to bluish-green lustres under most angles of light except within scrobal depression and often partly blue to purple more dorsally or on vertex ( Fig. 114 View Figures 111–116 ). Antenna ( Fig. 111 View Figures 111–116 ) with scape similarly dark as flagellum; length of flagellum + pedicel about 1.0– 1.1 × width of head; flagellum with length of F1 + anelli about 1.6–2.1 × as long as wide and about 1.3–1.6 × length of pedicel, F2 and F3 at most about 1.4 ×, and clava about 2.3–2.9 × as long as wide (Figs). Mesosoma ( Figs 103, 104, 107 View Figures 103–110 ) sometimes mostly dark brown with coppery or violaceous-coppery lustres in smaller individuals, but usually variably dark oliveto bluish-green with variably distinct coppery or violaceous-coppery lustres at least on scutellum paramedially; tegula uniformly brown ( Fig. 108 View Figures 103–110 ). Mesonotum with mesoscutum reticulate; scutellum longitudinally reticulate-imbricate on either side of midline ( Fig. 107 View Figures 103–110 ). Fore wing ( Figs 115, 116 View Figures 111–116 ) hyaline; speculum broadly bare dorsally; basal cell apically and speculum posteriorly delineated by complete rows of setae, and mediocubital row usually with less than but at most only three setae basal to basal fold; basal cell in ventral view without setae ventroapically, at most with a few inconspicuous spots or minute spicule-like setae in line adjacent to submarginal vein; speculum broadly bare dorsally; postmarginal vein 1.5–1.75 × length of stigmal vein. Legs ( Fig. 104 View Figures 103–110 ) dark except protibia dorsolongitudinally, knees narrowly, mesotibia sometimes very narrowly apically, and basal one or two tarsomeres pale. Metanotum ( Figs 109, 110 View Figures 103–110 ) with dorsellum mesh-like coriaceous to reticulate-imbricate or rarely shallowly reticulate. Propodeum ( Figs 109, 110 View Figures 103–110 ) with median carina over most of length; often somewhat more finely sculptured than dorsellum, alutaceous to mesh-like coriaceous; spiracle obliquely oval, the rim at least touching and usually slightly overlapped by metanotum. Gaster ( Fig. 103 View Figures 103–110 ) brown or dorsally under different angles of light with variably extensive and distinct, usually more blue to purple lustres than on mesosoma, except broadly brownish along posterior margins of basal four tergites and more narrowly along posterior margin of fifth tergite; up to about 1.75 × as long as wide in air-dried specimens and about as long as combined length of head + mesosoma ( Fig. 103 View Figures 103–110 ); syntergum short.
Male ( Figs 105, 106 View Figures 103–110 )
Similar to female except for antennal structure and as follows: body often less distinctly metallic green; legs often lighter brown and tibiae sometimes more extensively to almost entirely pale. Antenna ( Figs 112, 113 View Figures 111–116 ) with flagellar rami long and slender, with long, hairlike setae, and without mps ( Fig. 112 View Figures 111–116 ) or only extremely rarely with at most one mps on R3 of one antenna ( Fig. 113 View Figures 111–116 ); scape with row of sensory pores along ventral margin within apical half (cf. Fig. 102 View Figures 98–102 ).
Distribution
North America (see Gibson et al., 2005).
Hosts
Ceutorhynchus obstrictus on B. napus ; Ce. neglectus on Erucastrum gallicum (Wild.) O.E. Schulz and Rorippa palustris (L.) Besser; Ce. omissus on Erysimum cheiranthoides L.; Ce. typhae on Cap. bursa-pastoris .
Discussion
Morphologically, females of N. duplicatus and N. hippia cannot yet be differentiated reliably. However, based on our molecular results, N. duplicatus appears to be restricted to North America and N. hippia to Europe. Males of the two species are easily differentiated by the absence compared with presence of mps on R2 and R 3 in N. duplicatus vs. N. hippia . Males and females of N. duplicatus are also similar to those of N. aenigmaticus , which is also known only from Europe but may eventually be found in North America (see Discussion under latter species). Individuals share a comparatively broad speculum and the propodeal spiracles at least touching if not slightly overlapped by the posterior margin of the metanotum (cf. Figs 96 View Figures 92–97 , 110 View Figures 103–110 ). However, individuals of N. duplicatus have a uniformly dark tegula ( Fig. 108 View Figures 103–110 ), whereas those of N. aenigmaticus have an entirely to partly pale tegula ( Figs 94, 95 View Figures 92–97 ). Males of the two species are additionally similar in lacking mps from all three rami. However, one male from British Columbia (Abbotsford) that we identify as N. duplicatus has a single mps on the outer surface of R3 of the left antenna ( Fig. 113 View Figures 111–116 ), the right antenna lacking mps from all rami. This is similar, if not the identical pattern as the lectotype male of N. tidius ( Fig. 155 View Figures 151–156 ) (see Discussion under latter species). Our identification of the British Columbia male as N. duplicatus rather than N. tidius is based on the propodeal spiracle being slightly overlapped by the metanotum and the individual being reared from canola along with several other females and males typical for the species. Owing to the condition of the antennae of some males it was not always possible to be completely certain that the R2 and R3 ramus completely lacked mps, but presence of at most a single mps is at least very rare for N. duplicatus males. When Gibson et al. (2005) synonymized N. duplicatus under N. tidius , the complete absence of mps from the rami of North American males was noted and illustrated ( Gibson et al., 2005: fig. 3). The lectotype male of N. tidius had been examined, but presence of the single mps on R3 was not observed. The presence of mps on the apical two rami of some European specimens was also discussed, but was assumed to represent intraspecific variation within the very few males then seen from Europe.
In North America, females of N. duplicatus key to couplet 8 and males to couplet 13 using Gahan (1941), who differentiated females of N. leucarthros from those of N. duplicatus by the former having a comparatively longer gaster and a darker marginal vein. The noted gastral difference does appear to be valid (cf. Figs 103 View Figures 103–110 , 140 View Figures 139–144 ). Females of N. leucarthros have a more elongatelanceolate gaster, except rarely at least 1.8 × as long as wide. Females of N. duplicatus have a more ovatelanceolate gaster that is at most about 1.75 × as long as wide. However, this feature is not always sufficient to distinguish females because of artefacts of differential drying (cf. Figs 140 View Figures 139–144 , 145 View Figures 145–150 ). Furthermore, Gahan (1941) did not key N. tidius . Both sexes of N. duplicatus are usually readily differentiated from those of N. leucarthros and N. tidius by placement of the propodeal spiracles. These are slightly separated from the metanotum in N. leucarthros ( Figs 143 View Figures 139–144 , 147 View Figures 145–150 ) and N. tidius ( Figs 172, 173 View Figures 172–175 ). However, it is sometimes difficult to be certain whether or not the spiracles are slightly overlapped by the metanotum vs. touch the metanotum or touch the metanotum vs. being slightly separated from it. Some of the variation in spiracular position is probably explained by the metanotum being slightly moveable relative to the propodeum, but it is also affected by the exact angle at which the mesosoma is viewed (cf. Figs 131, 132 View Figures 125–132 ). Males are additionally distinguished by flagellar features. Males of N. leucarthros are readily identified by uniquely having mps on all three rami and very short ( Fig. 150 View Figures 145–150 ) rather than long, hair-like setae ( Figs 112, 113 View Figures 111–116 ). Except for the rare exception discussed above, males of N. duplicatus lack mps from all three rami whereas those of N. tidius typically have at least one mps on at least R2 and often R3 (see Discussion under N. tidius ). Furthermore, N. duplicatus males have the sensory pores within the apical half of the scape (cf. Fig. 102 View Figures 98–102 ) whereas they are essentially mesal in position for N. tidius males ( Fig. 171 View Figures 165–171 ), the distance between the basal-most pore and the basal margin at most only slightly greater than that between the apical-most pore and apical margin. Females of N. tidius are additionally distinguished from those of the other two species by the fore wings being more densely or at least more extensively setose so that dorsally the speculum is comparatively narrow ( Fig. 168 View Figures 165–171 ), and they usually also have at least three and typically more setae on the mediocubital fold basal to the basal fold ( Fig. 168 View Figures 165–171 ). However, so do most N. leucarthros females. Females of N. duplicatus have fewer than three setae ( Fig. 116 View Figures 111–116 ) except very rarely. Female N. tidius usually also have distinctively more elongate-slender funiculars ( Fig. 167 View Figures 165–171 ) than the funiculars of females of the other two species ( Figs 111 View Figures 111–116 , 149 View Figures 145–150 ). In addition to their more elongate-lanceolate gaster and typically more numerous setae on the mediocubital fold, female N. leucarthros often also have quite distinct although short setae ventroapically within the basal cell when viewed ventrally. They are also typically somewhat brighter green to bluish-green with at most limited, inconspicuous coppery lustre compared with N. duplicatus females (cf. Figs 103 View Figures 103–110 , 145 View Figures 145–150 and 104 View Figures 103–110 , 146 View Figures 145–150 ), although this difference is not always obvious.
NECREMNUS HIPPIA (WALKER) REVISED STATUS
( FIGS 117–138 View Figures 117–124 View Figures 125–132 View Figures 133–138 )
Eulophus Hippia Walker, 1839: 185–186 View in CoL . ♀ lectotype (BMNH, here designated). Synonymy under N. leucarthros View in CoL by Gahan (1941: 201) and Boucˇek (1959: 151), and under N. tidius View in CoL by Graham in Bouček & Askew (1968: 67).
Entedon cyrinus Goureau, 1851: 147 View in CoL , pl. 6, no. VIII, figures 7–9. ♀ syntype (OXUM, examined). Synonymy under N. tidius View in CoL by Graham, 1963: 190. syn. nov.
Type material
Walker (1839) described Eulophus hippia based on at least three females, the description also including two variants. The BMNH has five specimens indicated to form part of the type series. One point-mounted female has the following seven labels: (1) a circular, purplebordered label with ‘LECTOTYPE’; (2) a circular label with ‘38/7.12/53’ on three separate lines; (3) a small, rectangular, handwritten label with ‘Britain’; (4) a rectangular label with ‘ Eulophus Hippia Walker’ handwritten on one side and ‘Stood under this name in old B.M. Coll. C. Waterhouse.’ printed on the other side; (5) a rectangular, handwritten label with ‘Type M. de V. G.’; (6) a rectangular label with ‘ Eulophus hippia Walker LECTOTYPE: ♀ M. de V. Graham det. 1958’ partly printed and handwritten; and (7) a square label with ‘B.M. TYPE HYM. 5.2507’. This female lacks its gaster and most of its middle and hind legs including the tibiae and tarsi except for the right metatibia. The tegula is extensively yellow, only the apex being partly brown ( Fig. 123 View Figures 117–124 ). The head and mesosoma are also much brighter green, with only limited coppery lustre on the mesonotum under some angles of light, than a second female labelled as ‘type’. This latter female is card mounted ( Fig. 117 View Figures 117–124 ) and has five labels: (1) a circular, red-bordered label with ‘Type’; (2) a handwritten label with ‘ Hippia ’; (3) a printed label with ‘Walker coll. 1901–120’; (4) a handwritten label with ‘in BM. under Eulophus hippia Walk. ’; and (5) a handwritten label with ‘this is prob. not type of hippia & is a Necremnus, MG. 1952 .’. This female has entirely dark tegulae ( Fig. 120 View Figures 117–124 ) and is dark with very slight violaceous lustres under some angles of light ( Figs 117, 119 View Figures 117–124 ). The three other BMNH specimens are point mounted and have circular, blue-bordered ‘PARALECTOTYPE’ labels. One of these, a female, has the same double-sided label as for the specimen with Graham’s lectotype labels, whereas another female and one male have printed labels with ‘Standing over Eulophus hippia in BMNH collection viii.2012 ’. The mesonotum of the former female is variably green to coppery under some angles of light and the tegula is partly yellowish along the inner margin and narrowly basally, whereas the latter female and male are very similar to the female labelled as ‘type’. All the specimens have the propodeal spiracles slightly overlapped by the metanotum ( Fig. 121 View Figures 117–124 ) and at least the females have similar fore wing setal patterns with a comparatively broad speculum ( Fig. 122 View Figures 117–124 ) and similar antennae ( Fig. 118 View Figures 117–124 ). The male lacks both antennae. As Graham did not validly designate a lectotype through publication under ICZN rules, all of the specimens remain available for designation. In order to stabilize the concept of the name, we designate as the lectotype of E. hippia the card-mounted female originally labelled as ‘Type’ because the original description specifically states that the tegulae were pitch black (squamulae piceae), it has a handwritten label with ‘ Hippia ’ similar to most other specimens selected as Walker Necremnus lectotypes, it is entire, and it is more likely to remain complete because of its card mount- ing. A rectangular red lectotype label has been added to the specimen. We designate the incomplete, pointmounted female bearing Graham’s lectotype label as a paralectotype of E. hippia , as we do the other two females and one male previously bearing paralectotype labels.
There is no indication in the original publication of the number of females from which Goureau (1851) described Entedon cyrinus . A single female remains in OXUM labelled as ‘Type’. It has the following five labels: (1) a card cut into an L-shape with ‘ex mined leaves of Ranunculaceae- [?], Paras of Phytomyza flava ? Meig’ handwritten; (2) a small, diamond-shaped, blue label with a ‘W’ that is underscored and overscored with a dark line; (3) a circular, red-bordered printed label with ‘-TYPE O.U.M.’; (4) a rectangular, handwritten label with ‘beetle – room drawer 5 left bottom’; and (5) a rectangular, handwritten label with ‘ Entedon cyrinus Goureau, 1851 , OX. UNI. MUS. NAT. HIST. ( OUMNH)’. The female is entire but is in poor condition because it is encased in glue on a curved piece of paper ( Fig. 124 View Figures 117–124 ). However, the anterior rims of the propodeal spiracles appear to be slightly overlapped by the metanotum and the fore wings appear to have a comparatively broad speculum. The tegulae also appear to be entirely dark. For these reasons we newly synonymize Entedon cyrinus Goureau under Necremnus hippia (Walker) , but do not designate the female as the lectotype in case another, better preserved specimen is found in the future.
Description
Female
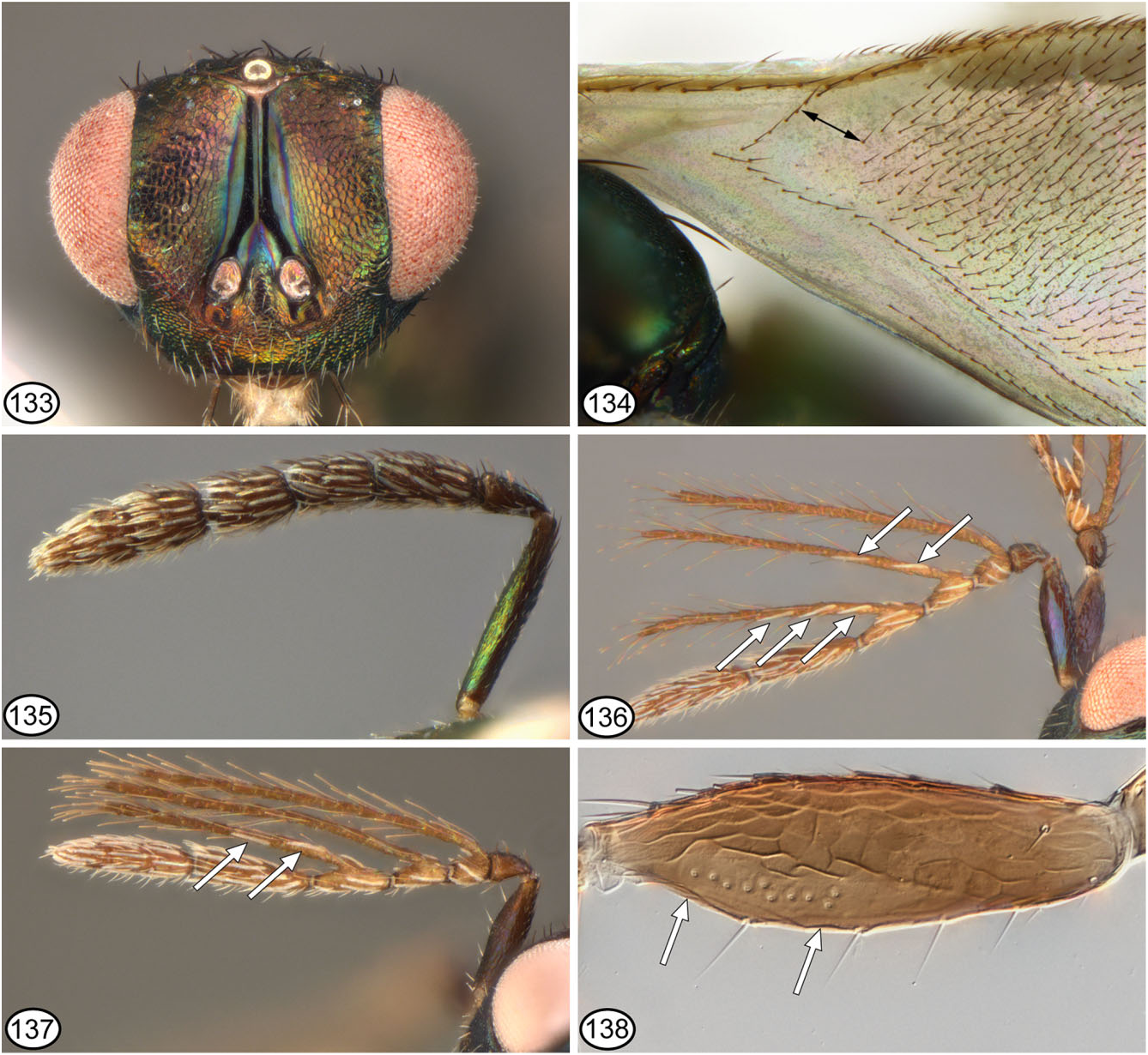
Body 1.1–2.1 mm in length. Head primarily brown in smallest individuals, but usually green with slight coppery lustre to extensively coppery except for scrobal depression ( Fig. 133 View Figures 133–138 ), and vertex often slightly more bluish to purple. Antenna ( Figs 118 View Figures 117–124 , 135 View Figures 133–138 ) with scape similarly dark as flagellum; length of flagellum + pedicel about 1.0–1.2 × width of head; flagellum with length of F1 + anelli 1.4–2.1 × as long as wide and 1.0–1.5 × length of pedicel; F2 and F3 at most about 1.7 ×, and clava 2.7–3.3 × as long as wide. Mesosoma usually green to bluish-green with slight coppery lustre, most distinctly on scutellum ( Figs 125, 130 View Figures 125–132 ), although sometimes darker with more extensive coppery lustre; tegula uniformly dark brown ( Figs 120 View Figures 117–124 , 127 View Figures 125–132 ). Mesonotum with mesoscutum reticulate ( Fig. 119 View Figures 117–124 ); scutellum longitudinally reticulate-imbricate on either side of midline ( Fig. 130 View Figures 125–132 ). Fore wing hyaline; speculum broadly bare dorsally ( Fig. 134 View Figures 133–138 ); basal cell and speculum posteriorly delineated by complete rows of setae, and mediocubital fold with one to three seta basal to basal fold ( Fig. 134 View Figures 133–138 ); subcubital setal line composed of one row of setae along at least about basal half ( Fig. 134 View Figures 133–138 ); basal cell ventrally usually entirely bare, only rarely with one to three spots or minute setae near submarginal vein; postmarginal vein 1.6–1.9 × length of stigmal vein. Legs ( Figs 125, 126 View Figures 125–132 ) dark except protibia dorsolongitudinally, knees narrowly, mesotibia narrowly apically, and basal tarsomeres of meso- and metatarsi pale. Metanotum with dorsellum mesh-like coriaceous to often reticulate-imbricate ( Figs 129, 130 View Figures 125–132 ) or sometimes quite distinctly reticulate posteriorly. Propodeum ( Fig. 121 View Figures 117–124 ) with median carina over most of length; more finely mesh-like coriaceous to alutaceous than dorsellum; spiracle oblique- ly oval, the rim usually variably distinctly overlapped by metanotum ( Figs 129, 130 View Figures 125–132 ), but if anterior margin visible ( Fig. 132 View Figures 125–132 ) then rim sinuate or slightly depressed. Gaster ( Fig. 125 View Figures 125–132 ) variably extensively green to sometimes more blue or purple dorsally, although posterior margins of basal four tergites at least broadly brownish along posterior margins and sometimes up to basal five tergites mostly to entirely brown or copperybrown; about 1.5–1.9 × as long as wide and as long as combined length of head + mesosoma; syntergum short.
Male ( Fig. 128 View Figures 125–132 )
Similar to female except for antennal structure and as follows: basal cell ventrally with at least three and usually more spots or minute setae in line along most of length of submarginal vein; mediocubital fold with up to four setae basal to basal fold. Antenna ( Figs 136, 137 View Figures 133–138 ) with flagellar rami long and slender, with long, hair-like seta, and with variably numerous and conspicuous mps on second and third rami, usually with two or more mps on both rami and rarely with up to five mps on at least third ramus; scape with line of sensory pores along ventral margin obviously more within apical half, the distance between basal-most pore and basal margin much greater than distance between apical-most pore and apical margin, the basal separation more similar to length of sensory row ( Fig. 138 View Figures 133–138 ).
Distribution
Europe ( Italy, Romania, Switzerland).
Hosts
Ceutorhynchus cardariae on Car. draba ; Ce. typhae on Cap. bursa-pastoris .
Discussion
Based on tegular colour pattern, the type series of N. hippia appears to consist of N. hippia in the present sense (those with dark tegulae, Fig. 120 View Figures 117–124 ) as well as N. aenigmaticus (the two females with partly yellowish tegulae, Fig. 123 View Figures 117–124 ). Colour of the tegula is the only feature that we found to reliably differentiate females of N. hippia from those of N. aenigmaticus , although based on observed material N. aenigmaticus females apparently more commonly have minute spots within the basal cell near the submarginal vein. Males of the two species are differentiated both by colour of the tegulae and by presence in N. hippia ( Figs 136, 137 View Figures 133–138 ) or absence in N. aenigmaticus ( Fig. 99 View Figures 98–102 ) of mps on R2 and R3. Owing to their broad fore wing speculum ( Fig. 134 View Figures 133–138 ) and uniformly brown tegula, females of N. hippia are not reliably distinguished from those of N. duplicatus and can also be mistaken for smaller N. leucarthros females. Females of N. leucarthros usually have the propodeal spiracles slightly separated from the metanotum ( Figs 143 View Figures 139–144 , 147 View Figures 145–150 ), although this differential feature is not always obvious, as discussed under N. duplicatus . Female N. leucarthros typically also have a somewhat more elongate-lanceolate gaster and more numerous spots along the submarginal vein in the basal cell that often in ventral view appear as very short but quite distinct setae, at least apically toward the basal fold.












No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Necremnus aenigmaticus
| Gebiola, Marco, Bernardo, Umberto, Ribes, Antoni & Gibson, Gary A. P. 2015 |
Necremnus duplicatus
| Gibson GAP & Baur H & Ulmer B & Dosdall L & Muller F 2005: 384 |
| Gahan AB 1941: 203 |
Entedon cyrinus
| Graham MWR de V. 1963: 190 |
| Goureau CC 1851: 147 |
Eulophus Hippia Walker, 1839: 185–186
| Gahan AB 1941: 201 |
| Walker F 1839: 186 |