
Orrichthys longimanus, Carnevale & Pietsch, 2010
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2009.00623.x |
|
DOI |
https://doi.org/10.5281/zenodo.10545591 |
|
persistent identifier |
https://treatment.plazi.org/id/2816A226-775A-9653-42E1-1F77FF307FD4 |
|
treatment provided by |
Valdenar |
|
scientific name |
Orrichthys longimanus |
| status |
sp. nov. |
† ORRICHTHYS LONGIMANUS SP. NOV. ( FIGS 5–7 View Figure 5 View Figure 6 View Figure 7 )
† Histionotophorus bassani Sorbini, 1972 : pl. 6, fig. 1 (misidentification).
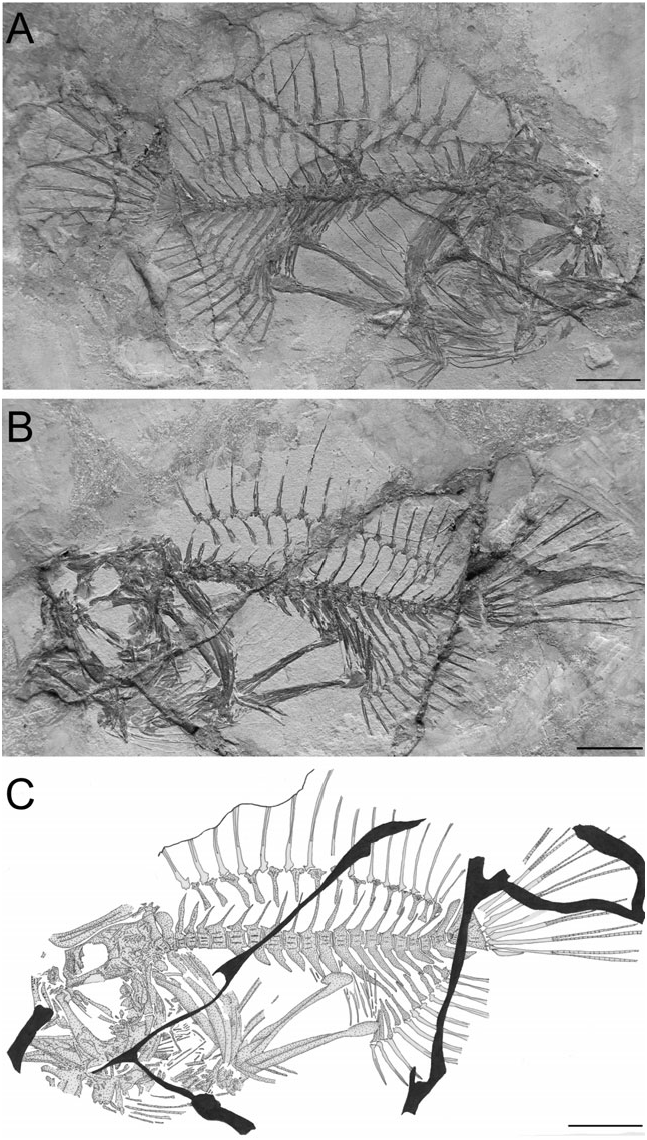
Holotype: MCSNV T.160/161, nearly complete skeleton in part and counterpart, 63 mm SL; late early Eocene , Ypresian ; Monte Bolca, Pesciara cave site.
Paratype: MCSNV T.164/165, nearly complete skeleton in part and counterpart, 54.1 mm SL; from the type locality .
Diagnosis: As given for the genus.
Description: The body is short and globose (depth as great as 52.5% SL) ( Fig. 5 View Figure 5 ). The caudal peduncle is short and deep. The head is compressed, relatively large (about 37% SL). The snout is short, the orbital diameter moderately large. The neurocranium is slightly oblique in position. The mouth is nearly horizontal and extremely large ( Figs 5 View Figure 5 , 6 View Figure 6 ). There is a membrane between the second and third dorsal-fin spines and between the third spine and the dorsal mid-line of the body. The dorsal-fin rays are broadly elongate. The external margins of the dorsal and anal fins are characterized by a gently curved profile. The arm-like pectoral fin is supported by greatly elongate pectoral radials. The pelvic-fin rays are also relatively long (24.9% SL). The caudal fin is rounded. Additional counts and measurements are given in Table 2.
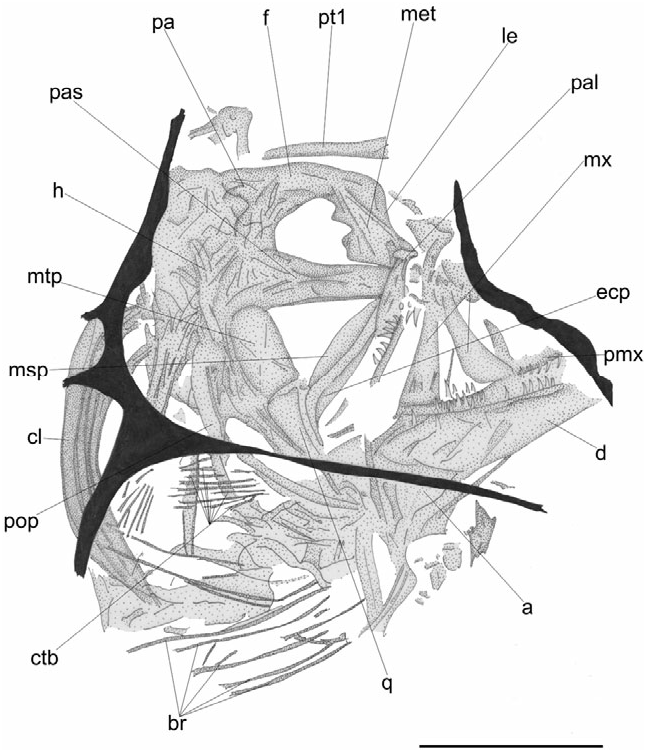
The neurocranium is massive, moderately high, and subrectangular in outline ( Figs 5 View Figure 5 , 6 View Figure 6 ). The bones are well ossified and thickened ( Fig. 6 View Figure 6 ). The frontals All values are in millimetres. Values in parentheses are in per cent of standard length.
are large and robust. The parietal appears to be characterized by a nearly rounded profile; the parietals are separated from each other by the supraoccipital. The mesethmoid is thick, with an irregular posterior profile. What appear to be the lateral eth- moids partially cover the anterior portion of the mesethmoid. The bones of the ethmoid block are separated from the basicranial elements by a very small gap, which was probably filled originally by the ethmoid cartilage. The vomer is toothless, with a flattened ventral surface. The parasphenoid is extremely thick, dorsoventrally expanded, with a median lateral ridge. The bones of the otic and occipital regions are difficult to determine.
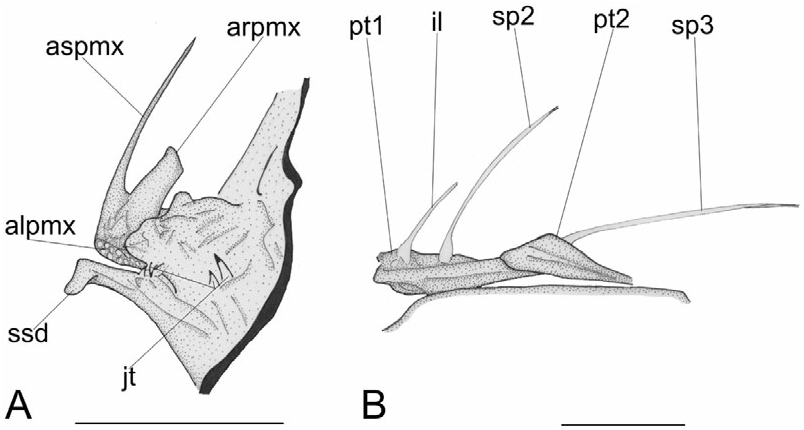
The premaxilla bears a slender elongate ascending process and a flattened spatulate articular process ( Fig. 7A View Figure 7 ). There are a few rows of large depressible caniniform teeth with recurved tips. The maxilla is greatly elongate and robust, with an enlarged flattened posterior end, its length contained approximately five times in SL (see Table 2). The dentary is massive and thick, with a prominent symphysial spine; the mandibular teeth are identical to those of the upper jaw ( Fig. 7A View Figure 7 ). The articular bears a posteroventral bony lamina, with a rounded profile.
The bones of the suspensorium ( Fig. 6 View Figure 6 ) are fragmented in both specimens examined. The hyomandibula has a slender primary shaft, two articular heads, and a short opercular process. The quadrate is roughly triangular and dorsoventrally expanded. The symplectic is difficult to recognize. The metapterygoid is flat and nearly rectangular in shape, with a convex dorsal margin and two diagonal ridges. The ectopterygoid is greatly elongate, crescent-shaped, and posteriorly expanded. The endopterygoid consists of an elongate thin plate, with a rounded dorsal profile. The palatine has an enlarged robust articular head.
What appears to be the preopercle is elongate and slightly curved ( Fig. 6 View Figure 6 ). The opercle cannot be determined. The subopercle is crescent-shaped, apparently without a spine along the anterior margin.
The hyoid bar is difficult to recognize as a result of inadequate preservation. There are five slender and elongate branchiostegal rays ( Fig. 6 View Figure 6 ). The bones of the branchial skeleton are badly fragmented. The ceratobranchials appear to be slightly curved. Large caniniform pharyngobranchial teeth are recognizable.
The vertebral column is slightly curved ( Fig. 5 View Figure 5 ). There are 21-22 vertebrae, 12-13 abdominal and nine caudal. The anterior-most vertebra is strongly associated with the occipital region of the neurocranium. With the exception of the anterior-most four vertebrae, which are nearly square in shape, the centra are massive, subrectangular, and higher than long. The four anterior-most vertebrae bear anteroposteriorly enlarged neural spines; the two anteriormost are characterized by having a rounded profile, whereas those of the third and fourth are stout and pointed. The neural spines of the fifth to eighth vertebrae are shorter, not clearly interdigitating with the dorsal-fin pterygiophores situated above. Welldeveloped neural prezygapophyses characterize all the vertebral centra. Vertebrae 7 (8) to 20 (21) bear flattened, anteroposteriorly enlarged and posteroventrally directed haemal spines, each with a pointed tip ( Fig. 5C View Figure 5 ).
The penultimate vertebra bears enlarged spatulate neural and haemal spines. The hypural plate is roughly triangular, with a small median notch along its posterior margin ( Fig. 5C View Figure 5 ). There are no epurals. The caudal fin consists of nine rays, four in the upper lobe and five in the lower one. The lowest ray is reduced to a small splint of bone; the remaining rays are bifurcated distally.
The spinous dorsal fin is well preserved in the paratype ( Fig. 7B View Figure 7 ); the illicium and two following spines are slender. The first pterygiophore is greatly enlarged; it consists of an elongate anteriorly expanded bony lamina, with a thick median ridge. The second pterygiophore is rather large, blade-like, with an elongate lateral bony crest. The soft dorsal fin contains 16–17 rays, supported by 15–16 pterygiophores ( Fig. 5 View Figure 5 ). These pterygiophores are characterized by having an elongate primary shaft and an enlarged, fan-like distal end. The four anterior pterygiophores are obliquely orientated with posteroventrally directed shafts. The two posterior pterygiophores lie in the interneural space between the 17th (or 18th) and 18th (or 19th) vertebrae. The posteriormost dorsal-fin pterygiophore (as well as its counterpart in the anal fin) bears a posteriorly directed process on the posterior margin of its distal end.
The anal fin consists of 11 rays supported by ten pterygiophores ( Fig. 5 View Figure 5 ). The proximal shaft of the two anterior pterygiophores is bent posteriorly. The four anterior pterygiophores lie in the interhaemal space between the 17th (or 18th) and 18th (or 19th) vertebrae.
The pectoral fin inserts above the anal-fin origin ( Fig. 5 View Figure 5 ). It contains eight or nine elongate rays. The supracleithrum is elongate and laterally flattened. The cleithrum is rather large and crescent shaped. The scapula and coracoid are poorly preserved and difficult to interpret. There is a single elongate postcleithrum. The two radials are strongly elongate; the ventral-most has a distally expanded portion and bears the bases of the pectoral-fin rays. The pelvic fin contains one spine and five rays ( Fig. 5A View Figure 5 ). The basipterygium is stout with an expanded distal end. The skin is naked.
Etymology: The specific name is derived from the Latin longus, meaning ‘long’; and manus, ‘hand,’ in allusion to the exceptionally long, hand-like, pectoralfin lobe of this species.
Remarks: The first documentation of this new handfish genus and species was provided by Sorbini (1972), who figured the holotype ( MCSNV T.161) but misidentified it as † Histionotophorus bassani . The information obtained in this study indicates that † Orrichthys longimanus is defined by three autapomorphies, plus many other features that are found in various combinations in other brachionichthyid or antennarioid fishes. Some of these diagnostic features, including large caniniform jaw teeth, an enlarged maxilla, dorsoventrally developed anterior part of the body, and anteroventrally directed anterior-most anal-fin pterygiophores, can all be considered trophic adaptations. In particular, these characters clearly reflect an increased development of the oro-pharyngeal cavity (i.e. an expansion of the throat and abdomen) for engulfing large prey. Although a similar strong development of feeding adaptations is not evident in † Histionotophorus bassani , it should be noted that the mouth and abdomen of the latter are still comparatively enlarged with respect to those of extant brachionichthyids. Therefore, the microphagous habits characteristic of extant handfishes can be interpreted as a specialization amongst lophiiforms, whereas the functional adaptations of the trophicrelated structures of † Orrichthys longimanus and, to a lesser extent, of † Histionotophorus bassani , seem to be consistent with those of other antennarioids, and more generally of other lophiiform fishes (e.g. see Gregory, 1933; Gregory & Conrad, 1936; Pietsch, 2009).
| T |
Tavera, Department of Geology and Geophysics |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Orrichthys longimanus
| Carnevale, Giorgio & Pietsch, Theodore W. 2010 |
Histionotophorus bassani
| Sorbini 1972 |