
Bolbites onitoides, HAROLD, 1868
|
publication ID |
https://doi.org/ 10.1093/zoolinnean/zlab015 |
|
publication LSID |
lsid:zoobank.org:pub:42E7A370-FCD8-492A-8604-C42863BDE6E2 |
|
DOI |
https://doi.org/10.5281/zenodo.6354401 |
|
persistent identifier |
https://treatment.plazi.org/id/27085036-DC08-FFE5-3ECE-08B8DE29F8A1 |
|
treatment provided by |
Plazi |
|
scientific name |
Bolbites onitoides |
| status |
|
BOLBITES ONITOIDES HAROLD, 1868 View in CoL View at ENA
( FIGS 1–15 View Figure 1 View Figure 2 View Figure 3 View Figure 4 View Figure 5 View Figure 6 View Figure 7 View Figure 8 View Figure 9 View Figure 10 View Figure 11 View Figure 12 View Figure 13 View Figure 14 View Figure 15 )
Bolbites onitoides Harold, 1868a: 81–82 View in CoL ; Harold, 1869a: 1016; Bates, 1870: 174; Steinheil & Strobel, 1872: 555; Burmeister, 1874: 129; Fabre, 1899: 73, 75–77; Judulien, 1899: 376, figs 5, 7; Laloy, 1902: 171; Kolbe, 1905: 487; Bruch, 1911: 189; Gillet, 1911: 80; Fabre, 1919: 242–244; Lucas, 1920: 141; d’Olsoufieff, 1924: 20, 58, 136, 138, pl. 3, figs 1, 10; Janssens, 1940: 1; Balthasar, 1941: 350; Pessôa & Lane, 1941: 470–471, fig. 32, pl. 10, fig. 75; Blackwelder, 1944: 208; Balthasar, 1951: 335; Martínez, 1959: 94; Halffter & Matthews, 1966: 132; Krantz, 1967: 12, 48, 53, 55, 86–87; Haffter & Matthews, 1971: 159; Edmonds, 1972: 818, 854; Halffter & Edmonds, 1982: 86, 89; Kirk, 1992: 53; Luzzatto et al., 1994: 63; Cabrera Walsh & Gandolfo, 1996: 582–587, fig. 1i; Monteresino et al., 1996: 109; Morelli & Gonzalez-Vainer, 1997: 197; Morelli et al., 2002: 54; Philips et al., 2004b: 46, 56, 58–59, 62, figs 2, 18, 30; Hamel-Leigue et al., 2006: 6, figs 41–42; Krajcik, 2006: 16; Ocampo & Hawks, 2006: 562, 564–567, figs 5–8; Hamel-Leigue et al., 2009: 49, 56, 69, 85, fig. 8, photos 1–2; Scholtz, 2009b: 254–255, figs 12.6a–b; Vazde-Mello & Grossi, 2010: 220; Vaz-de-Mello et al., 2011: 44, 51, fig. 22; Krajcik, 2012: 54; Cupello & Vaz-de-Mello, 2013: 441–442; Cupello & Vaz-de-Mello, 2015: 16, 18–19, figs 7C, 8C; Alonso et al., 2019: 299; Cabrera Walsh, 2020: 761; Gillett & Toussaint, 2020: 665, 667, 670, 673–674, figs 3–4.
Bolbites onithoides [incorrect subsequent spelling]: Di Iorio & Turienzo 2015: 21.
Type series: Lectotype: Male, here designated (1, Harold’s handwriting, bordered in red, ‘ Montevideo B. onitoides t. Harold’; 2, bordered in black, ‘Ex-Musaeo E. Harold’; 3, red paper, ‘LECTOTYPE’; 4, green paper bordered in black, ‘Muséum Paris ex Coll. R. Oberthür 1952’; 5, MC’s handwriting, red paper, ‘ LECTOTYPE Ƌ Bolbites onitoides Harold des Cupello & Vaz-de-Mello, 2019’), MNHN – ex Edgar von Harold and R. Oberthür collections. Potential paralectotypes (see Remarks below): 1. male (1, bordered in black, ‘Ex-Musaeo E. Harold’; 2, green paper bordered in black, ‘Muséum Paris ex Coll. R. Oberthür 1952’), MNHN – ex Edgar von Harold and R. Oberthür collections. 2. female (1, bordered in black, ‘Ex-Musaeo E. Harold’; 2, green paper bordered in black, ‘Muséum Paris ex Coll. R. Oberthür 1952’), MNHN – ex Edgar von Harold and R. Oberthür collections.
Type locality: Uruguay: Montevideo .
Etymology: The specific name onitoides , a noun in the nominative case used in apposition, translates into English as ‘the one like Onitis ’ and refers to the overall similarity of B. onitoides to members of the genus Onitis . It is formed from the combination of the name Onitis plus the suffix -oides, the latter derived from the combination of the linking vowel - o - and the Ancient Greek noun εἶδος (eidos) for ‘form’ or ‘likeness’ ( Brown, 1956; Papavero, 1994; Wiktionary, 2020). Our interpretation of the name as being a noun instead of as an adjective follows Article 31.2.2 of the Code. According to Harold (1869a), Fabricius’s (1798) genus name Onitis derives from a Greek word spelled by Harold as ὄνἰς and translated into Latin as ‘ stercus asininum ’ (i.e. ass dung). Brullé (1838), in contrast, states the name is derived from the Greek word ὀνῖτις (onitis), which he said refers to an unspecified plant name. Greek dictionaries and lexica indeed list ὀνῖτις as a feminine noun and say it refers to acrid herbs such as marjoram, Origanum L. ( Liddell & Scott, 1897; Brown, 1956; Wiktionary, 2020). The word suggested by Harold, on the other hand, is not found with that spelling in Greek dictionaries, but the similar form ὄνος (ónos) is present and indeed translates as ‘ass’ or ‘donkey’ ( Liddell & Scott, 1897; Wiktionary, 2020). If Harold is correct, then it is possible that Fabricius (1798) derived Onitis from the combination of the stem of ὄνος (viz., ὄν–, on -) plus the suffix -ῖτις (- itis), the feminine equivalent of -ῑ́της (-ites). Under either Harold’s or Brullé’s suggestion, Onitis would be feminine in gender, which disagrees with the way Fabricius (1798, 1801) and (as far as we know) all other authors have ever treated the name, i.e. as masculine, including equivalent names in botanical nomenclature ( IPNI, 2021).
Diagnosis: Bolbites onitoides is readily identified as a Phanaeini by the presence of iridescent colouration, eyes completely divided by the canthus into a dorsal and a ventral portion, a long metanepisternal tab ( Fig. 5H View Figure 5 ), and its metaventrite with an acuminate anteromedian prominence flanked by long setae ( Fig. 4A View Figure 4 ). In turn, it differs from all other phanaeines by having meso- and metatarsi with claws and from other Phanaeina by its antennae with free apical lamellae. Its unusual lack of dorsal sexual dimorphism while having a rich ventral one is also an unique condition among the Phanaeina.
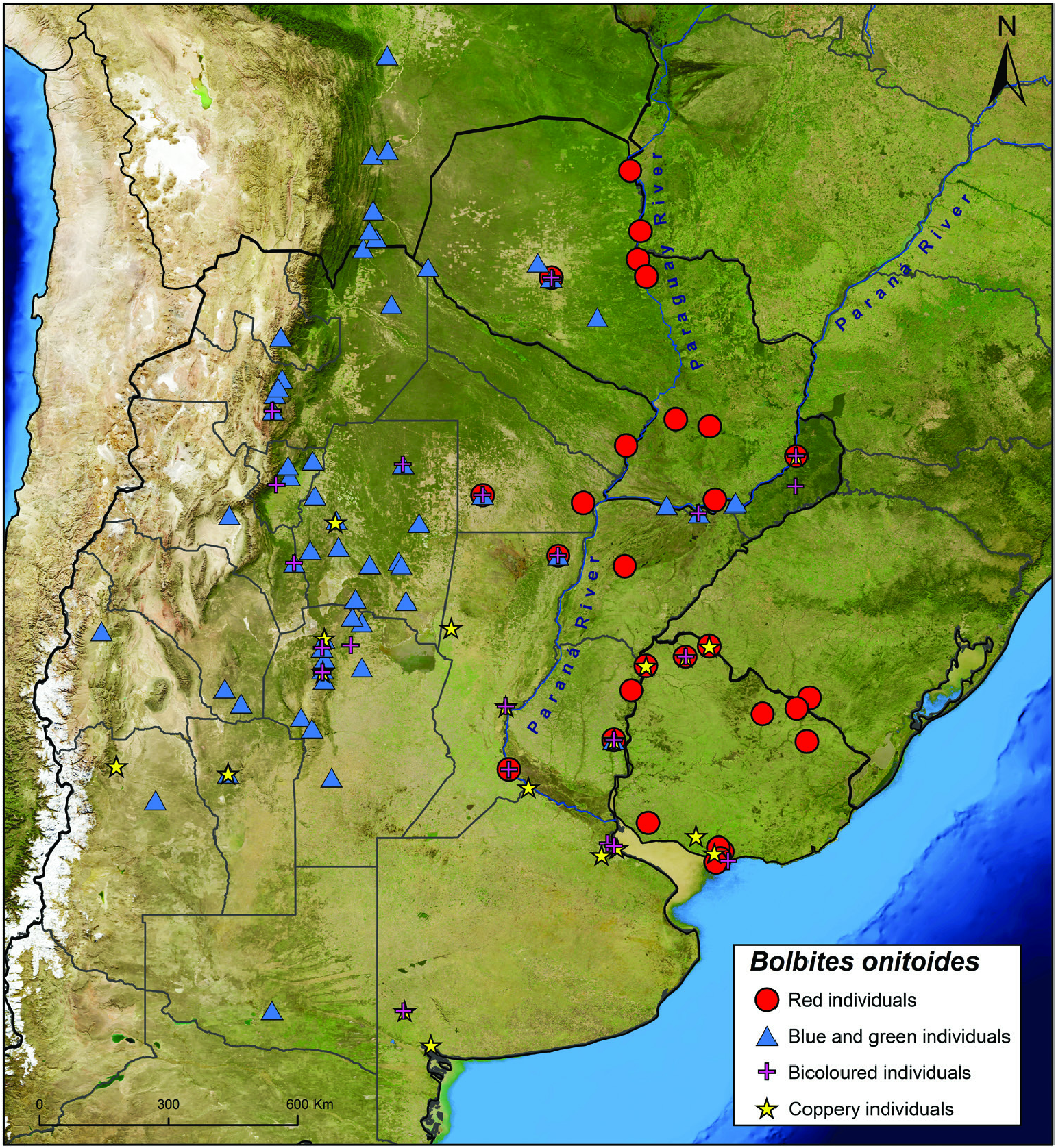
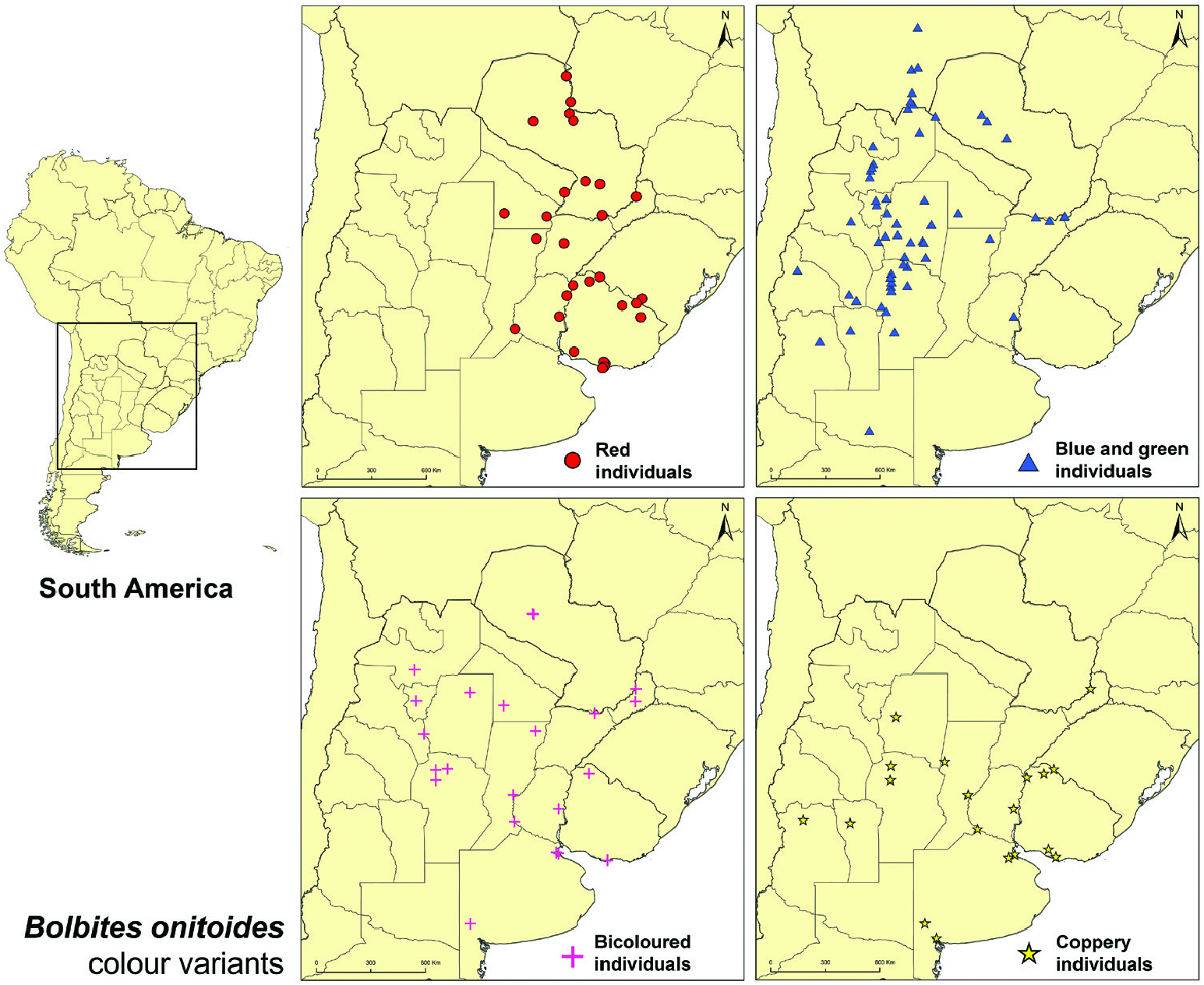
Redescription: Colouration: Clypeus sexually dimorphic (see below). Frons, paraocular areas, pronotum and elytra mainly metallic light to dark blue in western populations ( Fig. 1G, H View Figure 1 ) and metallic red in eastern populations ( Fig. 1B View Figure 1 ), both phases with different degrees of greenish reflections over dorsum; throughout the range, specimens sometimes with metallic red head and pronotum and metallic blue or green elytra ( Fig. 1D–F View Figure 1 ) or dorsum entirely metallic coppery ( Fig. 1A View Figure 1 ). Venter, including ventral surface of legs, and pygidium always ranging from dark blue to light green, never red or copper ( Fig. 2B, C View Figure 2 ); centre of metaventrite sexually dimorphic (see below).
Head: Clypeus at anterior edge with two small, rounded teeth separated by a broad U-shaped emargination; anterior edge only slightly bent upwards; integument entirely covered by net of transverse ridges; clypeal process a rounded transverse carina. Junction of clypeus and paraocular areas deeply notched on outer edge. Paraocular areas and frons with integument similar to clypeus, but ridges less well-formed and, posteriorly, sometimes becoming individual granules. Cephalic process raised between clypeus and frons into a straight transverse carina ( Fig. 3 View Figure 3 ); carina usually with three (smaller specimens) or five (larger specimens) tenuous elevations, the central one higher than the lateral ones, elevations never acute or with aspect of tubercles or teeth such as those of many Coprophanaeus s.s. (in worn specimens, carina truncate, without elevations). Eyes completely divided into dorsal and ventral portions by prolongation of canthus; dorsal portion slightly smaller than ventral portion.
Thorax: Pronotum on anterior region with transverse C-shaped ridge flanked by shallow depressions ( Fig. 3 View Figure 3 ); pronotal ridge at most as wide as cephalic carina ( Fig. 3 View Figure 3 ); without horns or any other ornamentation in both sexes ( Figs 2A View Figure 2 , 3 View Figure 3 ); integument almost entirely sculptured, postero-central region usually with small rounded impressions that become progressively more confluent towards anterior region, giving a rugose aspect to integument; anterolateral angles usually covered by shiny scale-like granules; lateral margin irregularly serrate, with sexually dimorphic presence of posterior tab; anterior margin not interrupted behind eyes; posterior margin complete; lateral fossae deep, broad, well developed, smooth and shiny centrally; with two deep, well-developed, elongate posterior fossae. Hypomera, mesepimera, metanepisterna and outer lobes of metaventrite covered by abundant long, black setae. Metanepisterna with long dorsal tab covering part of epipleura, which is evidently depressed at this point ( Fig. 5H View Figure 5 ). Mesoventrite constricted at centre by anterior projection of central lobe of metaventrite ( Fig. 4A View Figure 4 ). Metaventrite with well-developed anteromedian prominence with fairly acute apex and flanked by long and dense black setation, laterals without granules typical of Gromphas ( Fig. 4A View Figure 4 ); in lateral view, anteromedian prominence vertical anteriorly, not projected, at most only slighly indicated apically ( Fig. 2A View Figure 2 ); centre of metaventrite glabrous and covered by sexually dimorphic ill-delimited punctation, never smooth, some specimens with tenuous, ill-delimited mid-longitudinal groove.
Legs: Shape of profemora, protibiae, protibial spur, protibial ventral carina, metacoxae, metafemora and metatibiae sexually dimorphic (see below). All femora with long, black setae ( Figs 5A, B View Figure 5 , 6C, D View Figure 6 ); setae denser on profemora than on meso- and metafemora. Profemora with small acute tubercle on ventral surface ( Fig. 5A, B View Figure 5 ). Protibiae with four external teeth ( Fig. 5C, D View Figure 5 ); ventral surface covered by series of oblique ridges. Protarsi absent in both sexes. Meso- and metatarsi with five tarsomeres and short, curved tarsal claws ( Fig. 5F, G View Figure 5 ); tarsomere I to IV progressively smaller; tarsomere V (apical) longer than IV and with strong apical projection rising from beneath tarsal claws ( Fig. 5F View Figure 5 ). Metacoxae usually with long setae at apex ( Fig. 6A, B View Figure 6 ); sometimes setae absent due to abrasion (but setiferous punctures still visible over integument surface) or surface naturally glabrous; density of setation sexually dimorphic (see below). Metatibial spur gently curved apically.
Elytra: With nine thin, ill-delimited striae, sometimes almost impossible to discern from the rest of integument except on bright humeri and apical calluses; ninth stria adjacent to epipleura. Integument with diffuse shine by being mostly covered by foveolate microsculpture (microfoveae only clearly discernible from one another at 70× magnification or more); bright, smooth areas limited to sutural margin, humeri and apical calluses; no granules or irregularities of any kind. Sutural margin elevated, highly shiny and smooth or with sparse micropunctures; apex sexually dimorphic. Humeri projected, usually entirely punctate and shiny. Apical calluses usually punctate and shiny. Base of elytra deeply depressed between third stria and humerus. Epipleura broad, with integument similar to most of rest of elytra.
Abdomen: With six ventrites. Ventrite I acute between apices of metacoxae ( Fig. 6A -B View Figure 6 ). Ventrites I–V glabrous and with integument covered by thin alveolar microsculpture (alveoli only clearly discernible at 80× magnification); ventrite VI with similar integument on laterals and smooth at centre. Ventrite VI narrow, much shorter than the others, as wide in males as in females. Pygidium with integument entirely coated by thin alveolar microsculpture similar to that of ventrites and by small, rounded impressions, not usual punctures ( Fig. 7 View Figure 7 ); basal margin complete, never interrupted by midlongitudinal groove of propygidium, sexually dimorphic (see below).
Genitalia: Genital segment: Mostly membranous ( Fig. 8B View Figure 8 ). Medial sclerite paired and elongate. Lateral sclerotized plates strongly sclerotized. Aedeagus: Parameres tapered towards apex in both lateral and dorsal views ( Fig. 8C, D View Figure 8 ); in dorsal view, with conical aspect ( Fig. 8C View Figure 8 ); without hooks or notches; coated dorsally by wide area of whitish, glabrous integument. Phallobase, in ventral view, without median incision on basal edge or mid-longitudinal sulcus. Endophallus with five endophallites as follows ( Fig. 9A, B View Figure 9 ): medial endophallite elongate and bent basally ( Fig. 9E View Figure 9 ); superior right peripheral ( SRP) endophallite elongate, bent apically and slightly variable in overall shape, depending on specimen ( Fig. 9F–H View Figure 9 ); axial (A) and subaxial ( SA) endophallites placed tightly together in resting position ( Fig. 9C View Figure 9 ), A endophallite forked at one end and acuminate at the other, SA endophallite broad and curved; fronto-lateral peripheral ( FLP) endophallite fairly reduced, triangular, scale-like Fig. 9D View Figure 9 . Spermatheca: C-shaped, apical branch slender, tapered and with rounded apex, basal branch broader ( Fig. 8A View Figure 8 ). Spermathecal gland not observed.
Sexual dimorphism: Male: Clypeus anterior-half or one-third dull black and posterior-half or two-thirds iridescent, with same colour as rest of head ( Fig. 3A, B View Figure 3 ); paraocular areas and frons usually almost completely iridescent. Pronotum raised into acute posterior tab on lateral margin ( Fig. 3A, B View Figure 3 ). Metaventrite with punctation at centre more smoothly marked than in females. Anterior leg longer than in females ( Fig. 3A View Figure 3 ), articulation between profemora and protibiae visible from above in pinned specimens mounted in traditional position. Profemora claviform, posterior edge with deep basal emargination followed by acute spine ( Fig. 5A View Figure 5 ). Protibiae long and slender, with tuft of long and dense setae apically ( Fig. 3A View Figure 3 , 5C View Figure 5 ); protibial spur shorter and broader than in females, clearly bent downwards ( Fig. 3A View Figure 3 , 5C View Figure 5 ); ventral carina interrupted by series of acute teeth irregular in size, central tooth larger than others ( Fig. 5E View Figure 5 ). Metacoxae with plate projections bearing long, acute posterior spine near insertion of trochanters ( Fig. 6A View Figure 6 ); setation of apical surface usually shorter and sparser than in females, not rarely even entirely glabrous. Metafemora straight on anterior edge, posterior edge with large rounded ridge ( Fig. 6C View Figure 6 ). Metatibiae with strong ridges on external and internal edges, internal edges with few small spines ( Fig. 6C View Figure 6 ). Sutural margins of elytra with short spines at apex ( Fig. 7A View Figure 7 , yellow arrow). Pygidium raised into pair of long spines at basolateral angles of pygidial margin, foveate on surface near spines ( Fig. 6C View Figure 6 , 7A View Figure 7 ); lateral margin partly interrupted near spines in larger specimens. Female: Clypeus entirely dull black, traces of iridescence at most near small areas adjacent to frons and paraocular areas ( Fig. 3C View Figure 3 ); anterior region of paraocular areas and of frons sometimes also dull black. Pronotum not raised into acute tab on lateral margin ( Fig. 3C View Figure 3 ). Metaventrite with punctation at centre coarser than in males. Anterior leg shorter than in males ( Fig. 3C View Figure 3 ), articulation between profemora and protibiae not visible from above. Profemora entirely broad in females, without posterior emargination or spine ( Fig. 5B View Figure 5 ). Protibiae broader and shorter than in males, without well-defined apical tuft of long setae (at most, with few shorter setae on internal apical angle) ( Fig. 5D View Figure 5 ); protibial spur long, spiniform, slightly bent downwards; ventral carina simple, without teeth. Metacoxae unarmed, without plate projection or spines ( Fig. 6B View Figure 6 ); setation of apical surface usually much longer and denser than in males, only rarely glabrous. Metafemora usually (but not always) slightly curved at basal half of anterior edge ( Fig. 6D View Figure 6 ); posterior edge unarmed. Metatibiae with sharp teeth along external edge, but without strong ridge on external and internal edges ( Fig. 6D View Figure 6 ). Sutural margin of elytra unarmed apically ( Fig. 7B View Figure 7 ). Pygidium gently depressed near basolateral angles; basolateral angles not raised into spines, at most slighly elevated; pygidial lateral margin with emargination less evident than in males ( Fig. 7B View Figure 7 ).
Measurements (in millimetres): Males: Total length (TL): 14.8–21.5 [average (AV) = 18.8 ± 1.85]. Partial length (without head; PL): 11.1–17.5 (AV = 15.5 ± 1.48). Maximum width of pronotum ( WP): 7.9–12.7 (AV = 11 ± 1.15). Maximum width of elytra ( WE): 8.2–12.9 (AV = 11.4 ± 1.11). Females: TL: 14.0–20.7 (AV = 17.9 ± 1.93). PL: 11.9–16.7 (AV: 14.4 ± 1.69). WP: 8.6–12.5 (AV = 10.7 ± 1.32). WE: 8.8– 12.6 (AV = 10.9 ± 1.29).
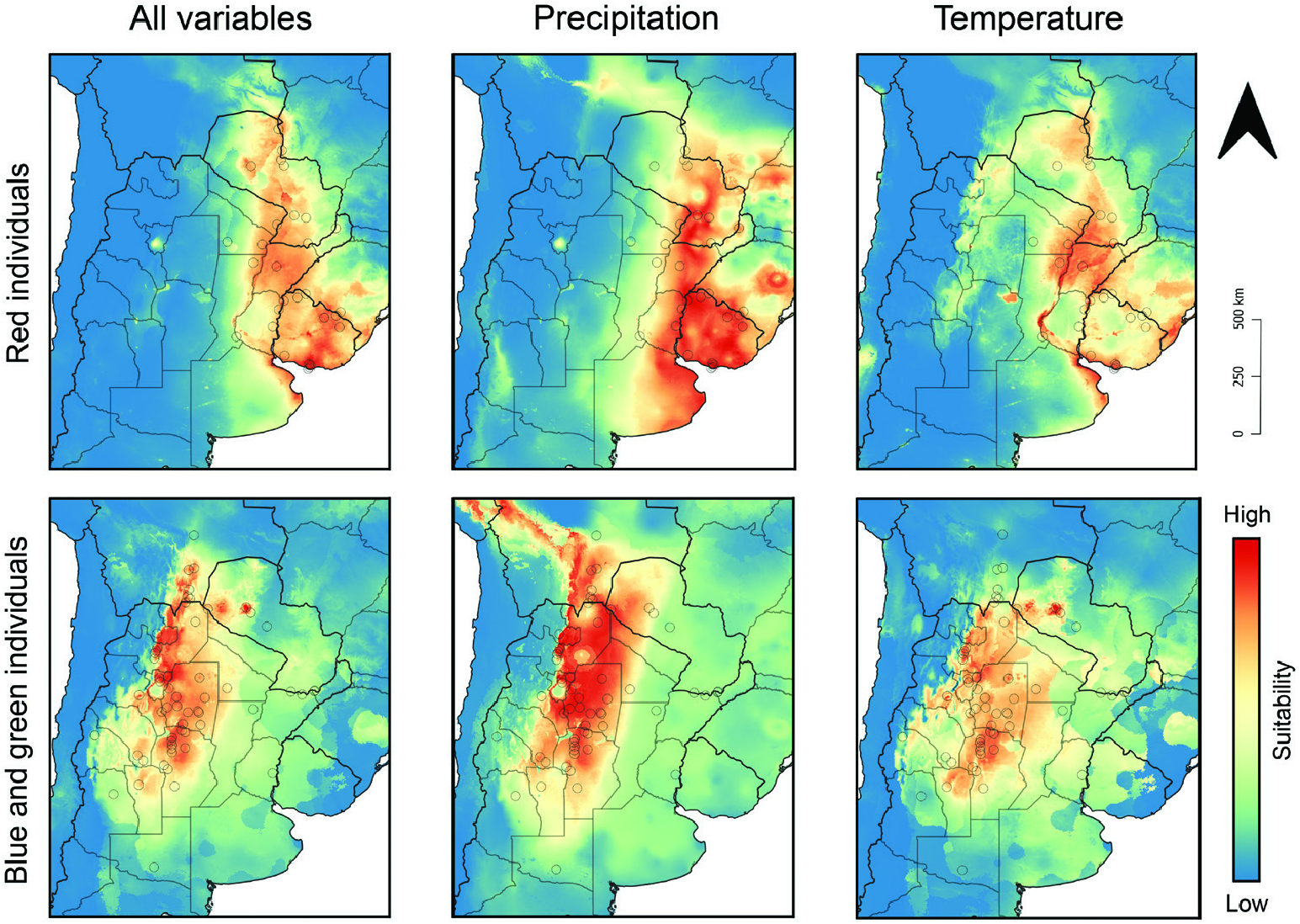
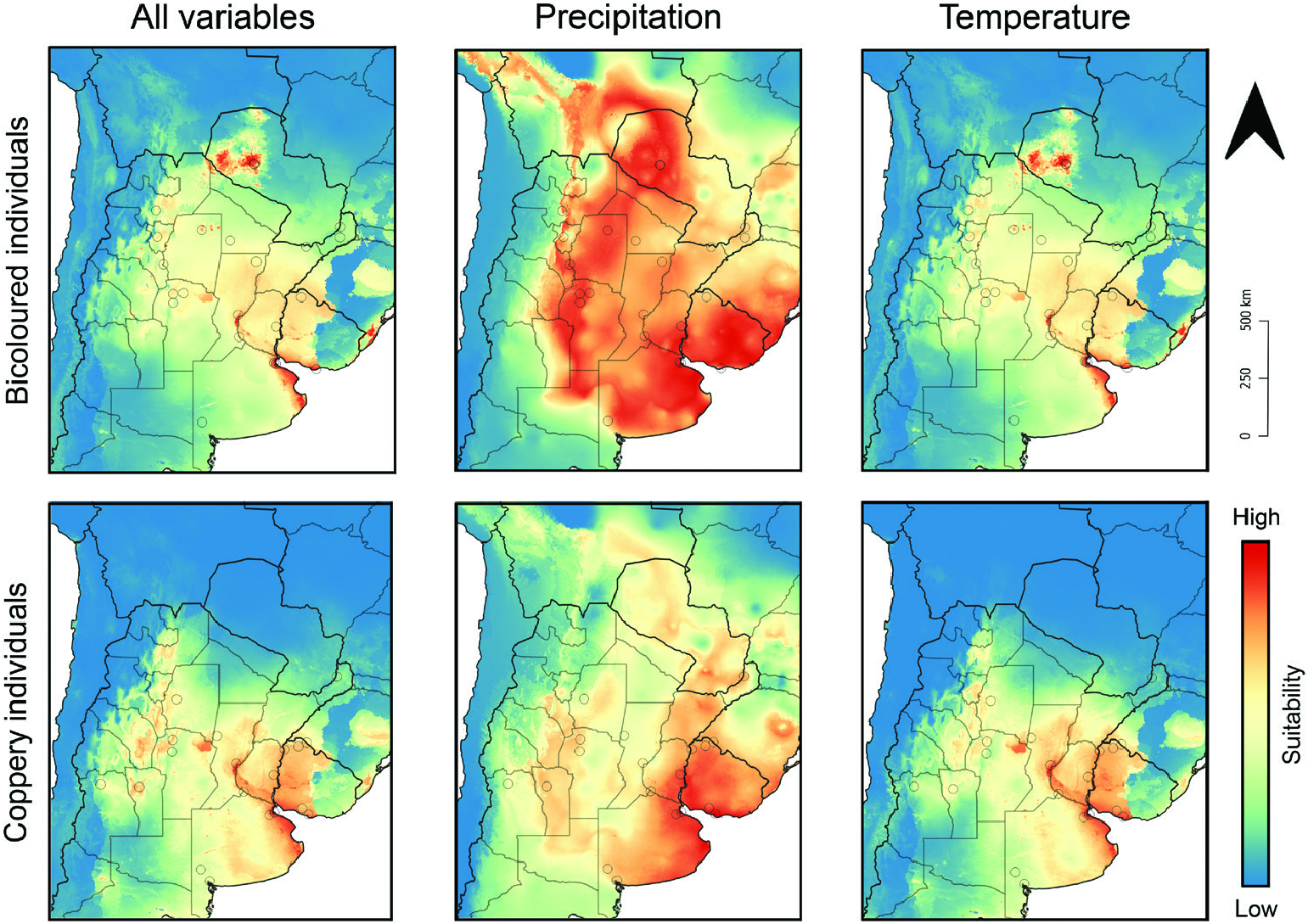
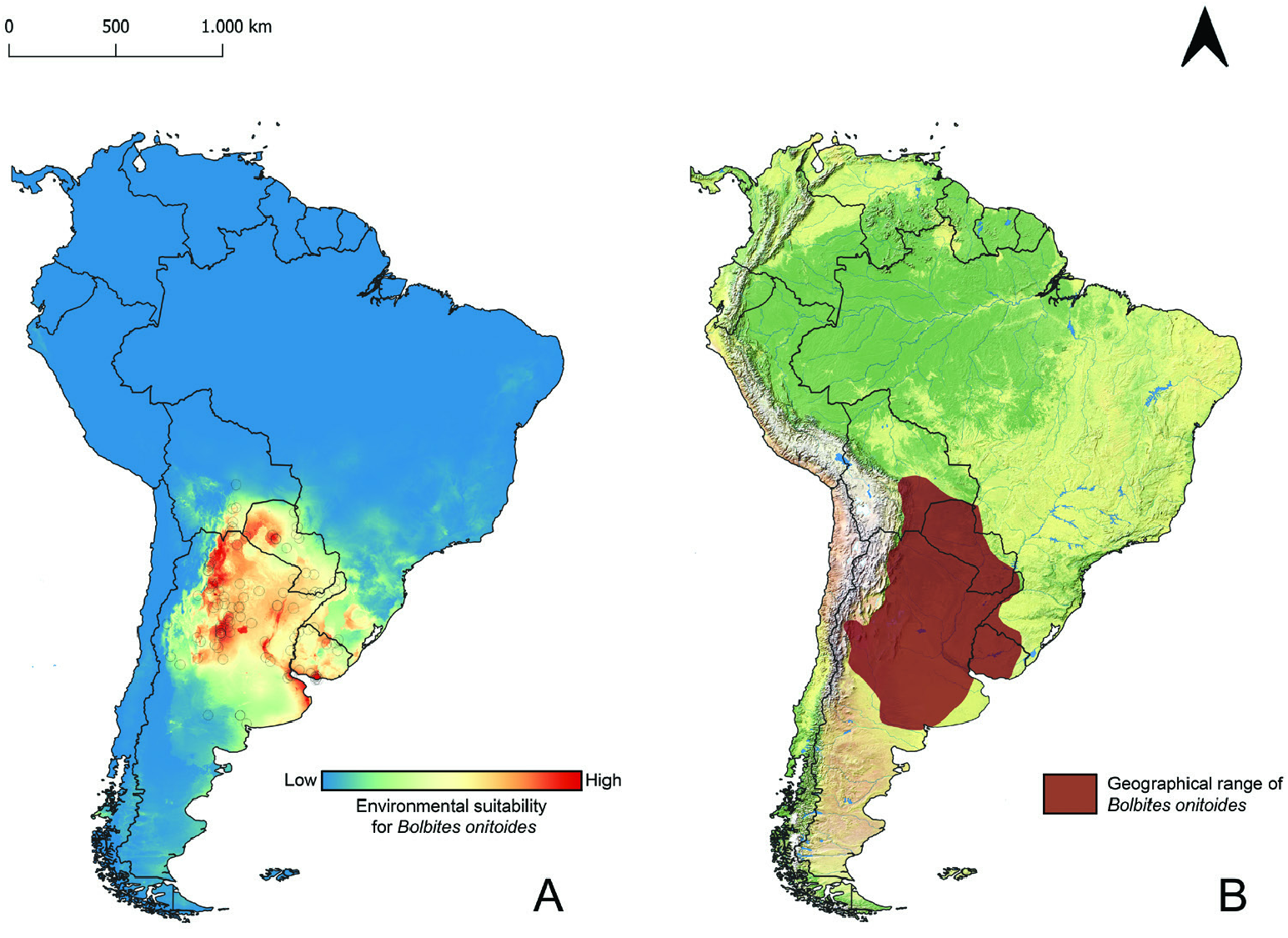
Geographical range: Present mostly across xerophytic deciduous forests, savannas and grass- and shrublands from the dry Chaco in southern Bolivia and northwestern Paraguay and the humid Chaco in eastern Paraguay and Mato Grosso do Sul state in Brazil south to the Uruguayan and southern Brazilian Savanna and the Espinal region in central Argentina. Some populations also present in higher and drier lands along the Argentinian Low Monte region ( Figs 11 View Figure 11 , 12 View Figure 12 ; Supporting Information, Appendix S1).
Biomes and ecoregions: Tropical and subtropical grasslands, savannas, and shrublands: Dry Chaco, Humid Chaco, Uruguayan Savanna. Temperate grasslands, savannas, and shrublands: Humid Pampas, Espinal, Low Monte.
Collecting localities: Red specimens: BOLIVIA [?]. BRAZIL: Mato Grosso do Sul: Porto Murtinho; Rio Grande do Sul: Aceguá, Guaraí . PARAGUAY: Alto Paraguay: Puerto Casado, Puerto Esperanza ; Concepción: Puerto Max; Presidente Hayes: Cruce de los Pioneros; Paraguarí. Guairá: Mbocayaty del Guairá, Villarrica; Itapuá: San Cosme y Damián . ARGENTINA: Formosa: Formosa. Chaco: Chacabuco (Charata), Liberdad ( Puerto Tirol ) ; Corrientes ; Santa Fe: General Obligado (Villa Ana), Rosario ; Entre Ríos: Uruguay (Pronunciamiento) ; Misiones: Eldorado ; Córdoba [?] ; Buenos Aires [?]. URUGUAY: Artigas ; Salto: Belén ; Rivera: Vichadero ; Tacuarembó: Zapucay ; Paysandú: Paysandú ; Cerro Largo ; Colonia ; Canelones: Canelón Chico, Canelones ( Juanicó ) ; Montevideo: Cerro de Montevideo, Playa Punta Yeguas. Blue and green specimens: BOLIVIA: Santa Cruz: Andrés Ibáñez, Cordillera (Camiri, Tatarenda ) ; Tarija: Gran Chaco (El Bagual ; Yacuiba: Yaguacua ; Villamontes ). PARAGUAY: [unspecified department]: ‘ Trans-Chaco’ ; Boquerón: Filadelfia, Fortín Guachalla ; Presidente Hayes: Cruce de los Pioneros, Pozo Colorado . ARGENTINA: Jujuy. Salta: Capital (Salta), Cerrillos (La Merced), Chicoana, General José de San Martín (Embarcación, Salvador Mazza), La Viña ; Chaco: Chacabuco (Charata) ; Misiones: Capital (Posadas), Eldorado (9 de Julio) ; Corrientes: General Paz (Itá Ibaté), Ituzaingó ; Entre Ríos: Uruguay (Pronunciamiento) ; Santiago del Estero: Alberdi (Campo Gallo), Avellaneda (Colonia Dora, Icaño), Banda (La Banda), Barrancas, Choya (Frías, Villa la Punta), Jiménez (El Charco), Loreto, Mitre (Villa Unión), Moreno (Puna), Ojo de Água ; Tucumán: Burruyacú (Burruyacú), Cerro San Javier, Trancas (Tapia) ; Catamarca ; La Rioja: General San Martín (Ulapes), Rosario Vera Peñaloza (Chepes) ; San Juan: Iglesia (Angualasto) ; San Luis: Belgrano ; Córdoba: Calamuchita (Villa Yacanto de Calamuchita), Ischilín (Copacabana, Quilino), Jaime Pérez (Jaime Peter), Punilla (Capilla del Monte, Valle Hermoso), Río Cuarto , Río Seco , San Alberto (Las Rabonas), Sobremonte ( San Francisco del Chañar ), Totoral (La Pampa) ; Santa Fe: General Obligato (Villa Ana) ; Mendoza. La Pampa: Lihuel Calel ( Parque Nacional Lihué Calel ). Bicoloured specimens: PARAGUAY: [unspecified department]: ‘ Trans-Chaco’ ; Presidente Hayes: Cruce de los Pioneros . ARGENTINA: Salta: La Viña. Tucumán ; Santiago del Estero: Alberdi (Campo Gallo), Choya ( Frías ) ; Chaco: Chacabuco (Charata) ; Misiones: Cainguás ( Dos de Mayo ), Eldorado ; Corrientes: Ituzaingó ; Santa Fe: General Obligado (Villa Ana), Rosario , Santa Fe ; Entre Ríos: Uruguay (Pronunciamiento) ; Córdoba: Jaime Pérez (Jaime Peter), Punilla (San Esteban), Tulumba (San José de la Dormida) ; Buenos Aires: Puan (Felipe Solá), San Isidro (Boulogne Sur Mer) ; Ciudad Autónoma de Buenos Aires : Núñez. URUGUAY: Artigas ; Montevideo. Copper specimens: BRAZIL: Rio Grande do Sul: Guaraí . ARGENTINA: Mendoza. San Luis: Belgrano ; Córdoba: Ischilín (Quilino), Punilla (San Esteban) ; Santiago del Estero ; Santa Fe: Rosario, San Cristóbal (Hersilia), Santa Fe ; Misiones: Eldorado ; Entre Ríos: Uruguay (Pronunciamiento) ; Ciudad Autónoma de Buenos Aires ; Buenos Aires: Bahía Blanca , Merlo (Pontevedra), Puan (Felipe Solá), San Nicolás. URUGUAY: Artigas ; Salto: Belén ; San José ; Montevideo: Cerro de Montevideo
.
Intraspecific variation: Besides colour, which will be discussed below, the most noticeable variation observed among individuals of B. onitoides concerns the pronotal punctation. This is most evident over the posterior region of the pronotum near the posterior fossae, where punctures range from being dense and almost contiguous with one another to effaced and sparse; sometimes, the region between the posterior fossae lacks any punctures altogether. Although the latter condition seems to be more common in the eastern part of the distribution, populations from the entire geographical range of the species show individuals varying across the full variation spectrum. Other external characters that show some relevant degree of variation are the apical angle of the anteromedian prominence of the metaventrite, which may be more or less projected forwards and pointed depending on the specimen, and the setation at the apex of the metacoxae, which varies from dense to absent in both sexes (though it tends to be denser and longer in females than in males). Even though a strongly acuminate metaventral angle and a denser metacoxal setation are more commonly observed in western individuals, neither of these characters varies consistently with either distribution or dorsal colour variant, and specimens from all populations may show either extreme of variation.
In regard to genital structures, the endophallus of B. onitoides has one of the most simplified set of endophallites found among the phanaeines, lacking the accessory endophallite that is present in some species of the clade [e.g. Oruscatus davus (Erichson, 1847) , Gromphas aeruginosa and Phanaeus splendidulus (Fabricius, 1781) ; Tarasov & Génier, 2015]. But in spite of this simplification, the information stated in one of our previous works ( Cupello & Vaz-de-Mello, 2015) that B. onitoides does not have a medial endophallite (then called ‘medial sclerite’) is certainly incorrect, as can be clearly seen in Figure 9 View Figure 9 . We do not know the cause of our previous mistake; it is possible that we made wrong notes while dissecting the specimens and the error was repeated in the manuscript. It is worth noting that our observation of the medial endophallite being absent from the two species of Oruscatus Bates, 1870 is indeed correct, as can be confirmed in Tarasov & Génier’s (2015) illustration of the endophallites of O. davus .
The medial endophallite of B. onitoides , despite its overall shape being somewhat conserved among the specimens dissected, shows some minor variations in the development of the lateral projections at its folding area ( Fig. 9E View Figure 9 ). The SRP endophallite, like the medial one, shows some degree of variation, some specimens having one of its extremities bent, giving it a seahorse-like shape in lateral view ( Fig. 9F View Figure 9 ), while others lack such a bend and have a general F-shape in lateral view ( Fig. 9G View Figure 9 ). This variation is correlated with neither population of origin nor dorsal colour. Finally, the A+SA complex ( Fig. 9C View Figure 9 ) and the FLP ( Fig. 9D View Figure 9 ) appear to be the least plastic among the endophallites of B. onitoides , the latter being fairly vestigial is this species. The axial endophallite ( Fig. 9C View Figure 9 ), in particular, with one of its ends forked and the other bearing an acuminate prolongation, has a shape reminiscent of that of other phanaeines illustrated in the literature, being particularly similar to that of Diabroctis mimas (Linnaeus, 1758) , Gromphas aeruginosa and Oruscatus davus ( Tarasov & Génier, 2015; Valois et al., 2018). This similarity is likely homologous not only among these taxa, but also to the equally forked shape seen in the Eucraniini ( Tarasov & Génier, 2015), the sister-clade to the phanaeines (see more details below). Therefore, the ancestor of the Eucraniini + Phanaeini clade likely also possessed a similar forked axial endophallite, a shape that was directly inherited by Bolbites and the other mentioned taxa, and from which the more differentiated state found in groups like Phanaeus and Coprophanaeus d’Olsoufieff, 1924 derived. But readers should be cautious as to how these endophallites have been identified in the literature. As confirmed by Marcely Valois (personal communication to MC, 13 February 2020), the labelling of the axial and subaxial endophallites of Diabroctis mimas is inverted in Valois et al. ’s (2018) figure 7. The endophallite identified as the subaxial in that figure is actually the axial, whereas the one labelled as being the axial is the subaxial. The same mistake is present in the illustration of the endophallites of Eucranium arachnoides Brullé, 1838 in Tarasov & Génier’s (2015) figure 21.
General remarks: Although a precise number was not mentioned, Harold’s (1868a) original description of B. onitoides was certainly based on more than one specimen, as he characterized both sexes separately. Therefore, the type series was originally composed of an unknown number of male and female syntypes. The male designated above as the lectotype of B. onitoides was the only specimen we found that can be confidently regarded as a syntype. It is one of three individuals – two males and one female – that once belonged to the Edgar von Harold collection and are now at the MNHN via the René Oberthür collection (cf. Horn et al., 1990a; Cambefort, 2006; Cupello, 2020). Even though all three specimens agree with Harold’s (1868a) description, including the described colour (Harold’s text reads coppery, and the three individuals are indeed copper with greenish reflections), only the now-lectotype bears a label with Harold’s handwriting identifying it as ‘ B. onitoides ’. As that same label also shows in Harold’s handwriting his typical ‘t.’ abbreviation for ‘type’, there is no doubt that specimen is indeed a syntype. In turn, the other two specimens lack any labels contemporary to Harold, the oldest one borne by each of them being the typical ‘Ex-Musaeo E. Harold’ label prepared after Oberthür’s purchase of the Harold collection. While for coming from Harold’s personal collection there is a potential that these two specimens are also syntypes (now, paralectotypes), and their lack of a Harold identification label by itself does not necessary rule out that possibility (19 th- century entomologists often labelled only one specimen per series), no conclusive evidence exists confirming that they were indeed studied for the original description. For that reason, we decided to list them in the type series section above as ‘potential paralectotypes’. No other candidate syntypes were found by us in any other museum, neither did Harold state the collection where the specimens he examined came from.
As explained by Vaz-de-Mello & Grossi (2010), B. onitoides was originally described from Uruguay ( Harold, 1868a) and subsequently recorded from Argentina ( Steinheil & Strobel, 1872), Bolivia and Chile ( d’Olsoufieff, 1924), with doubts from Peru ( Balthasar, 1941), Paraguay ( Martínez, 1959) and, finally, Brazil ( Vaz-de-Mello & Grossi, 2010). d’Olsoufieff ’s (1924) citation of the species for Chile was likely based on two blue specimens deposited in the MNHN that are almost certainly mislabelled, for we have no evidence that the species occurs so high in the Andes as to be present in Chile. Balthasar’s (1941, 1951) reluctant inclusion of B. onitoides in his list of the scarabaeines from Peru, on the other hand, was based not on the examination of mislabelled specimens, but expressly on second-hand information from unspecified sources; he himself, Balthasar admitted, had never seen any evidence supporting that record. Indeed, no specimens reliably collected either in Peru or in the neighbouring Bolivian Amazon region are known to exist, only in the Bolivian Chaco. The single potential record from Peru, that of an old RBINS red female labelled ‘Pérou’, is almost certainly a result of mislabelling. It is highly unlikely, therefore, that the species is indeed present in that country, and its absence from the most recent checklists of Peruvian dung beetles ( Figueroa et al., 2014; Ratcliffe et al., 2015) is correct. Finally, its presence in Colombia, as a MNHN blue male labelled as coming from Bogotá would suggest, is even more implausible. That specimen is certainly mislabelled.
Steinheil & Strobel (1872) were the first to report the polychromatism of B. onitoides . They mentioned the examination of reddish specimens from Concordia (Entre Ríos, eastern Argentina) and Montevideo ( Uruguay), and a blue-green female from ‘Salvador’ near San Luis (western Argentina). These observations are in line with the distribution of the colour variants observed by us. On the other hand, they also mentioned the study of 15 individuals from Mendoza (eastern Argentina) described by them as being entirely black and without any metallic sheen. This is odd as none of the 1204 specimens examined by us either is black or lacks metallic reflections. Also studying Argentinian specimens, Burmeister (1874) mentioned something equally strange. He examined individuals of the entire chromatic spectrum of the species, including the first report of the bicoloured variant. Nevertheless, Burmeister referred to likely blue specimens and the elytra of the bicoloured individuals as being ‘black’ (‘ schwarze ’ in his original German). We re-examined the Bolbites of the Burmeister collection at the MACN and found only six blue specimens and no individuals of the other colour variants. The MACN general collection itself has some old, undated, bicoloured and red specimens that may have been studied by Burmeister (1874), who worked in that museum from 1862 until his death in 1892 ( Ulrich, 1972; Bragg, 2007); but none of the MACN specimens is black or has elytra of that colour. As for the specimens studied by Steinheil & Strobel (1872), we reviewed the Bolbites of the MNHN, where the Steinheil collection is currently housed ( Horn et al., 1990b), and did not find any of the specimens mentioned in that work. It is likely they were all deposited in Strobel’s collection. According to Weber (2016), Strobel’s Argentinian Coleoptera are housed in the Museo di Storia Naturale dell’Università di Parma, Italy, and some other of his South American beetles were sent to the Museo Zoologico ‘La Specola’, Florence. Unfortunately, we did not have the chance to visit any Italian museums for this work to confirm Steinheil & Strobel’s (1872) information. However, like Burmeister’s we suspect their specimens were actually dark blue instead of black as they described.
Judulien (1899) illustrated and briefly described the larva of B. onitoides , but the instar was not stated and cannot be inferred from the data provided. As we observed for the larva of Gromphas inermis Harold, 1869 (see: Cupello & Vaz-de-Mello, 2013), Judulien’s information that the B. onitoides larva bears a fivearticulated antenna is contrary to Edmonds & Halffter’s (1978) observation that the Scarabaeinae larvae have four antennomeres. Yet, under a close reading of Judulien’s work, we see that he assigned a five-articulated antenna to almost all the scarabaeine larvae he described, Sulcophanaeus menelas ( Castelnau, 1840) (as Phanaeus splendidulus ), Canthon muticus Harold, 1867 and C. edentulus Harold, 1868 being the only exceptions. Judging from his illustrations, we think it is possible that Judulien mistook the antennifer for the first antennomere, so miscounting the total number of articles in the antennae of the specimens he examined.
Ecology: Bolbites onitoides inhabits natural and artificial grasslands, savannas and dry deciduous forests from about sea level to at least 1450 m ( Kirk, 1992; Hamel-Leigue et al., 2009; Alonso et al. 2019 and label data). Label data indicate that adults are active throughout the rainiest half of the year, having been recorded from October to April. In Bolivia, in particular, Hamel-Leigue et al. (2009) recorded individuals from September to January, suggesting an earlier annual activity period there than in the examined southern part of the distribution. Also, according to Hamel-Leigue et al. (2009) and more recently Cabrera Walsh (2020), the flight activity time of B. onitoides seems to be diurnal; this can also be inferred from the metallic colouration of the species, which, at least among dung beetles, is usually associated with diurnal or crepuscular species ( Hernández, 2002; Feer & Pincebourde, 2005; Cabrera Walsh, 2020).
Concerning food habits, B. onitoides is known to be preferentially coprophagous, having been reported from cow, sheep, horse and human excrement ( Fabre, 1899, 1919; Halffter & Matthews, 1966; Kirk, 1992; Cabrera Walsh & Gandolfo, 1996; Morelli & Gonzalez-Vainer, 1997; Hamel-Leigue et al., 2009; and label data). The Belgrano ( Argentina) series studied for this work, which was collected in a trap baited with rotten chicken meat, shows that the species may also be at least occasionally necrophagous. Fabre (1899, 1919) speculated that, before the European colonization of the New World and the introduction of Old World livestock, Bolbites and the other large dung beetles of the South American grasslands relied mainly on ground sloth (Xenarthra: Folivora) dung for food. But since these giant mammals were extinct in South America for about 10 000 years before the arrival of the first European settlers ( Steadman et al., 2005), Fabre’s suggestion is, at most, only partly correct. The complete answer is probably much more trivial: between the extinction of the ground sloths and the arrival of Europeans, Scarabaeinae communities must have consumed, as they likely had done before and certainly continue to do today, dung produced by other autochthonous mammalian fauna of those habitats, including animals such as tapirs, peccaries, giant armadillos, capybaras, monkeys, deer and, for the past 15 000–11 000 years ( Dillehay, 1999; Raghavan et al., 2015), humans. Naturally, this does not exclude the possibility that, when the ground sloths were still alive, South American dung beetles, including Bolbites and its ancestors, did indeed include their excrement in their diet, as suggested by Fabre.
The nesting behaviour of B. onitoides has in part been directly observed and reviewed from the literature several times ( Fabre, 1899, 1919; Judulien, 1899; Laloy, 1902; Halffter & Matthews, 1966; Halffter & Edmonds, 1982; Cabrera Walsh & Gandolfo, 1996; Cupello & Vaz-de-Mello, 2013). In summary, B. onitoides builds pyriform brood balls typical of the Phanaeina and other dung beetles that conform to pattern II of Halffter & Edmonds’ (1982) nesting behaviour classification. Halffter & Edmonds (1982), relying on information provided by G. T. Fincher, asserted that brood balls of Bolbites apparently differ from those of the other Phanaeina by lacking a soil covering. Philips et al. (2004) repeated that information, but incorrectly mentioned Cabrera Walsh & Gandolfo (1996) as reference. In reality, all the authors who have observed Bolbites brood balls at first-hand have stated exactly the opposite: that they are entirely covered by a thick layer of soil or cement like those of the other Phanaeina (Judulien apud: Laloy, 1902; Cabrera Walsh & Gandolfo, 1996). Fabre (1899, 1919), utilizing information provided by Judulien ( Fabre, 1899: 70; 1919: 238), described in vivid prose how this external layer and the rest of the brood ball are built by B. onitoides . Cabrera Walsh & Gandolfo (1996) additionally found that all the Phanaeini species studied by them, including B. onitoides , leave a hole in the top of the cement covering of the brood ball egg chamber, an opening that presumably functions for aeration of the egg, something that had already been noted by Judulien (1899) and repeated by Fabre (1899, 1919) and Laloy (1902). The hole is subsequently covered with loose dung fibres and a thin layer of cement, but this porous, loose covering does not obstruct the air passage. This cement covering of the egg chamber and its top aeration hole are probable synapomorphies of the entire phanaeine clade (i.e. Phanaeina + Gromphadina) ( Cabrera Walsh & Gandolfo, 1996). Unfortunately, no work has so far described the roles played by each sex of B. onitoides during the construction of the nest, its provisioning with food and the preparation of the brood ball. Even so, it is reasonable to imagine that the species should follow the same pattern of the other phanaeines as discussed by Halffter & Edmonds (1982).
Finally, Martínez (1959) reported that B. onitoides , like many other dung beetles, is an intermediate host for the acanthocephalan worm Macracanthorhynchus hirudinaceus (Pallas, 1781), an intestinal parasite of mammals. Krantz (1967), in turn, asserted that mites (Acari) of the species Holocelaeno hypocrita Berlese, 1917 and H. jugulans Berlese, 1917 are phoretics of B. onitoides individuals in Argentina.
Material examined: Altogether 698 males and 506 females.
Bicoloured specimens: ARGENTINA: no further date – one female ( MFNB) and one female ( SDEI – ex Kraatz collection) . BUENOS AIRES: no further data – one male ( MNHN) ; no specific locality or date, Richter leg. – one male ( MLPA) ; Puan, Felipe Solá (‘ Est. Felipe Solá ’), II.1942, no collector – one female ( BDGC – ex A. Martínez and Henry & Anne Howden collections) ; San Isidro, Boulogne Sur Mer (‘ Boulogne’ ), II.1941, no collector – one female ( DZUP – ex F. Justus Jor collection) . CIUDAD AUTONOMA DE BUENOS AIRES: no specific locality or date, S. R. Castillo leg. – one female ( MACN) ; Núñez , 19.X.1919, no collector – one female ( MLPA) . CHACO: Chacabuco, Charata , 13.I.1993, Ugarte-Peña leg. – one male ( CEMT) ; Chacabuco, Charata, 1995, S. Bolle leg. – two males (both dissected) and two females ( BMNH) ; Chacabuco, Charata , I.1995, no collector – one male (dissected) ( NMPC – ex T. Taylor and S. Pokorný collections) ; Chacabuco, Charata , XII.2001, S. Bolle leg. – seven males and four females ( BMNH – ex J. Zídek collection) . CÓRDOBA: no further data – one male ( MACN) and one male and one female ( MLPA) ; no specific locality, XII.1896, Carlos Bruch leg. – one male ( MACN – ex Carlos Bruch collection) ; Jaime Pérez, Jaime Peter (‘ Rt. 16, 6 Km W Jaime Peter’), 09.XII.1978, Woodruff, Runnacles and Cordo leg. – one male and one female ( FSCA) ; Punilla, San Esteban, 18.XI.1976, G.J. Williner leg. – one male ( CMNC – ex François Génier collection) ; Tulumba, San José de la Dormida , II.1974, no collector – one male ( CMNC) . CORRIENTES: Ituzaingó , XII.1987, no collector – one male (dissected) ( NMPC – ex T. Taylor and S. Pokorný collections) . ENTRE RÍOS: Uruguay, Pronunciamiento , I.1962, no collector – one male and one female ( MLPA) . MISIONES: Cainguás, Dos de Mayo , II.1966, J. Foerster leg. – one male ( TAMU – ex W.D. Edmonds collection) ; Eldorado (‘ El Dorado’ ), III.1995, no collector – one male ( BDGC) and one female ( FSCA) . SALTA: no specific locality or date, Brauer leg. – three males and one female ( MFNB) ; La Viña, Ruta 68, Km 89, I.1987, no collector – one male ( CMNC) ; La Viña, Ruta 68, km 98, III.1991, no collector – one male ( CMNC) and one female ( CNCI – ex A. Martínez and Henry & Anne Howden collections) . SANTA FE: Estancia La Noria, San Javier River , 18.XII.1911, G. E. Bryant leg. – one female ( BMNH) ; Estancia La Noria, San Javier River , 09.I.1912, G. E. Bryant leg. – one female ( BMNH) ; Arroyo Colastine , ‘ 53 km S Santa Fe’, 19–21.XI.1980, R. Woodruff and H. Cordo leg. – two males and two females ( FSCA) ; General Obligado, Villa Ana , X.1924, K.J. Hayward leg. – one male ( BMNH) ; Rosario, no date, Laske leg. – one female ( SMF) . SANTIAGO DEL ESTERO: no further data – one male ( MFNB) ; no specific locality or date, Wagner leg. – one male and two females ( MLPA) ; no specific locality, IV.1962, no collector – two males ( MLPA) ; Alberdi, Campo Gallo , III.1943, no collector – one male ( DZUP – ex F. Justus Jor collection) ; Alberdi, Campo Gallo , III.1943, Prosen leg. – one male ( CMNC) , one female ( DZUP – ex CMNC) and one male and one female ( ZSM) ; Alberdi, Campo Gallo , II.1951, Prosen leg. – one male ( ZSM) ; Choya, Frías , I.2006, E. Abadie leg. – one male ( CEMT) ; Salado River , no date, Wagner leg. – one male and two females ( MLPA) . TUCUMÁN: no further data – one male ( NHMB – ex H. Marchand collection) . PARAGUAY: Chaco , XI.1996, no collector – one male and one female ( CNCI) ; ‘ Trans-Chaco’, no further data – one female ( CEMT) . PRESIDENTE HAYES: Cruce de los Pioneros , 20.I.1996, C. Aguilar leg. – one male ( TAMU – ex W. D. Edmonds collection) . URUGUAY: no further data – one male ( BMNH – ex Nevinson collection) and one female ( MFNB) . ARTIGAS: Arroyo de la Tavernada (?), 18.XI.1954, C. B. Carbonell leg. – one male ( FSCA – ex E. N. Kellesvig-Waering collection) . MONTEVIDEO: no specific locality or date, Tremorelas leg. – one female ( MLPA) ; ‘ Costa Sur’ , 26.II.1908, no collector – one female ( MLPA) . Dubious data: ‘ Chacoand. Aula (or Alba)’ – one female ( MLPA) . No data: one male [labelled with the nomen in litteris ‘ Escrapus (?) / singularis / Ur. Rei ’] ( MCZC – ex F.C. Bowditch collection), one male and one female ( NHMW – ex W. Zelenka collection) and one female (labelled with the nomen in litteris ‘ Ascropus cupreus Olivier’) ( OUMNH – ex Hope – Westwood collection) .
Blue and green specimens: ARGENTINA: no further data – one male ( MCZC – ex H.C. Fall collection), three females ( MFNB), one female ( NMPC – ex SMF and E. Witte collection), one male ( PMOC), three females ( RMNH – ex H. Gerth collection), three males and two females ( SDEI – ex Kraatz collection), two males and two females ( SMTD) and three females ( SPPC); ambiguous data (Salta or Misiones?) – one male ( MNHN); further data illegible – one male ( NHMB – ex Frey collection); Gran Chaco, no further data – three males ( RBINS); Pampa Occidental, no further data – one male ( MACN – ex Hermann Burmeister collection); Salado River, no further data – three males and one female ( BMNH – ex Nevinson collection). CATAMARCA: no further data – three males ( MACN). CHACO: no further data – one male ( NHRS); no specific locality, XI.1908, Carlos Bruch leg. – one male ( MACN – ex Carlos Bruch collection); Chacabuco, Charata, II.1997, no collector – two males and two females ( PMOC). CÓRDOBA: no further data – five males ( BMNH), one male ( BMNH – ex Nevinson collection), one male and three females ( MACN), one male ( MACN – ex Hermann Burmeister collection), one male ( MLPA), four males ( MNHN), one male ( NMPC), six males and six females ( SPPC) and one male and one female ( SMTD); no specific locality or date, Davis leg. – one male ( CNCI), five males and three females ( MCZC), one male ( NMPC – ex Nickerl collection), three males and one female ( RBINS), one male ( RMNH) and one female ( SMF – ex Pipitz and E. Witte collections); no specific locality or date, H. Richter leg. – two females ( MLPA); no specific locality or date, Weyenb. leg. – one female ( RMNH); no specific locality, 1900s, C. Bruch leg. – one female ( RBINS – ex J. J. Gillet collection); no specific locality, 07.XI.1904, C. Bruch leg. – one female ( SDEI – ex Kraatz collection); no specific locality, 10.XI.1904, C. Bruch leg. – one male ( SDEI – ex Kraatz collection); Calamuchita, ‘El Sauce’, XII.1940, Manuel J. Viana leg. – one male and two females ( MACN); Calamuchita, Villa Yacanto de Calamuchita, no date, Breyer leg. – one male ( MACN); Ischilín, Copacabana, 07.III.1980 (cow dung), Woodruff leg. – two females ( FSCA); Ischilín, ‘near Copacabana’, 08.III.1980 (pitfall with pig dung), Woodruff, Cordor and Fincher leg. – three males and six females ( FSCA); Ischilín, Quilino, XII.1965, Cichero leg. – one female ( CMNC); Jaime Pérez, Jaime Peter (‘Rt. 16, 6Km W Jaime Peter’), 09.XII.1978, Woodruff, Runnacles and Cordo leg. – 22 males and 17 females ( FSCA); Punilla, Capilla del Monte, no date, Hasseus (or Hosseus?) leg. – one male and one female ( ZSM); Punilla, Valle Hermoso, I.1942, Viana leg. – one male ( MZSP – ex Ferraciolli collection); Río Cuarto, no further data – one male ( MACN); Río Seco, no date, illegible collector name – one female ( MACN); ‘Royo Cortado’, 15.XII.1939, ‘C.M.A.M.’ leg. – one male ( MLPA); San Alberto, Las Rabonas, I.1964, no collector – one male ( MLPA); San Alberto, Las Rabonas, Sierras Grandes, I.1967, no collector – four males and three females ( MLPA); Sobremonte, San Francisco del Chañar, ‘Rumi-Puca’, 15.I.1966, Cichero leg. – two males ( CMNC); Totoral, La Pampa, no further data – one female ( MNHN). CORRIENTES: General Paz, Itá Ibaté, IX.1979, no collector – one female ( FSCA); Ituzaingó, XII.1987, S. Bolle leg. – two males (both dissected) and one female ( BMNH – ex J. Zídek collection). ENTRE RÍOS: Uruguay, Pronunciamiento, XII.1988, S. Bolle – one female ( BMNH – ex J. Zídek collection). JUJUY: no further data – three males and two females ( MACN) and one female ( MLPA). LA PAMPA: Lihuel Calel, Parque Nacional Lihué Calel, 12.II.1986, no collector – one male ( CEMT). LA RIOJA: no further data – two males ( MACN); no specific locality, I.1894, Carlos Bruch leg. – one male ( MACN – ex Carlos Bruch collection); no specific locality, I.1904, C. Bruch leg. – one female ( MLPA), one female ( RBINS – ex J. J. Gillet collection) and one male ( SDEI – ex Kraatz collection); General San Martín, Ulapes, 29.XII.2007, O. Boilly leg. – two females ( OBCL); Rosario Vera Peñaloza, Chepes (‘ 6 km E Chepes’), 10.XII.1978 [goat dung], Woodruff, Runnacles and Cordo leg. – one male ( FSCA). MENDOZA: no further data – one male ( BMNH – ex Nevinson collection), six males and two females ( MFNB), one male ( MNHN), two males ( MNHN), three males and one female ( MZSP), one male ( NHMB – ex H. Rolle and Frey collections), two females ( RBINS – ex H. Rolle and Le Moult collections), one male ( RMNH), seven males and two females ( SPPC), four males and two females ( SMTD) and one male ( ZFMK – ex H. Rolle and Fuhlrott collections); no specific locality or date, illegible collector name – one male ( MFNB). MISIONES: no further data – one male ( ZSM – ex Juan Foerster collection); Capital, Posadas, III.1997, Lucio Coronel leg. – one female ( FSCA); Eldorado, 9 de Julio, no further data – one male ( MACN). SALTA: no further data – two males and two females ( MLPA), two males and one female ( NHRS), nine males and three females ( SPPC), and one male ( SMTD); no specific locality or date, Brauer leg. – two males and one female ( MFNB); no specific locality or date, Hans Mayer leg. – one female ( NMPC); no specific locality, 1905 (?), Steinbach leg. – two males and two females ( MFNB); no specific locality, I.1949, no collector – one male ( FSCA); no specific locality, 07.II.1992, no collector – one male ( CEMT); Capital, Salta, Cerro San Bernardo, 1450 m, 15.II.1982, H. Howden and A. Howden leg. – one male ( CMNC); Cerrillos, La Merced, no date, Raimoser leg. – three males and four females ( NHMW); Chicoana, Ruta 68, II.1984, no collector – one male ( CMNC); ‘El Vino’, 24.I.2008, O. Boilly leg. – one female ( OBCL); General José de San Martín, Embarcación, XI.1953, no collector – one male (dissected) and one female (dissected) ( DZUP – ex M. Alvarenga collection); General José de San Martín, Salvador Mazza (‘Pocitos’), I.1962, A. Martínez leg. – four males and two females ( MZSP); La Viña, X.1984, M. Viana leg. – five males and five females ( FSCA); La Viña, 01.III.1991, no collector – one male and one female ( CEMT); La Viña, ‘El Carmen’, III.1943, no collector – one male and one female ( CMNC); La Viña, Ruta 68, km 89, I.1987, no collector – one male ( CMNC); ‘Picharal’, no further data – one male ( NMPC). SAN JUAN: no further data – one female ( MACN); Iglesia, Angualasto, XII.1960, no collector – one female ( MLPA). SAN LUIS: no further data – two males and three females ( MACN); no specific locality or date, Carlos Bruch leg. – one male ( MACN – ex Carlos Bruch collection); no specific locality or date, H. Richter leg. – two females ( MLPA); no specific locality, 1913, H. Gerth leg. – two males and one female ( SMF); Belgrano, ‘Fundo El Molle’, 33°02’22.8”S, 66°30’47.6”W, 622 m, 07.II.2017 (trap with chicken meat), G. Arriagada leg. – two males (both dissected) and two females ( CEMT). SANTA FE: Estancia La Noria, San Javier River, 15.XII.1911, G. E. Bryant leg. – one female ( BMNH – ex G.E. Bryant collection); Estancia La Noria, San Javier River, 18.XII.1911, G.E. Bryant leg. – three males ( BMNH – ex G. E. Bryant collection); Estancia La Noria, San Javier River, 22.XII.1911, G. E. Bryant leg. – one male ( BMNH – ex G.E. Bryant collection); Estancia La Noria, San Javier River, 10.I.1912, G. E. Bryant leg. – one female ( BMNH – ex G. E. Bryant collection); Estancia La Noria, San Javier River, 07.II.1911, G. E. Bryant leg. – one male ( BMNH – ex G. E. Bryant collection); General Obligado, Villa Ana, X.1924, K. J. Hayward leg. – one male ( BMNH). SANTIAGO DEL ESTERO: no further data no further data – four males and two females ( MACN), one male and 11 females ( MFNB), one male ( MLPA), two males and three females ( MNHN), two males and one female ( NHRS) and nine males and 12 females ( SPPC); no specific locality or date, Wagner leg. – six males ( MLPA) and one female ( SMF); uncertain specific locality (‘La Palisa bei Bracho’), ‘ Chaco de Santiago’, banks of the Salado River, no date, Wagner leg. – one male and one female ( NHMW); Alberdi, Campo Gallo, no further data – one male ( MNHN); Alberdi, Campo Gallo, III.1943, Prosen leg. – one male ( ZSM); Avellaneda, Colonia Dora, II.1951, Prosen leg. – one male ( ZSM); Avellaneda, Icaño, XII.1991, no collector – one male (dissected) ( NMPC – ex. T. Taylor and S. Pokorný collections); Avallaneda, Icaño, XII.1991, S. Bolle leg. – two males (both dissected) and two females ( BMNH – ex J. Zídek collection); Avellaneda, Icaño, Salado River, no date, Wagner leg. – 28 males and 11 females ( MNHN); Avellaneda, Icaño, Salado River, La Palma del Bracho, no further data – two males ( MNHN); Avellaneda, Icaño, Mistol Paso, banks of the Salado River, III or XII.1909, E. R. Wagner leg. – one male ( NHMB – ex Frey collection); Avellaneda, southern Chaco north of Icaño (‘ Chaco Austral au N d’Icaño’), no further data – six males and two females ( MNHN); Banda, La Banda, XI.1941, Alvaredo leg. – one female ( MACN); Banda, La Banda, IV.1960, no collector – one male and one female ( TAMU – ex W. D. Edmonds collection); Barrancas, no further data – one male and two females ( MNHN); ‘ Chaco de Santiago’, no further data – three males and one female ( NHRS) and one male ( SMTD); ‘ Chaco de Santiago’, Salado River, no further data – four males and three females ( RBINS), four females ( RBINS – ex A. Janssens’ collection), seven males and six females ( RBINS – ex Le Moult collection) and 48 males (two dissected) and 39 females ( RBINS – ex Wagner and Le Moult collections); ‘ Chaco de Santiago’, Salado River, I.(no year), no further data – one female ( RBINS – ex Le Moult collection) and two males (one dissected) and five females ( RBINS – ex Wagner and Le Moult collections); ‘ Chaco de Santiago’, Salado River, II.(no year), no further data – one female ( RBINS – ex Le Moult collection) and three males and two females ( RBINS – ex Wagner and Le Moult collections); ‘ Chaco de Santiago’, Salado River, III.(no year), no further data – one female ( RBINS – ex Le Moult collection); ‘ Chaco de Santiago’, Salado River, IV.(no year), no further data – two males and one female ( RBINS – ex Wagner and Le Moult collections); ‘ Chaco de Santiago’, Salado River, IX.(no year), no further data – one male ( RBINS – ex Wagner and Le Moult collections); ‘ Chaco de Santiago’, Salado River, XII.(no year), no further data – one male ( RBINS – ex Le Moult collection) and three males (one dissected) and four females ( RBINS – ex Wagner and Le Moult collections); ‘ Chaco de Santiago’, Salado River, 1909, no collector – six males and six females ( RBINS – ex Le Moult collection); Choya, III.1962, no collector – three males and one female ( MLPA); Choya, II.1963, Andrae (?) leg. – one male ( CMNC); Choya, II.1963, Forster leg. – one male ( CMNC); Choya, III.1974, no collector – one male and one female ( TAMU); Choya, VIII.1974, no collector – one male ( FSCA); Choya, Frías, XII.1952, no collector – one female ( CMNC); Choya, Frías, I.2006, E. Abadie leg. – one female ( CEMT); Choya, Villa la Punta, no further data – one female ( MACN); Jiménez, El Charco, II.1960, Berthold leg. – two males and two females ( ZSM); Loreto, IV.1963, no collector – one female ( MLPA); Mitre, Villa Unión, no further data – one male ( MNHN); Moreno, Puna, 11.XII.1939, Biraben-Bezzi leg. – two males ( MLPA); Ojo de Água, II.1974, A. Martínez leg. – three males and four females ( BDGC); Ojo de Água, Dique de Baez, II.1974, no collector – two males ( CMNC – ex A. Martínez and Henry & Anne Howden collections) and one male (dissected) ( DZUP – ex A. Martínez and Henry & Anne Howden collections, and CMNC); Ojo de Água, Dique de Baez (‘ Represa de Baez ’), III.1974, no collector – one male ( BDGC – ex A. Martínez collection); Salado River, no date, Wagner leg. – one male ( MACN), three males and one female ( MLPA), 15 males and 15 females ( IRSN). TUCUMÁN: no further data – one male ( NHMB – ex H. Marchand collection); further data illegible – one female ( MACN); no specific locality, III.1950, no collector – one male ( MZSP); Burruyacú, III.1946, Martínez leg. – one male ( MZSP); Burruyacú, Burruyacú, III.1946, no collector – one male ( CMNC); Cerro San Javier, XII.1948, A. Martínez leg. – one male ( MZSP); ‘Tinolina’, no further data – one male ( MNHN); Trancas, Tapia, 05.I.2009, O. Boilly leg. – one male ( OBCL). BOLIVIA: SANTA CRUZ: Andrés Ibáñez, ‘between Guayabas and Basilio’, 18°01’06”S, 63°11’24”W, 457 m, 01.I.2005, Mann, Hamel and Herzog leg. – one female ( OUMNH); Cordillera, Camiri, 20°05’45”S, 63°30’10”W, 786 m, 02.I.2005, Mann and Hamel leg. – five males and three females ( OUMNH); Cordillera, San Antonio, III.1986, Malkin (?) leg. – one male ( CMNC); Cordillera, Tatarenda, ‘Chaco’, no further data – one male ( NHRS). TARIJA: Gran Chaco, El Bagual (‘between Caiza & Creveux’), 21°50’19”S, 63°24’68”W, 487 m, 04.I.2005, Mann, Hamel and Herzog leg. – 15 males and 11 females ( OUMNH); Gran Chaco, El Bagual (‘between Caiza & Creveux’), 21°50’19”S, 63°24’68”W, 487 m, 04–05.I.2005, Mann, Hamel and Herzog leg. – two males and two females; Gran Chaco, El Bagual (‘between Caiza & Creveux’), 21°50’19”S, 63°24’68”W [sic], 487 m, 06.I.2005, Mann, Hamel and Herzog leg. – one male and two females ( OUMNH); Gran Chaco, Villamontes, I.1931, Eisennhauty leg. – six males and four females ( MFNB); Gran Chaco, Yacuiba, Yaguacua (‘Sachapera-Yaguacua’), 680 m, I.2005, D. Mann leg. – one male (dissected) ( NMPC – ex S. Pokorný collection); Gran Chaco, Yacuiba, Yaguacua (‘Sachapera-Yaguacua’) [transition between Chaco and Chaco Serrano zone; Acacia scrub, open habitat; loamy soil], 21°41’09”S, 63°33’10”W, 676 m, 03.I.2005, Mann, Hamel and Herzog leg. – one male and one female ( BDGC), seven males and three females ( OUMNH) and one male ( SPPC). PARAGUAY: no further data – one male ( FSCA), one male and one female ( MNHN) and one female ( NHMB – ex Nonfried and Frey collections); Chaco, no date, S. Bolle leg. – one male (dissected) ( BMNH – ex J. Zídek collection); ‘Trans-Chaco’, no further data – one male ( CEMT); ‘Trans-Chaco’, no date, T. Taylor leg. – one male ( BMNH – ex J. Zídek collection). BOQUERÓN: Filadelfia, Transchaco (Ruta 9) km 430, Drechsel leg. – two females ( ZFMK – ex Franz collection); Fortín Guachalla (‘Guachallas, Alto Pilcomayo’), X.1946, P. Willim leg. – one male ( CMNC). PRESIDENTE HAYES: Cruce de los Pioneros, 20.I.1996, C. Aguilar leg. – two males ( TAMU – ex W. D. Edmonds collection); Cruce de los Pioneros, 05.I.2003, C. Aguilar leg. – one male ( TAMU – ex W. D. Edmonds collection); Cruce de los Pioneros, 20.XII.2003, no collector – four males and three females ( TAMU – ex W. D. Edmonds collection); Pozo Colorado,Trans-Chaco, IV.1995, J. C. D’Alessandro leg. – two males and three females ( CEMT). Ambiguous data: ‘1908’ – one female ( MACN); Gran Chaco (Bolivian province of Gran Chaco?), no further data – one male ( BMNH – ex Nevinson collection) and one male and one female ( NHRS); ‘Jacanto’, 1930, no collector – one male ( MACN); ‘J. Caño E W’ – one male and one female ( MLPA); ‘ Pampas ’, no further data – one female ( RBINS – ex E. Candèze collection); ‘ Mont.’ (Montevideo or Monte, Argentina?), no further data – one male ( NMPC – ex SMF and E. Witte collection); ‘Parana. Febr.’ (province of Buenos Aires or Entre Ríos?), no further data – one female ( MACN – ex Hermann Burmeister collection); ‘Parana. Mrz.’ (province of Buenos Aires or Entre Ríos?), no further data – one male ( MACN – ex Hermann Burmeister collection); ‘S. Corral’, XI.1907, collector illegible – one male ( MACN); ‘S. Curral’, XII.1907, illegible collector – two females ( MACN). Illegible data: one female ( MACN). Probably erroneous data: ARGENTINA: BUENOS AIRES: no further data – three males and two females ( SMTD). BRAZIL: RIO GRANDE DO SUL: Porto Alegre, no further data – two males ( MHNG). URUGUAY: MONTEVIDEO: no further data – one male ( RBINS – ex J. J. Gillet collection). Erroneous data: CHILE: no further data – one male ( BMNH) and two males ( MNHN); no specific locality, 1870, Philippi leg. – one male ( NHMW). COLOMBIA: DISTRITO CAPITAL: Bogotá, no further data – one male ( MNHN). No data: four males ( BMNH – ex Nevinson collection), seven males and six females ( MACN), one male and one female ( MACN – ex Hermann Burmeister collection), one male ( MLPA), four males and three females ( MNHN), three males and two females ( NHMW – ex W. Zelenka collection), one female ( NHRS), one female ( NMPC – ex Dohon collection), one male ( RBINS), one male ( RBINS – ex E. Candèze collection), one female ( RBINS – ex Le Moult collection) and one female ( RMNH).
Coppery specimens: ARGENTINA: no further data – one male ( SMTD – ex C. Felsche collection) ; no specific locality or date, O.W. Thomas leg. – one male and one female ( BMNH) ; no specific locality or date, Taylor leg. – one male ( BMNH) . BUENOS AIRES: no further data – one male and one female (labelled with the nomen in litteris ‘ Escrapus singularis ’) ( OUMNH – ex Hope-Westwood collection) and one female ( SMF – ex Albert Hepp collection) ; no specific locality, II.1941, no collector – one male ( FSCA) ; no specific locality, II.1942, no collector – two males and one female ( FSCA) and two males and one female ( MZSP) ; no specific locality, I.1957, Chassot leg. – four males ( MHNG) ; Bahía Blanca , no further data – two males and three females ( TAMU – ex W. D. Edmonds collection) ; Bahía Blanca , II.1941, no collector – one male (dissected) ( CEIOC – ex J. F. Zikan collection) and two males and four females ( MZSP) ; Bahía Blanca , ‘ Patagonia’, no date, E. Weiske leg. – one male ( CNCI) , one female ( NHMB – ex Frey collection), one female ( RMNH – ex Valck Lucassen collection), one male and one female ( SDEI – ex Franklin Müller collection) and one male and one female ( SMF – ex B. Schwarzer collection) ; Merlo, Pontevedra , 02.II.1926, Brida leg. – one female ( NMPC) ; Puan, Felipe Solá , XI.1940, no collector – two males ( CMNC) ; Puan, Felipe Solá , II.1942, no collector – one male and one female ( CMNC) and one male (dissected) ( DZUP – ex F. Justus Jor collection) ; Puan, Felipe Solá , II.1942, B. Pohl leg. – one male ( MZSP) ; Puan, Felipe Solá , X.1942, B. Pohl leg. – two females ( MZSP) ; Puan, Felipe Solá (‘ Estación Felipe Sola’ ), XII.1943, no collector – two males and one female ( MNHN – ex Coiffait collection) ; Puan, Felipe Solá, II.1944, no collector – one male and one female ( NHMB – ex Frey collection) ; Puan, Felipe Solá , II.1950, no collector – one female ( CMNC – ex A. Martínez and Henry & Anne Howden collections) ; Puan, Felipe Solá , XI.1959, A. Martínez leg. – one female ( MZSP) ; Puan, Felipe Solá , II.1961, A. Martínez leg. – one female ( MZSP) ; Puan, Felipe Solá , XII.1961, A. Martínez leg. – one male ( MZSP) ; San Nicolás , V.1888, F. Ohaus leg. – one male (dissected) ( SMF) . CIUDAD AUTÓNOMA DE BUENOS AIRES: no specific locality, I.1942, no collector – one female ( DZUP – ex F. Justus Jor collection) . CÓRDOBA: ‘ El Jouer’ (?), XII.1938, M. Viana leg. – one female ( MNRJ) ; Ischilín, Quilino , XII.1965, Cichero leg. – one female ( CMNC) ; Punilla, San Esteban , 18.XI.1976, G. J. Williner leg. – one male ( CMNC) . CORRIENTES: no further data – one male (labelled with the nomen in litteris ‘ Aeschropus singularis Reiche’ ) ( BMNH) . ENTRE RÍOS: Uruguay, Pronunciamiento , XII.1988, S. Bolle leg. – one male (dissected) ( BMNH – ex J. Zídek leg.) . MENDOZA: no further data – one male ( NMPC – ex H. Rolle collection) . MISIONES: Eldorado , III.1995, no collector – one male and one female ( FSCA) . SAN LUIS: Belgrano, ‘ Fundo El Molle’ , 33°02’22.8”S, 66°30’47.6”W, 622 m, 07.II.2017 (trap with chicken meat), G. Arriagada leg. – one male (dissected) ( CEMT) GoogleMaps . SANTA FE: Arroyo Colastine , ‘ 53 km S Santa Fe’, 19–21.XI.1980, R. Woodruff and H. Cordo leg. – two males and two females ( FSCA) ; Rosario , V.1888, F. Ohaus leg. – one male ( NMPC – ex SMF) ; San Cristóbal, Hersilia (‘ La Hersilia’), no further data – one female ( NHMB – ex Hubrich and Frey collection) . SANTIAGO DEL ESTERO: ‘ Chaco de Santiago del Estero’, Salado River , no further data – one male ( RBINS – ex Le Moult collection) . BRAZIL: RIO GRANDE DO SUL: Guaraí , I.1945, no collector – two females ( MGPA) and one female ( MZSP) . URUGUAY: no further data – one female ( MFNB) , one male ( MZSP) , one female ( NMPC) , one male ( OUMNH – ex Hope-Westwood collection), one female ( RBINS – ex Le Moult collection) and one male (dissected) ( RBINS – ex Le Moult collection) ; no specific locality, 1885, Lehr leg. – one female ( ZMFK – ex Fuhlrott collection) . ARTIGAS: Arroyo de la Tavernada (?), 18.XI.1954, C. B. Carbonell leg. – one female ( FSCA – ex E. N. Kellesvig-Waering collection) . MONTEVIDEO: no further data – one male (lectotype of Bolbites onitoides ) ( MNHN – ex Edgar von Harold and R. Oberthür collections) and one male ( RBINS – ex P. de Moffarts collection) ; no specific locality, 1901, no collector – one female ( BMNH – ex Nevinson collection) ; no specific locality, 25.XII.1907, no collector – one male ( RBINS – ex J. J. Gillet collection) ; no specific locality, X.1952, no collector – one female ( DZUP – ex M. Alvarenga collection) ; Cerro de Montevideo, 12.X.1932 (on dung), no collector – one male and one female ( CEMT) ; Cerro de Montevideo (‘ Cerro ’), 18.I.1935, no collector – one male ( RMNH – ex Van Doesburg collection) ; Río de la Plata, no further data – one male and one female ( BMNH – ex Fry collection) . SALTO: ‘ 5 km NW Belén’ , 24.I.1963, A. Toccheton leg. – one female ( MZSP) . SAN JOSÉ: ‘ Mr. Claras ’, Río de la Plata (‘ La Plata ’), no further data – one female ( MNHG) . No locality data: no date, Parry leg. – one female ( RBINS – ex J. La Fontaine collection). Probably erroneous data : BRAZIL: RIO GRANDE DO SUL: Porto Alegre, no further data – two females ( MHNG) . Uncertain data: ‘10’, no further data – one male ( RBINS) ; ‘ Martinez ’ (?), 13.XI.1925, Bridarolli leg. – one male ( NMPC) . Erroneous data: ARGENTINA: Patagonia (‘ Patago - / nie ’), no further data – one male ( RMNH – ex A. J. Buis and F. C. Drescher collections) . AUSTRALIA: no further data – one male and one female ( NHMW – ex W. Zelenka collection) . No data: four females (one labelled with the nomen in litteris ‘ Coelatocollis Reiche’, another with the nomen in litteris ‘ Escrapus singularis Reiche’ ) ( BMNH – ex Nevinson collection), one male and one female (potential paralectotypes of B. onitoides ) ( MNHN – ex Edgar von Harold and R. Oberthür collection), one male ( NMPC) , two females ( RBINS) , one female ( RBINS – ex A. Janssens collection), one female ( RBINS – ex E. Candèze collection) and one male and one female ( RBINS – ex Le Moult collection) .
Red specimens: ARGENTINA: no specific locality or date, O. W. Thomas leg. – one female ( BMNH) ; no specific locality or date, Taylor leg. – one male ( BMNH) . BUENOS AIRES: no further data – one male and three females ( MACN) and one female ( MNHN – ex Granger collection) ; no specific locality or date, Carlos Bruch leg. – one male ( MACN – ex Carlos Bruch collection) ; no specific locality or date, J. Bosq leg. – one male and one female ( MLPA) ; no specific locality, I.1894, no collector – one female ( MLPA) . CHACO: Chacabuco, Charata , 13.I.1993, Ugarte-Peña leg. – one male ( CEMT) ; Chacabuco, Charata, I.1995, no collection – one male (dissected) and one female ( NMPC – ex T. Taylor and S. Pokorný collections) ; Chacabuco, Charata , XII.2001, S. Bolle leg. – one male ( BMNH – ex J. Zídek collection) ; Liberdad, ‘ 15 km from Puerto Tirol’, ‘ La Chanza’ , 03.III.1939, no collector – one male ( MLPA) . CÓRDOBA: no locality or date, Davis col. – one male and two females ( MCZC) and one male ( NMPC) . CORRIENTES: no further data – one male ( MLPA) , one male ( MNHN) and one female ( ZSM) . ENTRE RÍOS: Colón, San José, X.1925, D. Chaco expedition leg. – one male ( ZSM) ; Uruguay, Pronunciamiento , I.1962, no collector – two males ( MLPA) ; Uruguay, Pronunciamiento, XII.1988, no collector – one male (dissected) and one female ( NMPC – ex T. Taylor and S. Pokorný collection) ; Uruguay, Pronunciamiento , XII.1988, S. Bolle leg. – two males (both dissected) and one female ( BMNH – ex J. Zídek leg.) . FORMOSA: Formosa , II.1949, A. Martínez leg. – one female ( MZSP) . MISIONES: Eldorado , 07.VII.1989, no collector – one male and one female ( FSCA) . SANTA FE: General Obligado, Villa Ana , X.1924, K. J. Hayward leg. – one male ( BMNH) ; General Obligado, Villa Ana , III.1926, K. J. Hayward leg. – one male ( BMNH) ; Rosario, no date, J. Hubrich leg. – one male ( SMF) . BOLIVIA: no further data – one male (labelled with the nomen in litteris ‘ AEschropus singularis Reiche’ ) ( BMNH – ex Dejean collection) and one male ( MNHN) . BRAZIL: no specific locality or date, Parry leg. – one male ( RBINS – ex J. La Fontaine collection) . MATO GROSSO DO SUL: Porto Murtinho , 21°41’51”S, 57°53’20”W, 23.I.2008, F. R. Fernandes leg. – one male and one female ( CEMT) GoogleMaps ; Porto Murtinho, road to Pão de Açúcar , 25.I.2008, F. R. Fernandes leg. – one male ( CEMT) . RIO GRANDE DO SUL: Aceguá, Fazenda Sentinela , 31°27’30”S, 54°21’18”W, 08–14.XI.2011 (flight interception trap), R. M. Moraes leg. – one male ( CEMT) GoogleMaps ; Aceguá, Fazenda Sentinela , 31°27’19.41”S, 54°21’10.32”W, 08–14.XI.2011 (pitfall baited with human and pig excrement), R. M. Moraes leg. – one female ( CEMT) GoogleMaps ; Guaraí , I.1945, no collector – three males ( MGPA) and one male ( MZSP) . PARAGUAY: no further data – one male ( MNHN) ; Chaco , 28.X.1925, F. Felippone leg. – two males ( MACN) ; ‘ Napé’ (?), II.1928, F. Schada leg. – one female ( CEMT) . ALTO PARAGUAY: no specific locality, X.1949, A. Giai (?) leg. – one male ( MACN) ; Puerto Casado , XI.1950, A. Giai (?) leg. – two males and two females ( MACN) ; Puerto Casado (‘1.7 km SE Pto. La Victoria’ ), 22°17’40”S, 57°56’15”W, 78 m, 04–09.XII.2006, B. Garcete leg. – one male and one female ( TAMU – ex W. D. Edmonds collection) GoogleMaps ; Puerto Esperanza , 02.XII.2006, no collector – one female ( TAMU – ex W. D. Edmonds collection) . CONCEPCIÓN: Puerto Max, Estáncia Postillon, Paraguay River , no further data – two males and three females ( SMTD – ex C. Felsche collection) . GUAIRÁ: Itapé , II.1926, no collector – one female ( RMNH – ex Valck Lucassen collection) ; Mbocayaty del Guairá (‘ MBUCAYATY ’), 27.IX.1946, Schade leg. – one male ( BDGC – ex A. Martínez and Henry & Anne Howden collections) ; Villarrica, no further data – two males and one female ( MNHN) and one male and two females ( NHMB – ex Frey collection) . ITAPUÁ: San Cosme y Damián (‘ Cosme’ ), X.1948, F. H. Schade leg. – one male ( CMNC) . PARAGUARÍ: no specific locality, XI.1952, no collector – one female ( FSCA – ex E. N. Kellesvig-Waering collection) . PRESIDENTE HAYAS: Cruce de los Pioneros , 05.I.2003, C. Aguilar leg. – one female ( TAMU – ex W. D. Edmonds collection) ; Cruce de los Pioneros , 20.XII.2003, C. Aguilar leg. – one female ( TAMU – ex W. D. Edmonds collection) . URUGUAY: no further data – two males and three females ( MNHN) and one male ( MZSP) ; no specific locality or date, Fruhstorfer leg. – one female ( NHMB – ex Frey collection) ; no specific locality or date, Parry leg. – one female ( RBINS – J. La Fontaine collection) . ARTIGAS: ‘ Empalme Rutas 30 y 3’, 24.XII.1954, C.S. Carbonell leg. – two males ( CNCI) . CANELONES: no further data – one male ( MLPA) ; Canelón Chico, ‘ Los Brejos’ , 19.II.1907, Hermano Mario leg. – one male ( MLPA) ; Canelones, Juanicó, no further data – one female ( MLPA) . CERRO LARGO: no specific locality nor date, J. G. Hosmann leg. – one unsexed specimen ( MACN) . COLONIA: no further data – five males and one female ( MACN) . MONTEVIDEO: no further data – two males ( BMNH – ex Nevinson collection), four males and three females ( MNHN) , one male ( MZSP) , one male (dissected) ( RBINS – ex A. Janssens collection), one female ( RBINS – ex P. de Moffarts collection), one male (dissected) and two females ( RBINS – ex J. J. Gillet collection) and two males ( SMTD – ex C. Felsche collection) ; no specific locality or date, J. G. Hosmann leg. – one female ( MACN) ; no specific locality, II.1883, no collector – one female ( RBINS – ex Le Moult collection) ; no specific locality or date, Zufferey leg. – one female ( MHNG) ; no specific locality, 25.XII.1907, no collector – one female ( RBINS – ex J. J. Gillet collection) ; no specific locality, 25.XII.1909, no collector – one female; no specific locality, X.1952, no collector – one male (dissected) ( DZUP – ex M. Alvarenga collection) ; surroundings of Montevideo, 1867, Lasseaux leg. – one male ( NHMB – ex Frey collection) ; Cerro de Montevideo , 12.X.1932 (on unspecified dung), no collector – one male and one female ( BDGC) ; Cerro de Montevideo , 12.X.1932 (on horse and cow dung), no collector – five males and five females ( MLPA) ; Cerro de Montevideo (‘ Cerro ’), 18.I.1935, no collector – three males ( RMNH – ex Van Doesburg collection) ; Playa Punta Yeguas (‘ Pt Yeguas ’), 12.XII.1932, no collector – two females ( RBINS – ex Le Moult collection) . PAYSANDÚ: no further data – one male ( NMPC – ex R. Oberthür collection) ; Paysandú, Arroyo Sacra (‘ Sacra’ ), 20.XII.1928, no collector – one male ( MLPA) . RIVERA: Vichadero, Arroyo Coronilla , 23.X.1961, N. A. Monn leg – three males ( FSCA – ex E. N. Kellesvig-Waering collection) ; Vichadero, Santa Adelaida Farm, 31°41’04.58”S, 54°37’25.31”W, 03–11.X.2011 (pitfall baited with human and pig excrement), R. M. Moraes leg. – one female ( CEMT) GoogleMaps . SALTO: ‘ 5 km NW Belén’ , 24.I.1963, A. Toccheton leg. – one female ( MZSP) . TACUAREMBÓ: Zapucay , IV.1908, no collector – one female ( MLPA) . Probably erroneous data : ARGENTINA: TUCUMÁN: no further data – one male ( MNHB – ex H. Marchand collection) . BRAZIL: RIO GRANDE DO SUL: Porto Alegre, no further data – five males ( MHNG) . Uncertain locality: ‘ Martinez ’ (?), 13.XI.1925, Bridarolli leg. – one male ( NMPC) . No locality data: ‘ III.1943 ’ – one male ( MACN) . Ambiguous data: ‘ Cerro ’, XI.1932, no collector – one female ( RBINS – ex Le Moult collection) ; ‘ Mont. ’ (Montevideo or Monte, Argentina?) – one male ( NMPC – ex SMF and E. Witte collection) ; ‘ Río Negro (Menatra)’ – one female ( MLPA) ; ‘ Yventario’ (?) – one female ( MACN) . Erroneous data : PERU: ‘ Pérou’, ‘ Popelaire’, ‘5717’, no further data – one female ( RBINS) . No data: one male ( BMNH) , one male ( BMNH – ex Nevinson collection), eight males and three females ( MACN) , one male and three females ( MLPA) , four males and two females ( MNHN) , one male and one female ( RBINS – ex E. Candèze collection), two males and one female ( RBINS – ex Le Moult collection) and one male ( SMF – ex Albert Hepp collection) .










| R |
Departamento de Geologia, Universidad de Chile |
| SRP |
Boise State University |
| SA |
Museum national d'Histoire Naturelle, Laboratiore de Paleontologie |
| PL |
Západoceské muzeum v Plzni |
| MC |
Museo de Cipolleti |
| MFNB |
Museo Friulano di Storia Naturale |
| MNHN |
Museum National d'Histoire Naturelle |
| DZUP |
Universidade Federal do Parana, Colecao de Entomologia Pe. Jesus Santiago Moure |
| MACN |
Museo Argentino de Ciencias Naturales Bernardino Rivadavia |
| NMPC |
National Museum Prague |
| T |
Tavera, Department of Geology and Geophysics |
| FSCA |
Florida State Collection of Arthropods, The Museum of Entomology |
| TAMU |
Texas A&M University |
| CNCI |
Canadian National Collection Insects |
| SMF |
Forschungsinstitut und Natur-Museum Senckenberg |
| ZSM |
Bavarian State Collection of Zoology |
| NHMB |
Natural History Museum Bucharest |
| NHMW |
Naturhistorisches Museum, Wien |
| RMNH |
National Museum of Natural History, Naturalis |
| RBINS |
Royal Belgian Institute of Natural Sciences |
| NHRS |
Swedish Museum of Natural History, Entomology Collections |
| MZSP |
Sao Paulo, Museu de Zoologia da Universidade de Sao Paulo |
| ZFMK |
Zoologisches Forschungsmuseum Alexander Koenig |
| IRSN |
Institut Royal des Sciences Naturelles de Belgique |
| MHNG |
Museum d'Histoire Naturelle |
| MNRJ |
Museu Nacional/Universidade Federal de Rio de Janeiro |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Bolbites onitoides
| Cupello, Mario, Ribeiro-Costa, Cibele S. & Vaz-De-Mello, Fernando Z. 2022 |
Bolbites onithoides
| Di Iorio O & Turienzo P 2015: 21 |
Bolbites onitoides Harold, 1868a: 81–82
| Cabrera Walsh G 2020: 761 |
| Gillett CPT & Toussaint EFA 2020: 665 |
| Alonso CBG & Zurita GA & Bellocq MI 2019: 299 |
| Cupello M & Vaz-de-Mello FZ 2015: 16 |
| Krajcik M 2012: 54 |
| Vaz-de-Mello FZ & Edmonds WD & Ocampo FC & Schoolmeesters P 2011: 44 |
| Hamel-Leigue AC & Herzog SK & Mann DJ & Larsen TH & Gill BD & Edmonds WD & Spector S 2009: 49 |
| Scholtz CH 2009: 254 |
| Hamel-Leigue AC & Mann DJ & Vaz-de-Mello FZ & Herzog SK 2006: 6 |
| Krajcik M 2006: 16 |
| Ocampo FC & Hawks DC 2006: 562 |
| Philips TK & Edmonds WD & Scholtz CH 2004: 46 |
| Morelli E & Gonzalez-Vainer P & Baz A 2002: 54 |
| Morelli E & Gonzalez-Vainer P 1997: 197 |
| Cabrera Walsh G & Gandolfo D 1996: 582 |
| Monteresino E & Martinez A & Zunino M 1996: 109 |
| Luzzatto M & Monteresino E & Zunino M & Barbero E 1994: 63 |
| Kirk AA 1992: 53 |
| Halffter G & Edmonds WD 1982: 86 |
| Edmonds WD 1972: 818 |
| Krantz GW 1967: 12 |
| Halffter G & Matthews EG 1966: 132 |
| Martinez A 1959: 94 |
| Balthasar V 1951: 335 |
| Blackwelder RE 1944: 208 |
| Balthasar V 1941: 350 |
| Pessoa SB & Lane F 1941: 470 |
| Janssens A 1940: 1 |
| d'Olsoufieff G 1924: 20 |
| Lucas R 1920: 141 |
| Fabre J-H 1919: 242 |
| Bruch C 1911: 189 |
| Gillet JJE 1911: 80 |
| Kolbe HJ 1905: 487 |
| Laloy L 1902: 171 |
| Fabre J-H 1899: 73 |
| Judulien F 1899: 376 |
| Burmeister H 1874: 129 |
| Steinheil E & Strobel P 1872: 555 |
| Bates HW 1870: 174 |
| Harold E 1869: 1016 |
| Harold E 1868: 82 |