
Cephalastor minarum, Garcete-Barrett & Hermes, 2010
|
publication ID |
https://doi.org/ 10.1080/00222930903384774 |
|
persistent identifier |
https://treatment.plazi.org/id/291787AB-FF8D-FFD8-57BA-8D95FE334800 |
|
treatment provided by |
Felipe |
|
scientific name |
Cephalastor minarum |
| status |
sp. nov. |
Cephalastor minarum n. sp.
( Figures 3, 4 View Figures 1–6 , 8 View Figures 7–9 , 11 View Figures 10–13 , 15, 17 View Figures 14–18 , 32, 34 View Figures 31–34 , 38 View Figure 38 )
Diagnosis. This species is very close to C. humeralis in general appearance and structure, differing by having a largest bare area devoid of micropunctures in the middle of the mesoscutum which extends further forward ( Figure 11 View Figures 10–13 ), the ocular sinus is narrower ( Figure 3 View Figures 1–6 ), the pronotal foveae are completely fused together ( Figure 8 View Figures 7–9 ), the submedial propodeal carina is lamellar just up to the propodeal angle ( Figure 32 View Figures 31–34 ), the transverse carina on tergum I is distorted, flattened dorsally and the tumescence behind it is very low, without a central depression ( Figure 34 View Figures 31–34 ), and the pronotal band does not project backwards ( Figure 11 View Figures 10–13 ).
Description. Holotype female: colouration – black with yellow as follows: basal half of mandibles except small black triangle on the very base; small subapical spots narrowly fused, and large basal spots on clypeus; interantennal spot; frontal line; scrobal spots; small line on gena; scape beneath; thin dorsal pronotal band narrowly interrupted at middle; small sub-alar spot; ring-like mark on tegula; posterior scutellar band medially interrupted by a thin line; narrow basal line along lower half of the submedian propodeal carina; tiny condylar maculae adjacent to the bases of mid and hind coxae; most of fore tibia anteriorly and innerly; diffuse apical macula on outer surface of fore femur; tiny suffuse macula at very base of mid and hind tibia; thin apical bands on terga and sterna I–V. Light reddish brown to chestnut are: apex of mandible except the black points of teeth; clypeal apex; apex and posterior surface of scape; pedicel and flagellum beneath (lighter at basal flagellomeres); suffusion on pronotal lobe and along thin hind pronotal margin; tegula; apical suffusion on femora and tibiae (much less visible on mid and hind legs); inner face of mid and hind tibiae; some suffusion along lateral submarginal carina of propodeum; sternum I entirely; lateral suffusion on tergum I; obscure thin lateral suffusion on tergum II. Foveal brushes blackish-brown. Body covered with short, thin brownish appressed pubescence – mostly absent on frons, sides of vertex, gena, upper surface of pronotum and very small mid anterior area of mesoscutum – becoming paler on lower face, clypeus and mesosomal and metasomal sides and venter. Wings light grayish brown, just a bit darker at apex of basal cell and especially along dorsal half of marginal cell. Venation chestnut.
Dimensions: body from frons to apex of tergum II = 6.25 mm. Forewing length = 5.25 mm. Breadth of gena equivalent to 0.6 times the breadth of eye at emargination. Proportion of MOW:POL:OOL:MPOL equivalent to 1:2.5:2.5:1. Upper surface of tergum I 1.95 times wider than long.
Structure: head, clypeus and mesosoma very finely microgranulated as in other species of the genus. Clypeus evenly convex, bearing some rather sparse macropunctures separated by one to two diameters, just a bit flattened on top and shallowly depressed on apical area. Clypeal apex rather transluscent and with margin shallowly concave between teeth, which are separated by just a little more than the interantennal distance. Head with macropunctures separated by 0.5–2 diameters, absent from yellow-marked areas, becoming thinner and sparser on upper gena and coarser over the posterior declivity. Vertex tubercles rather square and with a well-marked middle longitudinal depression; foveal brushes rather small and narrowly separated from each other, but very dense, obscuring the foveae, which are separated by about one diameter apart; posterior declivity slightly convex rather than plainly flat. Gena with a narrow precarinal depression widening down behind the lower ocular lobe. Occipital carina rounded at gena and almost straight medially.
Anterior surface of pronotum with some few almost undiscernible micropunctures sublaterally adjacent to the carina and macropunctures mostly concentrated in two submedial clusters. Pronotal fovea oval. Pronotal carina well developed, almost straight in dorsal view and forming a regular arc in frontal view. Humeral angle sharp and slightly pointing upwards. Posterior corner of pronotum with a slight but well-marked pretegular ridge-like fold, which nevertheless does not form a carina. Pronotal macropunctation much coarser and denser than that of head; becoming denser at sides, with some confluent punctures. Tegulae smoothly reticulo-granulate, semicircular, with a few undiscernible punctures towards the anterior and posterior corners, and without developed outer rim. Macropunctation of mesoscutum coarse and dense anteriorly (similar to that on the upper surface of pronotum), becoming markedly thinner and sparser posteriorly. Micropunctation of mesoscutum rather dense and partially obscuring the microgranulation at lateral and posterior thirds. Scutellum with broad anterior crenation, slight medial longitudinal depressed line and with macropunctation similar to that of mesoscutum, though mostly absent from yellow-marked areas. Metanotum with well-marked toothed carina separating the anterior horizontal surface from the posterior almost vertical surface; both surfaces very coarsely striatopunctate/crenate. Mesepisternum irregularly punctate with sparse, small macropunctures. Metapleura and propodeum with very fine and dense microsculpture. Metapleura with almost no macropunctures apart from those along the anterior crenation. Upper surface of propodeum with very coarse and close elongate alveoles separated by thin careniform intervals and as wide as the horizontal surface of metanotum at middle. Lateral surface of propodeum covered with coarse flat-bottomed alveoles separated by thin, flat to slightly convex intervals. Posterior surface of propodeum with just some scarce thin macropunctures mostly present adjacent to the submedial carina. Submedial carina complete and lamelliform from propodeal angle down to the submarginal carina.
Transvere carina of tergum I sub-trapezoidal and shallowly emarginated on top in frontal view. Tergum I also with a low but regularly developed transverse tumescence crossing the middle of its upper surface. Sternum I with rather sharp basal transverse carina, higher than the longitudinal carina. Sternum II abruptly but roundly elevated sub-basally, preceded by a wide and longitudinally coarse striate groove in ventral view, and followed by a rather flat surface in lateral view. Micropunctation of metasoma very fine and dense, scale-like ( Figure 17 View Figures 14–18 ), sparser on the middle of sternum II, and denser and coarser on terga and sterna III–VI. Macropunctation almost entirely absent from anterior surface of tergum I; coarsely alveolar on the upper surface of the same tergum, especially coarse and dense behind carina and tumescence; very coarsely alveolate with thin careniform intervals on sternum I, except for its mid-apical rim; very fine and sparse on tergum II, becoming denser and coarser towards the apical margin; regularly coarse and sparse on sternum II, with some coarser points close to the hind margin; quite dense but rather fine on terga III and IV apically, and very fine, obscure to almost completely absent on terga and sterna V and VI.
Male unknown.
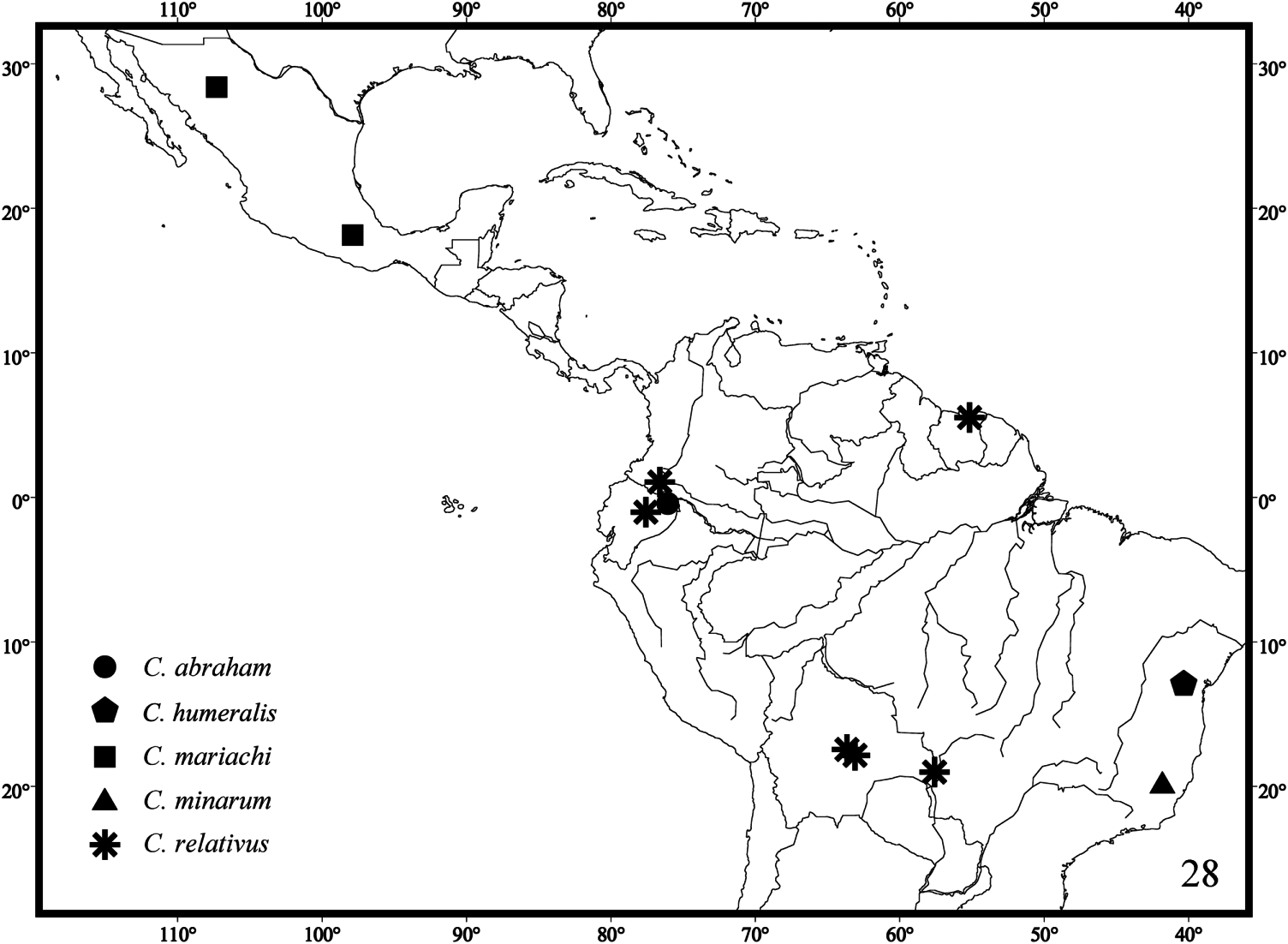
Type material. Holotype female: BRAZIL: MINAS GERAIS: Ipanema, Fazenda Montes Claros , 19°45′S 41°50′W, 30.iv.1998, G. & C. Melo ( DZUP). GoogleMaps
Etymology. The specific epithet refers to the State of Minas Gerais in Brazil, where the specimen was collected.
New collecting records
Two new collecting records were found while examining the Cephalastor material deposited in the DZUP: Cephalastor bossanova : one female: BRAZIL: PARANÁ: Piraquara, Mananciais da Serra, 25°30′S 48°59′W, 28.iv.2001, G.A.R. Melo. Cephalastor estela : one female: BRAZIL: ESPIRITO SANTO: Baixo Guandu, 29.iv-6.v.1970, C. & C.T. Elias ( Figures 37 View Figure 37 and 38 View Figure 38 ).
Phylogeny
List of characters used in the cladisitc analyses of Cephalastor
(1) Clypeal disc: (0) flat; (1) convex ( Figures 1, 3 and 5 View Figures 1–6 )
(2) Clypeus: (0) broader than long ( Figures 1, 3 and 5 View Figures 1–6 ); (1) longer than broad; (2) as broad as long
(3) Apical margin of clypeus: (0) concave ( Figures 1 and 3 View Figures 1–6 ); (1) not concave (figure 5a in Garcete-Barrett [2001])
(4) Cephalic foveae 1: (0) between compound eyes ( Figures 20 and 21 View Figures 19–21 ); (1) behind level of compound eyes ( Figure 19 View Figures 19–21 )
(5) Cephalic foveae between compound eyes (applicable only to terminals that received state 0 for Character 4): (0) one ocellar diameter behind level of lateral ocelli; (1) about half ocellar diameter behind level of lateral ocelli
(6) Cephalic foveae 2: (0) with arc-like carina posteriorly ( Figure 19 View Figures 19–21 ); (1) without arc-like carina posteriorly ( Figures 20 and 21 View Figures 19–21 )
(7) Modified cuticle around cephalic foveae 1: (0) undefined ( Figure 19 View Figures 19–21 ); (1) well defined ( Figures 2, 4, 6 View Figures 1–6 , 20 and 21 View Figures 19–21 )
(8) Modified cuticle around cephalic foveae 2 (applicable only to terminals that received state 1 for Character 7): (0) single transverse elyptical ( Figure 21 View Figures 19–21 ); (1) single subcircular ( Figures 4 View Figures 1–6 and 20 View Figures 19–21 ; figure 3c in Garcete-Barrett [2001]); (2) two small closely set ( Figure 6 View Figures 1–6 )
(9) Vertex: (0) with interocellar tubercles ( Figures 1–6 View Figures 1–6 , 20 and 21 View Figures 19–21 ); (1) without interocellar tubercles ( Figures 19 View Figures 19–21 and 22 View Figures 22–25 )
(10) Interocellar tubercles (applicable only to terminals that received state 0 for Character 9): (0) separated by wide depression ( Figures 1–6 View Figures 1–6 and 21 View Figures 19–21 ; figures 1c, 4b, 6c and 8c in Garcete-Barrett [2001]); (1) fused ( Figure 20 View Figures 19–21 ; figure 5b in Garcete-Barrett [2001]; figures 3c and 4c in Garcete-Barrett 2003)
(11) Head posterior declivity: (0) convex; (1) slightly flattened; (2) concave
(12) Gena: (0) broad ( Figures 14–16 View Figures 14–18 and 22 View Figures 22–25 ); (1) narrow ( Figure 23 View Figures 22–25 )
(13) Anterior pronotal surface 1: (0) with foveae ( Figures 2, 6–9 View Figures 1–6 View Figures 7–9 , 19–21 View Figures 19–21 ); (1) without foveae
(14) Foveae of anterior pronotal surface (applicable only to terminals that received state 0 for Character 13): (0) contiguous to completely fused ( Figures 6–9 View Figures 1–6 View Figures 7–9 and 20 View Figures 19–21 ; figures 1l, 2e, 4e and 5g in Garcete-Barrett [2001]); (1) not contiguous ( Figures 19 and 21 View Figures 19–21 ; figures 1e, 2e, 3h and 4f in Garcete- Barrett [2003])
(15) Anterior pronotal surface 2: (0) punctate ( Figures 2, 6–9 View Figures 1–6 View Figures 7–9 , 21 View Figures 19–21 and 23 View Figures 22–25 ); (1) not punctate ( Figures 19, 20 View Figures 19–21 and 22 View Figures 22–25 )
(16) Anterior pronotal surface 3: (0) concave ( Figure 22 View Figures 22–25 ); (1) convex ( Figure 23 View Figures 22–25 )
(17) Pronotal carina: (0) low obtuse dorsally; (1) sharp dorsally
(18) Background microsculpture of head and dorsal mesosoma: (0) reticulogranulate ( Figures 1–6 View Figures 1–6 , 10–13 View Figures 10–13 , 21 View Figures 19–21 and 23 View Figures 22–25 ; figures 3e and 5e in Garcete- Barrett [2003]); (1) smooth ( Figures 19, 20 View Figures 19–21 and 22 View Figures 22–25 )
(19) Mesoscutal micropunctation 1: (0) abundant ( Figure 11 View Figures 10–13 ; figure 3e in Garcete-Barrett [2003]); (1) scattered ( Figure 12 View Figures 10–13 ; figure 5e in Garcete- Barrett [2003])
(20) Mesoscutal micropunctation 2 (applicable only to terminals that received state 0 for Character 19): (0) present throughout; (1) absent meso-anteriorly ( Figure 11 View Figures 10–13 )
(21) Tegula: (0) rounded ( Figures 10–13 View Figures 10–13 ; figures 2f, 3g, 4f, 5h, 6e, 7g and 8j in Garcete-Barrett [2001]); (1) campanulate (figure 1j in Garcete-Barrett [2001])
(22) Outer margin of tegula: (0) expanded, hiding basalar sclerites ( Figures 10–13 View Figures 10–13 ); (1) not expanded, not hiding basalar sclerites
(23) Basal angle of second submarginal cell: (0) acute (figures 5a, 5b and 5c in Garcete-Barrett [2003]); (1) straight (figure 5d in Garcete-Barrett [2003])
(24) Second submarginal cell: (0) not petiolate (figures 5a, 5b and 5c in Garcete- Barrett [2003]); (1) petiolate (figure 5d in Garcete-Barrett [2003])
(25) Metanotum: (0) cristate ( Figures 27 and 30 View Figures 26–30 ); (1) not cristate ( Figures 26, 28 and 29 View Figures 26–30 )
(26) Crests of metanotum (applicable only to terminals that received state 0 for Character 25): (0) slight (figure 1g in Garcete-Barrett [2003]); (1) strong ( Figures 27 and 30 View Figures 26–30 ; figures 2f, 3j, 4h, 5h and 5g in Garcete-Barrett [2003])
(27) Propodeum dorsum: (0) with median shelf ( Figures 26, 28–32 View Figures 26–30 View Figures 31–34 ); (1) without median shelf ( Figure 27 View Figures 26–30 )
(28) Concavity of posterior surface of propodeum: (0) enlarged, covering the entire face ( Figures 26–28, 30–32 View Figures 26–30 View Figures 31–34 ); (1) reduced to median lower area ( Figure 29 View Figures 26–30 )
(29) Propodeum 1: (0) with submedian propodeal carina ( Figures 26–28 and 30 View Figures 26–30 ); (1) without submedian propodeal carina ( Figure 29 View Figures 26–30 )
(30) Propodeum 2: (0) with lateral propodeal carina (figures 1k and 2h in Garcete-Barrett [2001]); (1) without lateral propodeal carina (figure 3i in Garcete-Barrett [2001])
(31) Propodeal valvula: (0) rounded ( Figure 24 View Figures 22–25 ); (1) square ( Figure 25 View Figures 22–25 )
(32) Tergum I: (0) with transverse carina ( Figures 14–16 View Figures 14–18 , 25 View Figures 22–25 , 33 and 34 View Figures 31–34 ); (1) without transverse carina
(33) Disc of tergum I: (0) with transverse hump ( Figures 14 and 16 View Figures 14–18 ; figure 5e in Garcete-Barrett [2001]); (1) without transverse hump
(34) Sternum II transverse furrow: (0) wide (figures 7e and 7f in Garcete-Barrett [2001]); (1) narrow (figures 3e and 3f in Garcete-Barrett [2001])
Table 1 presents a character matrix for the cladistic analyses of Cephalastor . The two most parsimonious cladograms were obtained under equal weights (strict consensus shown) and one under an implied weighting scheme ( Figures 35 View Figure 35 and 36 View Figure 36 ). The trees were rooted between the clade Hypancistrocerus reflectorius ( Parancistrocerus areatus , Stenodynerus scabriusculus ( Stenodynerus convolutus , Stenodynerus suffusus )) and the remainder of the terminals in order to investigate a possible paraphyletic condition of Ancistroceroides and its relation with Cephalastor .
Despite the main goal of the analyses being to evaluate the character evolution and relationships among species of Cephalastor , some considerations about the outgroup are quite important as well. A clade composed by Hypancistrocerus , Parancistrocerus and Stenodynerus was recovered. A closer relation between Parancistrocerus and Stenodynerus was already observed by Carpenter and Cumming (1985) by the presence of foveae on the anterior pronotal surface, while Hypancistrocerus was not included in their analysis.
Ancistroceroides was not supported as monophyletic based on the species included in this study. The consensus tree under equal weights recovered a polytomy among Ancistroceroides conjunctus , A. rufimaculus and Cephalastor . Under the implied weighting scheme, A. rufimaculus was sister to the clade Cephalastor . Despite this small difference, we believe the cladogram in which the homoplasies were downweighted (implied weighting scheme; Figure 36 View Figure 36 ) is more stable and better supported ( Goloboff et al. 2008), since a better indication to understanding the relationship within Ancistroceroides sensu lato and with related genera is expressed therein. In fact, the morphological heterogeneity of this genus is evident and might prove the paraphyly of the taxon, although further investigation is needed.
The relationships within Cephalastor and the character state distribution were stable under both weighting schemes, except for some autapomorphies of C. sinusiticus . The monophyly of Cephalastor is supported in this study by five synapomorphies and one homoplasy as follows: the cephalic foveae located between the compound eyes (Char. 4 [0]), the presence of interocellar tubercles (Char. 9 [0]), the modified posterior declivity of the head (Char. 11[1]), the punctate anterior pronotal surface (Char. 15 [0]), the reticulo-granulate background microsculpture of the head and dorsal mesosoma (Char. 18 [0]), and the transverse hump on disc of tergum I behind the transverse carina (Char. 33 [0]). The presence of punctures on the anterior surface of the pronotum is a character state shared with several species of Stenodynerus and which was considered by Carpenter and Cumming (1985) as a synapomophy for their Stenodynerus -Microdynerus component.
The topology obtained in the present analysis differs little from that of Garcete- Barrett (2003), with C. sinusiticus being sister to the remaining Cephalastor species. C . bossanova and C. estela were not recovered as sister species as previously done. The new species C. humeralis and C. minarum were placed in an intermediary position in the cladogram in an unresolved politomy with a larger clade comprised by eight species ( C. mariachi , C. rominae , C. rufosuffusus , C. chasqui , C. paezi , C. abraham , C. relativus and C. tupasy ). This large clade is supported by two synapomorphies and has a Bremer support value of 2, and the same was also recovered in the analysis of Garcete-Barrett (2003). Most of this clade remains unresolved, despite for C. chasqui and C. paezi which were recovered as sister species and C. abraham that came out as sister to C. relativus + C. tupasy .











| DZUP |
Universidade Federal do Parana, Colecao de Entomologia Pe. Jesus Santiago Moure |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |