
Schismaderma branchi, Baptista & Pinto & Keates & Edwards & Rödel & Conradie, 2021
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5081.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:5C42BEA8-4D3E-4659-9A16-B60C19012B7B |
|
DOI |
https://doi.org/10.5281/zenodo.5776640 |
|
persistent identifier |
https://treatment.plazi.org/id/27058785-FFCC-204B-FF0A-22862DA6553D |
|
treatment provided by |
Plazi |
|
scientific name |
Schismaderma branchi |
| status |
sp. nov. |
Schismaderma branchi sp. nov.
( Figs. 6–10 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 View FIGURE 10 )
urn:lsid:zoobank.org:pub:5C42BEA8-4D3E-4659-9A16-B60C19012B7B
Holotype. ZMB 91630, adult male, road between Mussolo and Dumba Cambango, 10º06’40’’ S, 17º19’58’’ E, elevation ~ 1100 m a.s.l., Malanje Province, Angola, small depression near road, 26September 2016, coll. Pedro Vaz Pinto. GoogleMaps
Paratypes. Female. PEM A14246, a few kilometres south of Mussolo on a road between Cangandala National Park and Luando Strict Reserve, 10º08’56’’ S, 17º21’00’’ E, elevation ~ 1100 m a.s.l., Malanje Province, Angola, small pond near road, 27 October 2015, coll. Pedro Vaz Pinto ( Fig. 7 View FIGURE 7 ) GoogleMaps . Males. CHL 0338, road between Mussolo and Dumba Cambango, 10º08’51’’ S, 17º20’59’’ E, elevation ~ 1100 m a.s.l., Malanje Province, Angola, small pond near road, 26 September 2016, coll. Pedro Vaz Pinto GoogleMaps ; ZMB 91628, road between Mussolo and Dumba Cambango , 10º08’51’’ S, 17º20’59’’ E, elevation ~ 1100 m a.s.l., Malanje Province, Angola, small pond near road, 26September 2016, coll. Pedro Vaz Pinto GoogleMaps ; ZMB 91629, road between Mussolo and Dumba Cambango , 10º08’51’’ S, 17º20’59’’ E, elevation ~ 1100 m a.s.l., Malanje Province, Angola, small pond near road, 26 September 2016, coll. Pedro Vaz Pinto. GoogleMaps
Additional material. Tadpoles. PEM A14307, road between Mussolo and Dumba Cambango, 10º06’08’’ S, 17º19’50’’ E, elevation ~ 1100 m a.s.l., Malanje Province, Angola, pond near the road, 17 November 2017, coll. Pedro Vaz Pinto ( Fig. 9 View FIGURE 9 ) GoogleMaps .
Etymology. This new species is named after William Roy (Bill) Branch (1946–2018) as a tribute to this inspiring and passionate scientist, in recognition of his crucial role in African herpetology. He correctly suggested from an early stage that we were dealing with an undescribed Angolan Schismaderma , and strongly encouraged the collection of additional material and the investigative work leading to the current paper. Bill tragically passed away in October 2018, but his efforts and dedication during the last few years of his career had a major impact on the renascence of Angolan herpetology. His legacy will continue to inspire future generations. The species epithet is used as an invariable noun in apposition to the generic name. We suggest Angolan Red Toad and Sapo-vermelho de Angola as English and Portuguese common names, respectively.
Diagnosis. The molecular results confirm that the new species is a member of the family Bufonidae and the genus Schismaderma . This is also shown by the presence of granular ventral skin, pupil horizontally elongated in bright light, a conspicuous tarsal fold, inner metatarsal tubercle small and not flanged, no adhesive terminal discs on fingers and toes, and no hard claws on toes on adults ( Du Preez & Carruthers 2017). The genus Schismaderma is characterized by moderate-size, presence of external tympanum and tarsal fold, absence of externally visible parotoid glands, a prominent glandular fold running from above the tympanum laterally to near the groin and demarcating dorsal and ventral surfaces, dorsal skin granular, with reddish colouration and flat warts, ventral colouration white with bold black reticulations. This combination of features distinguishes Schismaderma from all other southern African bufonid genera and all other toad species in Angola. The back is usually uniformly coloured, sometimes carrying a pair of mid-dorsal dark blotches. The venter is heavily mottled in black, the gular region in males suffused with dark grey. The soles of hand and feet are black, with white single subarticular tubercles. The tympanum is clearly discernible, very large and rounded, the horizontal diameter larger than distance between eye and nostril. The tadpoles are also characteristic of Schismaderma , with a horse-shoe shaped fold of vascularised skin on the back ( Channing et al. 2012; Viertel & Channing 2017).
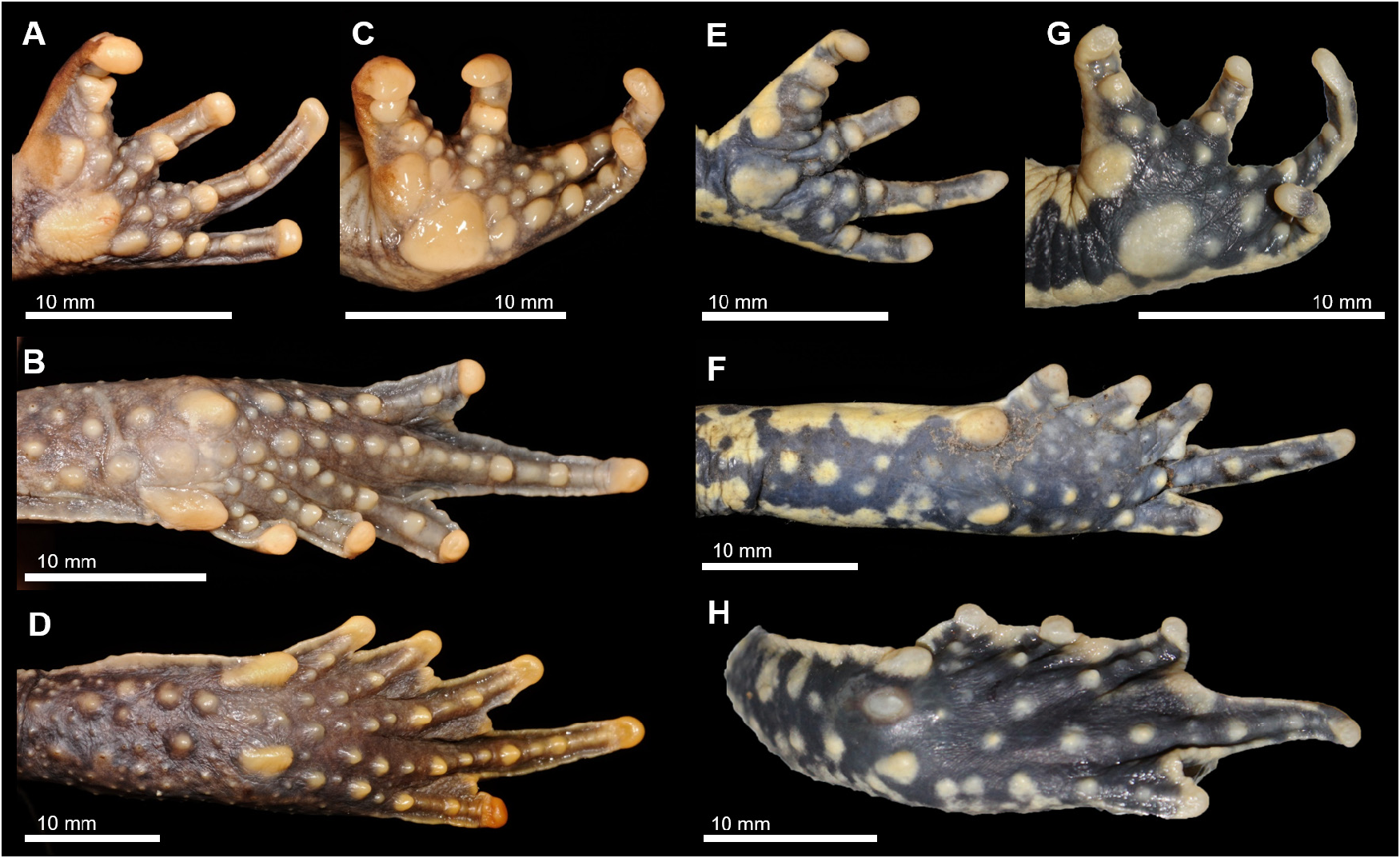
Schismaderma branchi sp. nov. can be distinguished from the only other congener, S. carens , by its smaller size; ventral skin of hind limbs, belly, chest and throat heavily mottled in black ( Figs. 6B View FIGURE 6 , 7B View FIGURE 7 ) (lightly speckled or immaculate in S. carens ); fewer and smoother tubercles on the palm of hands and sole of feet ( Fig. 8E–H View FIGURE 8 ) (prominent, well-developed, and more numerous in S. carens ); and larger inner metatarsal tubercle in relation to shortest toe. Males of S. branchi sp. nov. have shorter NL, TLB, TE, UEW, TypH and IMT, and longer EL, ED, TypW and FTL than males of S. carens .
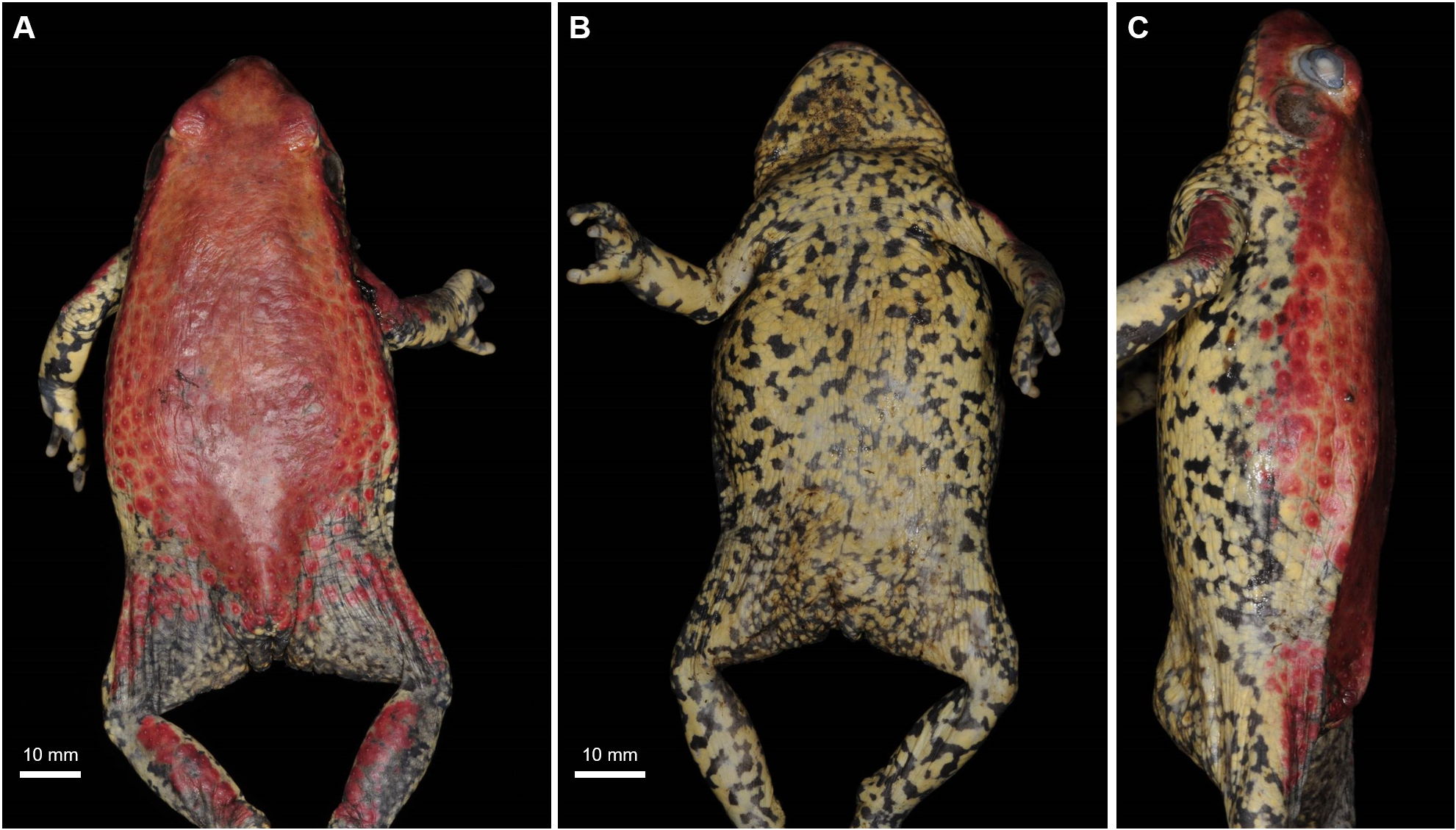
Description of the holotype. ZMB 91630 ( Fig. 6 View FIGURE 6 ) is a moderate-sized adult male (SUL 66.7 mm, with robust limbs ( Fig. 6 View FIGURE 6 ). Head triangular, longer than wide (24.9 mm x 23.4 mm); short snout (8.1 mm), projecting slightly beyond upper jaw; rostral tip slightly rounded in dorsal, ventral and lateral views. Loreal region concave, canthus rostralis rounded. Internarial region flat, small nostrils, directed dorsolaterally, much closer to the snout tip than to eyes. Tympanum and tympanic annulus clearly discernible. Tympanum diameter (6.9 mm) approximately 1.1 times eye diameter (6.1 mm). Eye diameter approximately 1.3 times eye-nostril distance (4.7 mm); eye diameter 0.6 times interorbital distance (10 mm). Interorbital distance approximately 2.8 times internarial distance (3.6 mm). Upper eyelid width 5.3 mm. Prominent eyes; not projecting laterally out of margin of head in dorsal view, projecting about 50% above margin of head in lateral view, not visible ventrally. Eye pupil ellipsoid oriented horizontally.
Well-developed limbs and digits. Thigh length 25.9 mm. Crus (tibiofibula) length 24.0 mm. Foot length 22.9 mm. First toe length 5.2 mm. Hand length 14.7 mm. Forearm length 14.9 mm. Tarsal fold present. No webbing on hands. Fingers with no terminal bulbs or discs. Nuptial pads on fingers absent. Relative length of fingers IV>III>I>II. Fingers and toes with rounded tips, and rounded, not very prominent single subarticular tubercles. A small round thenar tubercle and a large flat palmar tubercle on right hand ( Fig. 6D View FIGURE 6 ). Relative length of toes IV>III=V>II>I. Three phalanges of toe IV free of webbing; a well-developed margin of web reaching distal phalanges. Toes with small and single subarticular tubercles. Prominent oval inner metatarsal tubercle (2.8 x 1.9 mm), more than half the size of shortest toe, and around two times the size of outer metatarsal tubercle (1.7 x 1.3 mm) ( Fig. 8H View FIGURE 8 ). Both feet have an oval abrasion of the skin on the heel, between the inner and outer metatarsal tubercles ( Fig. 6B,E View FIGURE 6 ).
Dorsal granular skin with flat warts and with numerous, prominent conical asperities ( Fig. 6A View FIGURE 6 ) with a pale edge and a very small dark tip, only visible with magnifying glass. Parotoid glands absent or not visible externally. Lateral glandular ridge extending from above tympanum to near leg insertion. Skin of ventral surfaces and flanks granular ( Fig. 6A–B View FIGURE 6 ).
Colouration: After four years preserved in formalin, dorsal ground colour greyish-brown; plain dorsum with no markings; small darker blotches in fore and hind limbs. Lateral ridge paler dorsally and darker ventrally, flanks greyish-brown, darker than dorsum, mottled in black ( Fig. 6A View FIGURE 6 ). Tympanum with colour similar to dorsum, centre of tympanum with a paler blotch. Dorsal snout similar in colour and pattern to dorsum. Dorsal surface of forelimbs and fingers white with black blotches similar to ventral colouration. Lateral surface dark red speckled with black markings becoming more evident towards the belly. Dorsal surface of hind limbs greyish-brown with dark black markings. One dark blotch in middle of thigh. Base colour of dorsal foot greyish-brown; with scattered dark blotches, extending across toes. Region surrounding cloaca white heavily mottled in black. Margin of upper jaw greyish-brown with small dark markings posterior and anterior to eye. Vocal sac diffuse dark grey. Throat, belly, and ventral and anterior surfaces of fore and hind limbs white, heavily mottled in black; mottling slightly thinner in posterior part of the ventral region ( Fig. 6B View FIGURE 6 ). Soles of hands and feet black with white tubercles. Webbing of feet black with thin white edge ( Fig. 8H View FIGURE 8 ). The holotype was collected in the breeding season (as all other collected adults, see below). It is unclear if the colour might be duller during other parts of the year, as observed in S. carens (see Fig. 3 View FIGURE 3 ).
Variation of paratypes. In life, dorsal ground colour bright red. Iris golden with black pupil. Little variation was observed in the four paratypes. Three paratypes ( CHL 0338 ; ZMB 91628; ZMB 91629) are smaller than the holotype (SUL <60 mm), and one ( PEM A14246), the only female of the series, is larger (SUL = 68.7 mm). Measurements for the types are provided in Table 4 View TABLE 4 . In preserved specimens, dorsal ground colour varied from uniform dark to pale brownish, in formalin-preserved specimens ( CHL 0338 ; ZMB 91628; ZMB 91629) and bright red in a specimen preserved in ethanol ( PEM A14246). All preserved specimens have plain immaculate dorsum, except for ZMB 91628, which has a pair of mid-dorsal dark blotches. PEM A14246 had sacral markings in life, which faded after preservation in ethanol. CHL 0338 and ZMB 91629 have darker markings in dorsal surface of head, and CHL 0338 has darker markings on dorsal surfaces of fore and hind limbs slightly more conspicuous than the other specimens. Ventral colour pattern, soles of hand and feet, tubercles and feet webbing are similar among paratypes and holotype. Nuptial pads present but inconspicuous in all breeding males. The only collected female is larger than all collected males, a pattern that is expected for the species, like reported for its congeneric S. carens ( Channing & Rödel 2019) .
Tadpole morphology. Tadpoles at Gosner stage 26 ( Gosner 1960) have a morphology and behaviour typical of Schismaderma tadpoles, i.e. concerning body shape, presence of a horse-shoe shaped fold of vascularised skin on the back ( Fig. 9B View FIGURE 9 ), and congregating in compact schools ( Fig. 11F View FIGURE 11 ) ( Channing et al. 2012). Lateral view ( Fig. 9A View FIGURE 9 ): body round to ovoid. Head fold is present with the posterior edge free from the main body. Body width nearly equal to the body height. Snout oblique. Mouth directed ventrally. Spiracle sinistral, tubular, small, joined to body wall, positioned laterally and situated midbody. Dorsal view ( Fig. 9B View FIGURE 9 ): snout rounded. Nasals rounded, large (0.2 mm), positioned laterally, closer to eyes than to pupils. Eyes moderately sized (ED 8.6% of body length), positioned and directed dorsally; central parietal eye clearly visible. Ventral view ( Fig. 9C View FIGURE 9 ): internal collided intestine clearly visible through dermal skin. LTRF: 2/3. Size: Total length 13.7–17.8 mm and BL 5.3–7.8 mm. Tail 135% of the BL. Table 9 View TABLE 9 summarizes all measurements. Colouration in preservative: Body dark brown-black. Ventrum paler with scattered darker pigmentation and transparent anterior dermal skin. Tail musculature more heavily pigmented than fins.
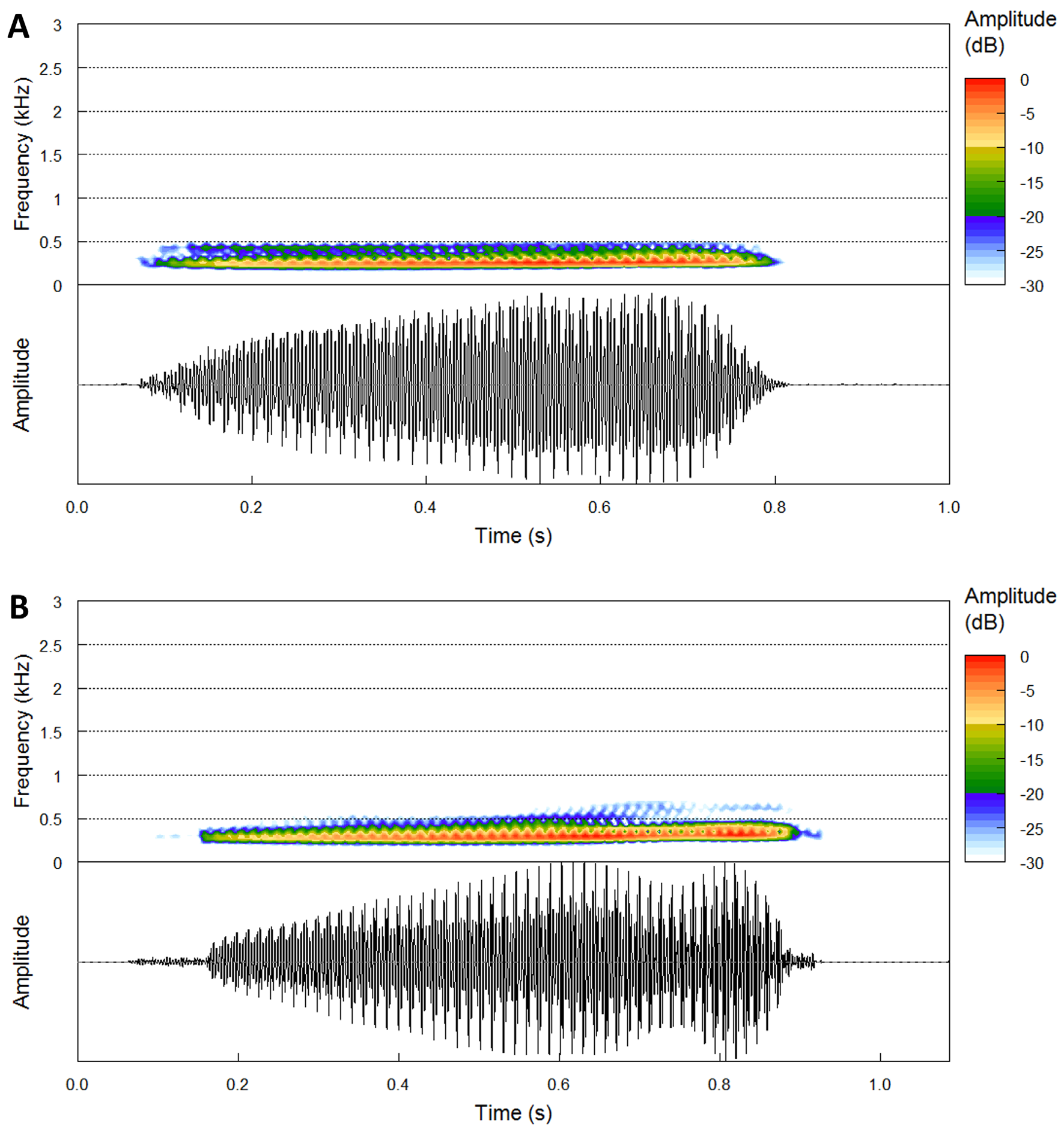
Advertisement call. The species was first found while calling in large, loud choruses from small ponds. The toads were observed calling while floating in the water, on two independent occasions in the early rainy season (see below). The call is a characteristic deep and loud “whoob, whoob.” Temporal and spectral parameters of the calls of S. branchi sp. nov. are provided in Table 10 View TABLE 10 and Fig. 10B View FIGURE 10 . Calls of S. branchi sp. nov. consist of a single note that is repeated in time, lasting 0.70 to 0.80 s (average 0.76 ± 0.04 s; n = 3), and inter-call intervals longer than the calls varying between 1.81 and 2.73 (average 2.19 ± 0.48 s). Average call repetition rate 24.57 times/minute (n = 24); average pulses per note 61–64 (average 62.33 ± 1.25; n = 3); pulse repetition rate 76.05–87.98 pulses per second (average 82.60 ± 4.94; n = 3); pulse duration 11.37–13.15 ms (average 12.15 ± 0.74 ms; n = 3). Dominant frequency of 375.0 (375.0 Hz; n = 3), mostly constant throughout the call; 90% frequency bandwidth 281.25–562.5 Hz.
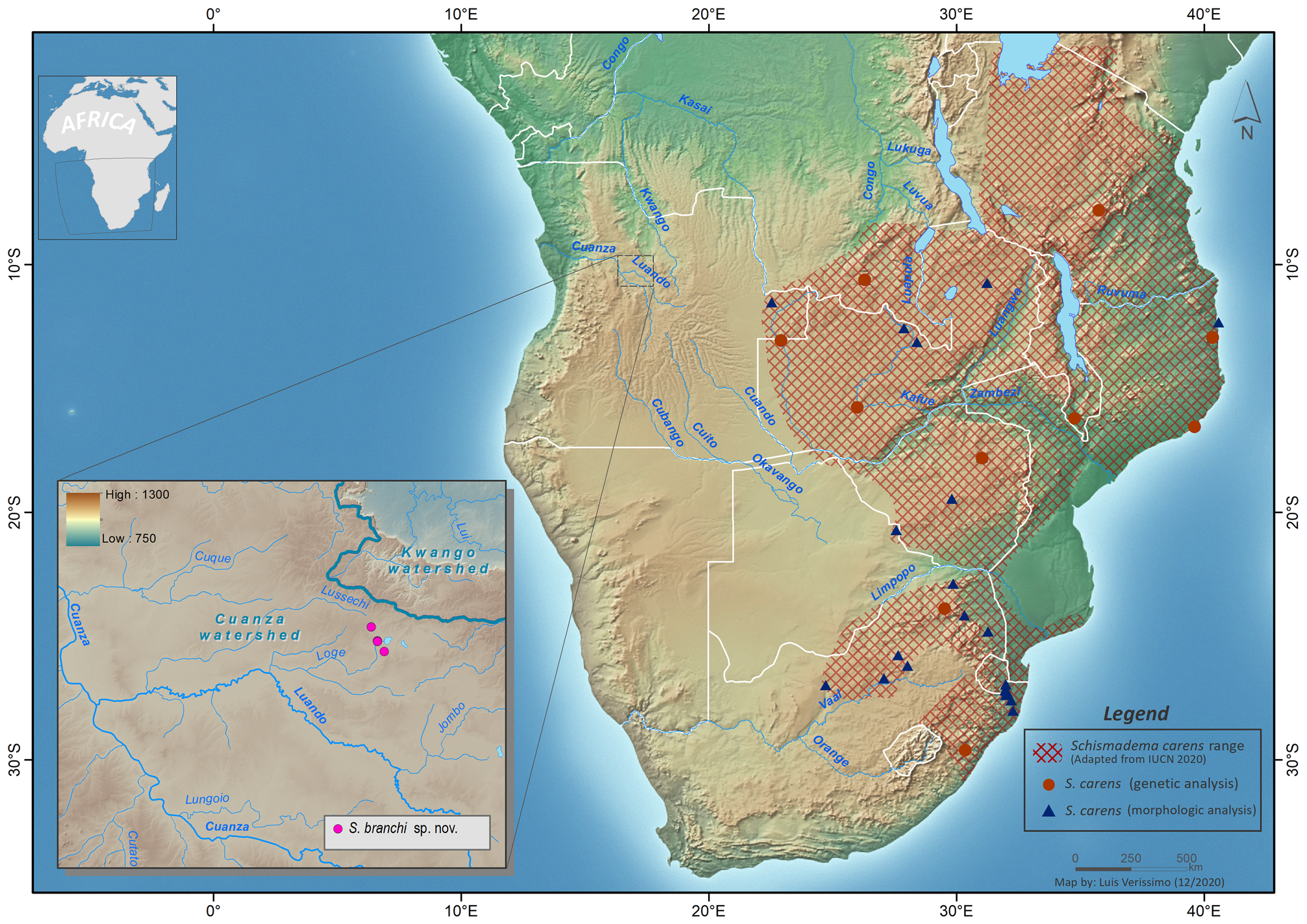
Habitat and life history notes. All specimens of S. branchi sp. nov. were found during the early rains, the species’ peak breeding season. Adults were breeding during the day in temporary pans that consisted of fairly small (2 to 5 m across) but relatively deep (1 to 2 m at the deepest section), and irregularly distributed depressions filled with murky water, recently accumulated after storms at the onset of the rainy season. These small pans were associated with small streams connected to large inland lakes within the Luando River sub-basin ( Fig. 1 View FIGURE 1 ). They had little vegetation apart from flooded grass ( Fig. 11F View FIGURE 11 ), and some were artificial, having originated from abandoned road construction works. The first time the species was found, in 27 October 2015, it was breeding in two of these temporary ponds. On a second visit to the site, in 26 September 2016, red toads were found breeding in other additional temporary ponds near the original site. Breeding was observed after heavy rains at the beginning of the rainy season in two consecutive years. On the first occasion, between 100 and 200 toads were seen in three ponds over a distance of around 5 km. On the second visit, around 50 toads were observed in two ponds. No red toads were found in the nearby streams’ main channels nor in larger perennial ponds. Observed breeding behaviour was typical of Schismaderma toads ( Balinsky 1969; Tandy & Keith 1972), with males calling loudly and incessantly throughout the day in shallow water at the edges of the pond, during a mating frenzy. Egg strings were observed floating in the water and attached to vegetation ( Fig. 11E View FIGURE 11 ). The eggs were arranged in one row; the number of eggs was not counted.Although S. carens is reported to have eggs in double strings ( Channing 2001; Du Preez & Carruthers 2017), the single strings we observed were very likely from S. branchi sp. nov., as no other amphibian species was recorded reproducing in the same ponds. Further observations are necessary to confirm if egg strings differ between the two species or not. Tadpoles moved through the pools in large dense schools, this schooling behaviour being characteristic of Schismaderma ( Fig. 11F View FIGURE 11 ). Although only found in pans while mating, S. branchi sp. nov. is expected to be terrestrial outside the breeding season like S. carens ( Balinsky 1969) . Habitat in the surrounding areas, where they likely spend most of the year, consisted of mesic woodlands and savannas within the Angolan Miombo woodlands ecoregion ( Burgess et al. 2004; Huntley 2019). Just a few kilometres to the east, there were extensive floodplains with papyrus reed beds surrounding large inland lakes. The species was found in sympatry with Afrixalus wittei , Hyperolius angolensis , H. benguellensis , H. cinnamomeoventris , Kassina senegalensis , Leptopelis bocagii , Ptychadena anchietae , Sclerophrys gutturalis , and S. pusilla .
Distribution. So far, S. branchi sp. nov. was only found in four nearby sites spanning 5 km along a road where it crosses small streams flowing to Lussechi and Loge rivers, both tributaries of the larger Luando River, part of the Cuanza River basin ( Fig. 1 View FIGURE 1 ). Just east of these sites are two large lakes, Capalanga and Quipemba surrounded by extensive swamps and floodplains and connected via the Loge River. Even though many sites have been surveyed within a 50 km radius, mostly to the north, west, and south, the species has not yet been recorded elsewhere. It is likely for S. branchi sp. nov. to have a wider distribution, which may extend into the neighbouring Cuango River drainage, but for the moment it appears to be restricted to the Luando River sub-basin, bordering the Cuanza-Congo watershed. The region where the species was found constitutes a fairly large inland depression stretching for about 100 km, roughly between the towns of Mussolo and Quirima.
Conservation status. This species is currently known from a very small area in central Angola and might be endemic to that region. It is expected to occur in a wider area in the surroundings of the type locality, but it has not yet been recorded in two formally protected areas nearby that were proclaimed primarily for the conservation of the endemic and threatened giant sable antelope ( Hippotragus niger variani ): Cangandala National Park and Luando Strict Nature Reserve, both within 50 km radius to the northwest and southwest from the type locality, respectively ( Fig. 1 View FIGURE 1 ). Its habitat does not appear to be locally threatened and the species probably tolerates disturbance, similar to S. carens ( Poynton 1996; IUCN 2013). Following IUCN criteria, uncertainties regarding abundance and distribution recommends it to be classified as Data Deficient ( IUCN 2012).




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.