
Ochotona antiqua Argyropulo & Pidoplichko, 1939
|
publication ID |
https://doi.org/10.5252/g2010n1a3 |
|
persistent identifier |
https://treatment.plazi.org/id/234087FC-FF8D-FFB4-FEC7-283AFDFCFEFE |
|
treatment provided by |
Felipe |
|
scientific name |
Ochotona antiqua Argyropulo & Pidoplichko, 1939 |
| status |
|
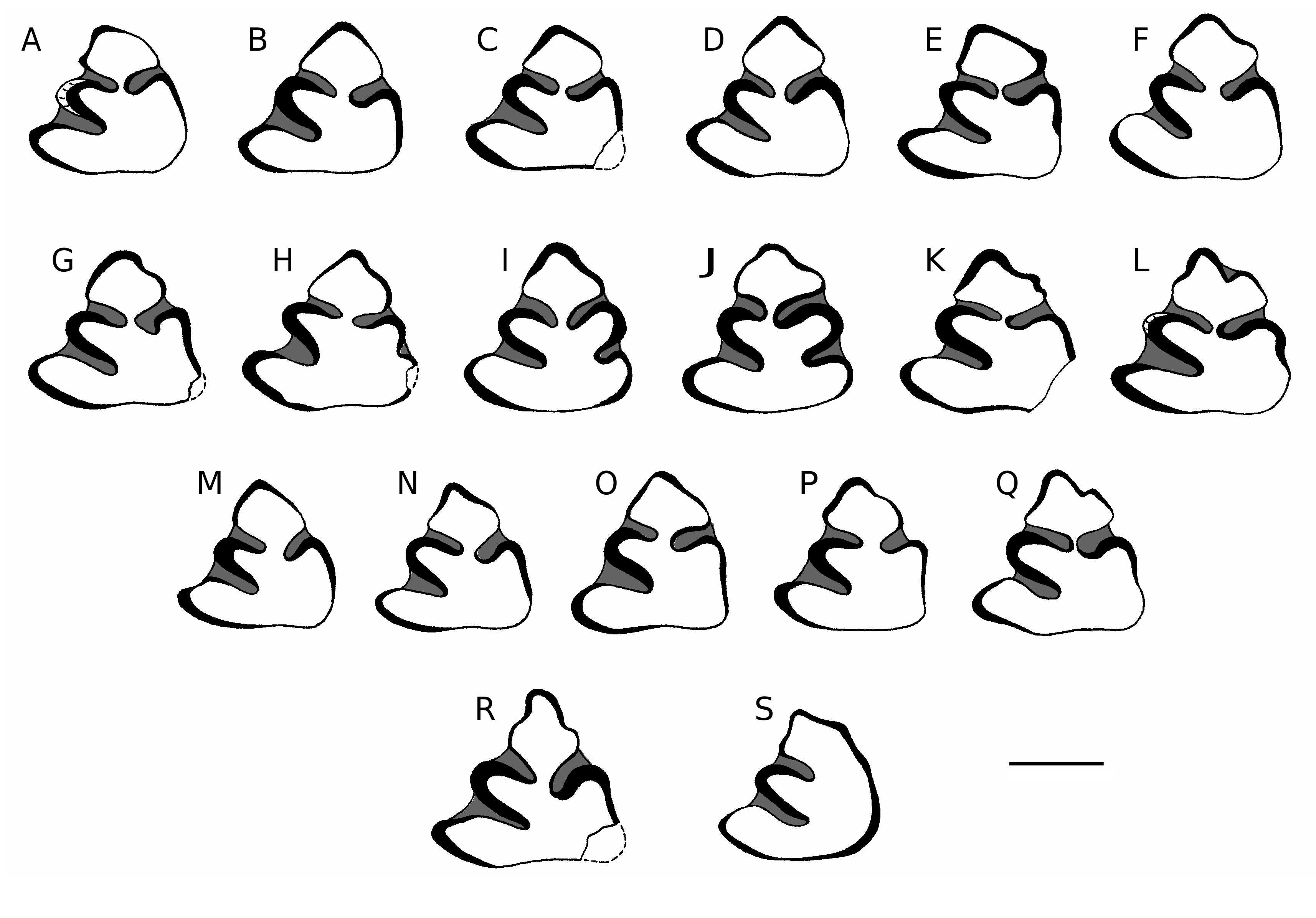
Ochotona antiqua Argyropulo & Pidoplichko, 1939 ( Figs 2 View FIG A-Q; 3; Tables 1; 2)
Ochotona pusilla antiqua Pidoplichko, 1938: 124 [ nomen nudum].
LECTOTYPE. — NNHM, no. 2616 (37-239), fragment of left mandibular ramus with p3-m2 ( Fig. 2I View FIG ); designated by Erbajeva (1988), see “Nomenclatural remarks” for details.
TYPE LOCALITY AND AGE. — Novopetrovka ( Odessa province, Ukraine); Early Pliocene (Ruscinian, MN 14) ( Fig. 1).
MATERIAL EXAMINED. — In addition to the lectotype (no. 2616) and paralectotype (no. 2618), the following specimens of O. antiqua from Novopetrovka ( NOP, N = 70) and Frunzovka-1 ( FRA, N = 18) were examined: 35 mandibles in various conditions with various teeth, or without them ( NOP21 , 22 ; NOP27-59 ), 11 maxilla fragments with various teeth, or without them ( NOP60 - NOP70 ), 42 isolated p3 ( NOP1-20 , NOP23-26 ; FRA 1-10 , FRA 12-19 ). Th e fossil remains of ochotonids are well preserved, mostly brownish and/ or blackish coloured .
EMENDED DIAGNOSIS. — A medium sized ochotonid; alveolar length of p3-m3 is 8.40-9.41-10.8, length of p3 is 1.39-1.53-1.76. The p3 anteroconid is large and wide, generally with a simple morphology and sharp anterior margin; the anteroconid-posteroconid junction is narrow and placed symmetrically to the tooth. The mandibular ramus is rather long and slight; its ventral margin is convex. Th e coronoid process is well developed.
MEASUREMENTS. — See Tables 1 and 2.
REDESCRIPTION
Ochotona antiqua is a medium sized ochotonid ( Tables 1, 2). Th e p3 possesses a relatively large and wide anteroconid, generally with a simple morphology and a sharp top. Non-plicated para- and protoflexids are nearly of the same depth, thus the very narrow (M of ccd is 0.06, N = 25) anteroconid-posteroconid junction is placed symmetrically to the tooth. The hypoflexid is rather short, generally less than half of the p3 width. The occlusal outline of P3 is trapezoidal, the mesial hyperloph is narrow (M = 65% of tooth width, N = 7). Th e p4-m3, as well as P4-M2, do not differ morphologically from the corresponding teeth of Ochotona species.
The mandible is relatively long, and in ventral view rather slight; it is higher below the p4 than below the m3. Th e mandibular diastema is relatively short (DR is M = 60, OR = 51-67, N = 18). The lower incisor extends posteriorly along the ventral border to the level of the area bounded by dividing lines of p4/m1 and m1/m2 alveoli and forms distinct tubercles on both the lingual and buccal sides of the horizontal ramus. The ventral margin of the mandible is convex. At the base of horizontal ramus, a relatively short groove extends anteriorly from the end of the tooth row along the close proximity of the lingual face ( Fig. 3 View FIG B-D). The posterior mental foramen is located ventrally to the area between m2 talonid and m3, or even more posteriorly. Th e ascending ramus is high, its coronoid process is well developed ( Fig. 3A View FIG , A’). Th e dorsal outline of mandibular condyle is triangular in shape, wide in anterior part and sharply narrower posteriorly.
VARIABILITY
The specimens of O. antiqua analyzed showed a great variability in some characters, without signifi cant differences between the localities under study. The anteroconid and paraflexid are the most variable
Ochotona antiqua Argyropulo & Pidoplichko, 1939 . Abbreviations:
parts of p3 pattern. The former is not plicated (i.e. it is with convex, or concave enamel walls without grooves, or even flexids) in 90.5% of the material under study (N = 42); three specimens (7%) are with non-cemented groove on the antero-lingual side (see e.g., Fig. 2Q View FIG ) and one specimen (2.5%) with cemented flexid ( Fig. 2L View FIG ). The ratio between symmetrical anteroconid (i.e. antero-buccal and antero-lingual enamel walls are of the similar length; Fig. 2D View FIG ) and asymmetrical one ( Fig. 2E View FIG ) is 45 vs 54% (N = 19 vs 23). Proto- and paraflexids are of the same length (55%, N = 42), nevertheless in 23% of studied p3s the former is slightly longer. The angle of paraflexid deflection from the longitudinal axis of p3 falls in a range between 43 and 74° with a mean of 59° (N = 42, CV = 14.61). In four specimens (9.5%; see e.g., Fig. 2C, E View FIG ), there is a slight indication of the “ Pliolagomys ”-like paraflexid (i.e. slightly V-shaped and bent anteriorly in its distal part). The variously developed mesoflexid occurs also in 9.5% (see e.g., Fig. 2 View FIG H-J); its occurrence seems not to be related to the age of the individual.
In side views, the outline morphology of mandible is not variable; the ventral margin of horizontal ramus is always convex ( Fig. 3A View FIG , A’). In ventral view ( Fig. 3 View FIG B-D), the buccal and lingual tubercles are developed in varying degree (viz. 57%, 30%, and 13% for morphotypes figured on Figure 3B, C View FIG , and D, respectively) and form the thickest part of the mandible. The general outlines of the p3 alveoli show the same structures ( Fig. 3 View FIG E-N) and proportions as the studied p3s; P3R is 97 (N = 28) for alveoli and 96 (N = 33) for related teeth. The posterior mental foramen is located mostly (52%, N = 32) below the m3. The root end of lower incisor extends mostly (41%, N = 32) below the trigonid/ talonid boundary of m1.
Because of the overlap of morphological characters between subadults and adults, we were able to reliably distinguish juveniles from adults only. Nevertheless, concerning the size of cheek teeth, values near the lower margin of size variation (see Tables 1, 2) are regarded here as indicative of the size boundary between subadults and adults, although the influence of subadults in measurements is possible. In any case, all specimens smaller than related size ranges (see Tables 1, 2) still retain their conical structure of teeth and were not taken into account in metric analysis.
COMPARISON
The newly re-described features as well as the variation range of the species allow us to provide here a more detailed comparison with relevant ochotonid taxa. Ochotona antiqua under study differs from stratigraphically and geographically close taxa especially in its p3 morphology and/ or in its size.
Bellatonoides kalfense ( Lungu, 1981) – “ Proochotona kalfense ” sensu Lungu 1981 ; Erbajeva 1988, 1994 – from the Vallesian (MN 9 or 10, Kalfian sensu Lungu 1978) from Kalfa ( Moldavia) differs from the studied species in its distinctly larger size (L of p3 is OR = 1.65-2.15). The p3 of Bellatonoides from Kalfa possesses buccally shifted anteroconid with a more rounded top. The enamel bridge between anteroconid and posteroconid is generally wider. Its paraflexid is less developed. The unique p3 from the Early Pliocene (MN 14) locality Krasnopol ( Ukraine) described by Topachevsky et al. (1993) as Pseudobellatona relicta – nomen dubium sensu Averianov 1998 (see below for details) – differs from O. antiqua in lacking paraflexid (only a slight depression is visible). In its size (see below), the taxon falls into the variation range of O. antiqua ( Table 1). The Early Pliocene (MN 15) Ochotonoma Sen, 1998 , known from localities of southeastern Europe and the Middle East, differs from Ochotona antiqua in its more plicated p3 anteroconid, and in its smaller and more robust mandible (see Sen 1998 and Čermák 2007 for details). All the species of Pliolagomys Agadjanian & Erbajeva, 1983 , known in the Pliocene localities from an area extending from Moldavia to Prebaikal ( Topachevsky & Skorik 1977; Agadjanian & Erbajeva 1983; Erbajeva 1988, 1994; Erbajeva & Shushpanov 1988), differ from O. antiqua in their larger size (L × W of p3 is OR = 1.35-2.60 × 1.60-3.00, N = 89 – the range is based mostly on published data, so the influence of juveniles is possible; alveolar length p3-m3: OR = 9.50-11.90, N = 12), more plicated anteroconid, and buccally moved anteroconid-posteroconid junction. The species under study differs from the former in its posteriorly bent paraflexid; only a slight indication of the “ Pliolagomys ”-like paraflexid was observed in 9.5% (N = 42) of the studied O. antiqua p3. Some p3s of young individuals of Pliolagomys may resemble O. antiqua in their occlusal size and pattern (i.e. more smooth and rounded outline of anteroconid, less asymmetric position of anteroconid-posteroconid junction and anteroconid top, as well as shorter and straighter paraflexid), but in these cases their crowns have a markedly conical structure. The shape of p3 alveolus in O. antiqua is distinguishable from that of Pliolagomys in being symmetrically located top in about half of specimens ( Fig. 3 View FIG E-I); in any case, the alveoli of O. antiqua are smaller ( Table 2) than those of Pliolagomys species.
The oldest species of the genus Ochotona View in CoL in Europe – O. eximia ( Khomenko, 1914) , “ Proochotona eximia ” sensu Khomenko 1914 ; Gureev 1964; Erbajeva 1988, 1994 – was described from the late Miocene (MN 11 or 12) locality Taraklia ( Moldavia). This taxon differs from O. antiqua in its notably larger size (see Khomenko 1914; Argyropulo & Pidoplichko 1939; Gureev 1964 for details). The species under study most closely resembles the poorly known Ochotona ursui Simionescu, 1930 , described from the Early Pliocene (Ruscinian, MN 15a; Terzea 1997) locality Măluşteni ( Romania). The lectotype, assigned to this species by Rădulescu & Samson (1995), differs from O. antiqua under study in its shorter, taller, and more robust horizontal ramus of mandible; MR is 82 in the former in contrast to 66 (M = 66, OR = 62-70, N = 29) in the latter. According to the additional material (three fragmentary mandibles without p3s) from the type locality, housed in the Natural History Museum of Basel, this ochotonid also differs from O. antiqua in its slightly elongated p3 alveolus pattern; P3R of alveoli are 103 and 97, respectively. The p3 of the lectotype also possesses a derived morphology of paraflexid (i.e. a slight indication of the “ Pliolagomys ”-like bent). Nevertheless, from the only available p3, without knowledge of morphotypes proportion in a more significant sample, it is difficult to evaluate the actual meaning and/or validity of this character in this species.
The description of Ochotona pseudopusilla , conforming Article 13 of ICZN (1999), given by Gureev & Schevtschenko in Gureev (1964), is insufficient to provide a proper comparison with O. antiqua . Unfortunately, neither holotype (No. M1-1) nor other ochotonid specimen from Schevtschenko’s collection, or at least an additional relevant ochotonid material from the type locality is available in the Institute of Geological Sciences ( Gureev 1964) or National Natural History Museum of the National Academy of Sciences of Ukraine in Kiev. Based on measurements given by Gureev & Schevtschenko (in Gureev 1964) in the original description of the species, the mandible of O. pseudopusilla should be smaller and slightly more gracile than that of O. antiqua ; OR of the p3-m3 length and height at m2 is 8.00-9.00 and 5.00-5.70, respectively, in the former compared to 8.40-10.60 and 6.50-7.00 in the latter (see Gureev 1964: 231 for details). Nevertheless, our measurements (see Table 2) as well as data provided by Argyropulo & Pidoplichko (1939) and Erbajeva & Shushpanov (1988) do not fully support their conclusions. In its size variation, O. pseudopusilla closely corresponds with O. antiqua . The morphological features of mandible in O. pseudopusilla stressed by Gureev & Schevtschenko (in Gureev 1964: 231, 232) do not allow a closer comparison with the relevant taxa. In any case, in our opinion, these characters (as e.g., the position of posterior mental foramen at mandible, etc.) suggest a closer affinity to O. antiqua rather than to O. pusilla .
The studied O. antiqua differs from all Villányian-Biharian species of Ochotona known from Central-West Europe in its larger size and/ or p3 ratio (see Sych 1980; Erbajeva et al. 2001; Čermák 2004).
The other forms, supposedly related to Ochotona antiqua , are known from the Ruscinian localities Nurnus, Armenia (MN 14/15, Kuchurganian/ Moldavian sensu Melik-Adamyan et al. 1988), and Kosyakino, Stavropol Region, Russia (MN 14 sensu Averianov & Tesakov 1998). The former one was reported by Melik-Adamyan (1986) as Ochotona ex gr. antiqua , the latter one by Averianov & Tesakov (1998) as Ochotona cf. antiqua . The p3s reported by Melik-Adamyan (1986: figs 4-7) are notably smaller (L × W of p3 is M = 1.38 × 1.35, OR = 1.24-1.50 × 1.25-1.47, N = 4; inferred from the figures 4-7) than those of typical O. antiqua . Moreover, the Armenian ochotonid differs from O. antiqua under study in its distinctively more trilobate appearance of p3 anteroconid as well as in its more evolved and wider p3 paraflexid. The ochotonid from Kosyakino is more similar to the studied O. antiqua (see Tables 1, 2; Fig. 2 View FIG ) than the Armenian form. Its size (L × W of p3 are 1.50 × 1.60 and 1.75 × 1.90; height of mandibles at p3 is 5.30 and 6.50) and the morphology of p3 (see Averianov & Tesakov 1998: fig. 1a, b) fit with that of O. antiqua (see Tables 1, 2; Fig. 2 View FIG ). Nevertheless, it differs slightly from O. antiqua in having narrower and more rounded anteroconid; AR is M = 50, OR = 43-56, N = 35 for O. antiqua from Novopetrovka and Frunzovka- 1 in contrast to 44 and 39 for ochotonids from Kosyakino.
Ochotona antiqua differs from the oldest representatives of Ochotona View in CoL in Asia known from the Latest Miocene to Early Pliocene of the Mongolian Plateau (see e.g., Qiu 1987; Erbajeva 1988, 2003); O. lagreli Schlosser, 1924 differs from the species under study in its larger size (L × W of p3 is M = 1.81 × 1.80, N = 106/101) and in having notably higher mandibular body (height at m1 is 9.00), O. birgerbohlini Averianov, 1998 – commonly known as O. minor Bohlin, 1942 (= a junior homonym of O. minor Link, 1795 ) differs in its notably smaller size (L × W of p3 is M = 1.11 × 1.12, N = 12; alveolar length p3-m3 is 6.60). Compare with new species described by Erbajeva et al. (2006) from the late Miocene of China ( Shanxi Province). The following species of Ochotona View in CoL from Asia are generally correlated with the late Pliocene, MN 16 ( Erbajeva 1994; Erbajeva & Zheng 2005). Ochotona antiqua differs from O. intermedia Erbajeva, 1976 and O. sibirica Erbajeva, 1988 ( Mongolia and Transbaikalia) as well as from O. gracilis Erbajeva & Zheng, 2005 and O. lingtaica Erbajeva & Zheng, 2005 (North China) in its larger size; from O. gromovi Erbajeva, 1976 (Transbaikalia) in its smaller size. Similarly sized O. plicodenta Erbajeva & Zheng, 2005 (North China) differs from O. antiqua by having a plicated paraflexid of p3.
NOMENCLATURAL REMARKS
The name “ antiqua ” was firstly used by Pidoplichko (1938: 124). Based on the material from Andriashevka, the new form of Ochotona was referred there to a new subspecies of O. pusilla . Although many authors, such as Argyropulo & Pidoplichko (1939), Gureev (1964), Agadjanian & Erbajeva (1983), Erbajeva (1988), Erbajeva & Shushpanov (1988), or Čermák (2004), considered this work as a description, this first usage of the name fails to conform to Article 13 of ICZN (1999); therefore it must be regarded as a nomen nudum. The name was made available by Argyropulo & Pidoplichko (1939); based on the analysis of the material from Pidoplichko’s collections (mainly from explorations in 1934, see Pidoplichko [1938] for details), they described a new species – Ochotona antiqua and designated syntypes (they used the term “types”, see Article 73.2) from Novopetrovka (no. 2616) and Grebenniky (no. 2618).
Unfortunately, from the following relevant publications (i.e. Gureev 1964; Agadjanian & Erbajeva 1983; Erbajeva 1988), it is not fully clear which specimen of the type series was subsequently designated as the single name-bearing type (i.e. lectotype). Agadjanian & Erbajeva (1983: 75) followed the terminology used by Argyropulo & Pidoplichko (1939). Gureev (1964: 231) used the term “type” for the specimen from Grebenniky (mandible without p3, no. 2618), but undoubtedly within the meaning of “ holotype ” (compare his usage of the terms “type” and “type ( lectotype)” in Gureev [1964: 83 and 106], or his subsequent usage of the term “ holotype ” for the above mentioned specimen [i.e. no. 2618] in Gromov & Baranova [1981: 70], etc.). Since Gureev (1964) did not mention the remaining specimen of the type series (i.e. the act of the explicit indication that he was selecting from the type series the particular specimen to serve as the name-bearing type; Article 74.5), we cannot consider it as a valid designation of the lectotype. In this sense of Article 74.5, we accept the work of Erbajeva (1988) as valid designation of the lectotype, insomuch she also wrongly used the terms, nevertheless, we consider here the strict combination of the terms “type” and “ paratype ” as the act of the explicit indication of specimens within the type series.
Summing up the above mentioned facts, in agreement with Erbajeva (1988) and Erbajeva & Shushpanov (1988), we consider here the syntype from Novopetrovka (no. 2616) to be the lectotype of the nominal taxon Ochotona antiqua and the remaining specimen, from Grebenniky (i.e. no. 2618), to be the paralectotype. The place of origin of the lectotype (i.e. Novopetrovka) is then the type locality of Ochotona antiqua (see Article 76.2).

| MN |
Museu Nacional, Universidade Federal do Rio de Janeiro |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Ochotona antiqua Argyropulo & Pidoplichko, 1939
| Čermák, Stanislav & Rekovets, Leonid I. 2010 |
Ochotona pusilla antiqua
| PIDOPLICHKO I. G. 1938: 124 |