
Torrenticola Piersig 1896
|
publication ID |
https://doi.org/ 10.1051/acarologia/20152155 |
|
persistent identifier |
https://treatment.plazi.org/id/2319C663-2159-ED20-AFCA-610BFD13F98D |
|
treatment provided by |
Marcus |
|
scientific name |
Torrenticola Piersig 1896 |
| status |
|
Torrenticola Piersig 1896 View in CoL
Atractides: Koch 1837: 10-11 View in CoL (in part) Koch 1842: 23 (in part) Thor 1899: 29 Thor 1902: 408 Koenike 1908: 231 Koenike 1909: 78 Walter 1908: 352 Koenike 1910: 144 Viets KH 1911: 492 Halbert 1911: 16 Viets KH 1914a: 222 Viets KH 1914b: 372 Viets KH 1916: 261 Walter 1922: 105 Thor 1923: 50 Sokolow 1926: 72 Szalay 1927: 73 Marshall 1929: 317 Halík 1930: 316 Viets KH 1930: 178 Marshall 1933: 40 Szalay 1933: 201 Sokolow 1934: 310 Viets KH 1935a: 502 Viets KH 1935b: 595 Viets KH 1936: 232 Husiatinschi 1937: 87 Oudemans 1937: 1672 Viets KH 1939: 428 Enami 1940: 213 Sokolow 1940:263 Lundblad 1941: 99 Marshall 1943: 306 Halbert 1944: 72 Szalay 1947: 289 Angelier 1949: 228 Angelier 1950: 353 Baker and Wharton 1952: 294 Walter and Bader 1952: 131.
Torrenticola: Piersig 1896: 155 Piersig 1897: 259 Koenike 1898: 376 Piersig and Lohmann 1901: 137 Piersig 1902: 849 Wolcott 1905: 196 Walter 1907: 457 Maglio 1909: 289 Monti 1910: 52 Viets KH 1916: 383 Oudemans 1941: 178 Viets KH 1949: 296 Lundblad 1951: 159 Imamura 1953a: 207 Imamura 1953b: 411 View in CoL LAEska 1953: 292 Viets KH 1953: 35 Angelier 1954: 100 Mitchell 1954: 39 Habeeb 1955: 2 Viets KO 1955: 28 Lundblad 1956a: 147 Lundblad 1956b: 642 Viets KH 1956: 235
Habeeb 1957: 13 Imamura 1957: 354 Viets KO 1958: 64 Imamura 1959a: 426 Imamura 1959b: 64 Newell 1959: 1100 Crowell 1960: 36 Habeeb 1961: 1 Lundblad 1962: 291 Besch 1964: 168 Szalay 1964: 113 Imamura 1965: 238 Cook 1966: 63 Cook 1967: 61 Conroy 1968: 28 Lundblad 1968: 320 Cook 1969: 83 Lundblad 1969: 320 Lundblad 1970: 307 LAEska 1971: 458 Lundblad 1971: 307 van Rensburg 1971: 325 Viets KO 1971a: 402 Viets KO 1971b: 758 Barr 1972: 60 Lundblad 1972: 115 Prasad and Cook 1972: 8 Cook 1974: 147 Habeeb 1974: 1 Lundblad 1974: 307 Viets KO and Böttger 1974: 126 Viets KO 1977a: 533 Viets KO 1977b: 89 Conroy 1978: 117 Davids 1979: 55 Cook 1980: 45 Wainstein 1980: 144 Viets KO 1981a: 20 Viets KO 1981b: 26 Barr 1982: 155 Smith 1982: 905 Cook 1986: 49 Bader and Sepasgozarian 1987: 183 Viets 1987: 752 (in part) Bader 1988: 87 Di Sabatino and Cicolani 1990: 44 Wiles 1991: 43 Cramer 1992: 17 Di Sabatino et al. 1992: 255 Di Sabatino and Cicolani 1993: 32 Gerecke and Di Sabatino 1996: 295 Wiles 1997: 192 Cramer and Cook 2000: 51 Peši´c and Asadi 2002: 2 Di Sabatino et al. 2003: 393 Gerecke 2003: 142 Tuzovskij 2003: 405 Peši´c et al. 2004: 1 Turan and Peši´c 2004: 39 Valdecasus 2005: 13 Peši´c et al. 2006: 45 Goldschmidt 2007: 443-450 Di Sabatino et al. 2009: 25 Goldschmidt and Smit 2009: 180 Krantz and Walter 2009: 264 Di Sabatino et al 2010: 185 Erman et al. 2010: 18 Jin et al. 2010: 111 Smith et al. 2010: 493 Peši´c et al. 2011: 3 Peši´c and Smit 2011: 188 Peši´c et al. 2012a: 459 Peši´c et al. 2012b: 18 Tuzovskij 2012: 122 Peši´c et al. 2013: 23 Tuzovskij 2013: 182 Peši´c 2014: 207 Peši´c and Gerecke 2014: 368 Peši´c and Smit 2014a: 5 Peši´c and Smit 2014b: 4.
Rusetria: Thor 1897: 20 Thor 1902: 408 .
Schizatractides: Thor 1923: 50 [hypothetical subgenus; nomen nudum].
Synatractides: Thor 1923: 50 [hypothetical subgenus; nomen nudum].
Uratractides: Thor 1929: 196 [hypothetical genus; nomen nudum].
Type species: T. anomala ( Koch 1837) [original designation: Atractides anomalus Koch 1837 ]
Note: The above taxonomic history is not comprehensive and emphasizes major or often overlooked works. The reader should refer to Viets (1987) for additional information.
Generic diagnosis — Torrenticola View in CoL can be easily differentiated from other torrenticolines by having short posterio-dorsal subcapitular apodemes (long in Monatractides and testudacarines); five palpomeres (four in Neoatractides , the only torrenticolid with this condition); a gnathosoma that cannot be greatly extended ( Pseudotorrenticola have long, slender gnathosomae that can be fully retracted within the body and extended nearly the length of the body); a rostrum of variable length (but never completely reduced as in Monatractides and Stygotorrenticola ); and the presence of a medial suture (lacking in Stygotorrenticola , the only torrenticolid with this condition).
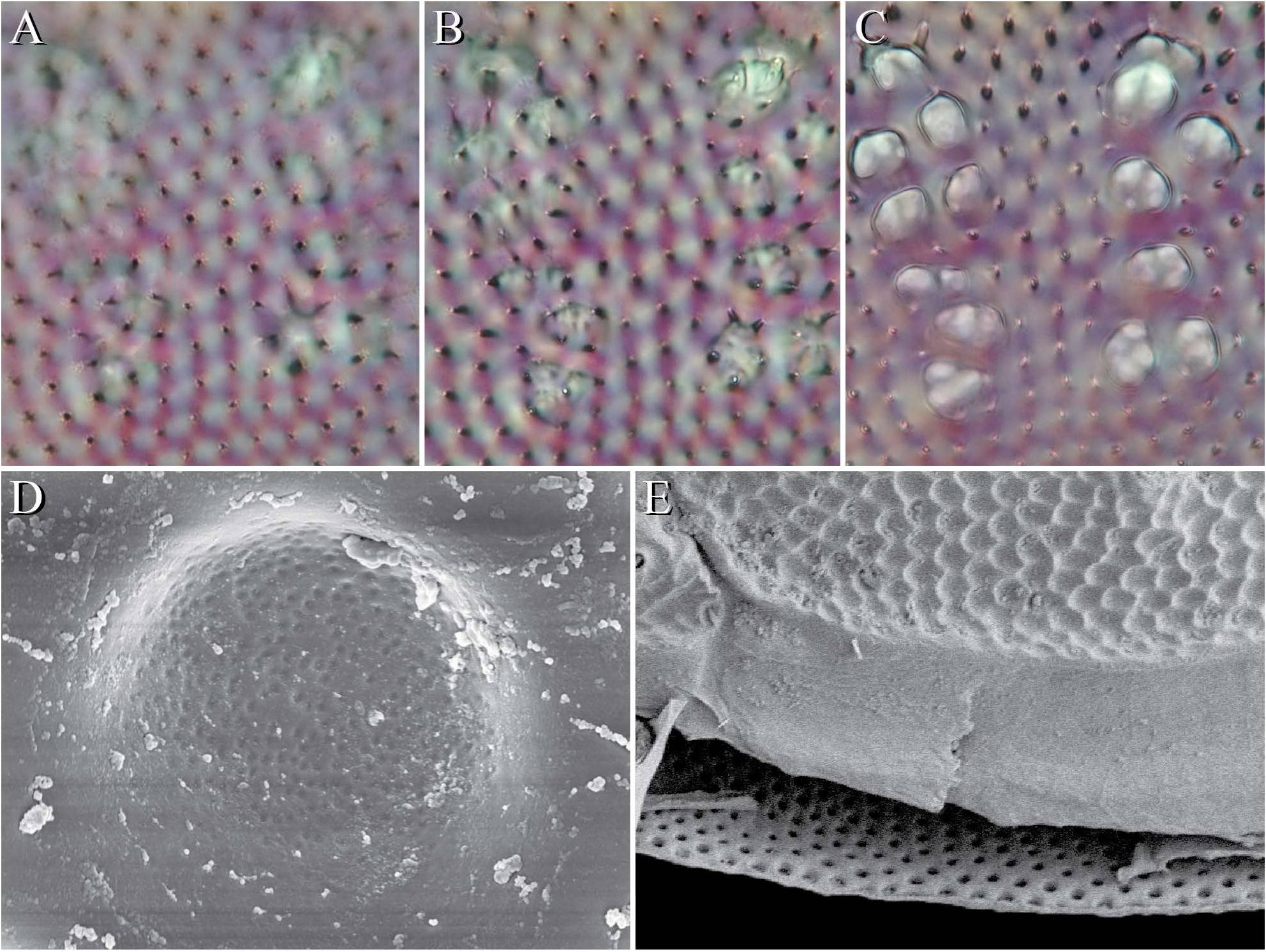
Torrenticola View in CoL can be further diagnosed by the following combination of characters. Body dorsoventrally flattened. Integument heavily sclerotized and distinctively sculptured, composed of shallow depressions; each depression representing the opening of many pits that converge within the integument to form a single channel ( Fig. 5 View FIGURE ). Fundamentally, the integument is yellowish; most species also have reddish central coloration on the dorsal plate. These colors are structural and therefore not affected by preservation technique ( Fig. 3 View FIGURE ). Upon this background, many species have developed additional coloration that is typically dark and is affected by preservation technique ( Fig. 3 View FIGURE ), suggesting these colors are not structural but pigmentation.
These color patterns can be striking and highly useful in species identification, although there is often considerable variation. Pigmented color patterns fade over time and are usually destroyed when specimens are mounted in certain media (e.g., Hoyer’s). Further coloration is achieved by internal structures; for example, the white Y-shape of the waste-filled hindgut ( Fig 2 View FIGURE E-F).
Gnathosoma capable of being withdrawn somewhat into anterior portion of idiosoma, but not attached to extensible tube. Subcapitulum with pronounced rostrum and short postero-dorsal apodemes. Oral opening generally occurs midrostrum beneath the chelicerae. Chelicerae elongate, fitting within a dorsally closed grove in subcapitulum, with movable digit modified into an up-turned fang. Pedipalps are five-segmented and variable. Often the femur and genu bear ventrodistal projections that are variously shaped and aid in species identification.
Idiosoma dorsoventrally flattened and separated into dorsal and ventral sclerotized regions by striated membrane with a fold (dorsal furrow) in the middle that contains six thin, posterior-lateral platelets organized in a ring around the posterodorsum, which are usually not visible in slide preparations. The idiosoma bears 5 pairs of lyrifis- suers (ly) and 17 pairs of glandularia (16 functional; one vestigial) each accompanied with a seta. The lyrifissures are obscured from view in most slide preparations as they reside either on the membrane of the dorsal furrow (ly-4 and -5), on the anteriormost platelet within the dorsal furrow (ly-3), or on the venter in areas that are not viewable in most slide preparations such as the area near the eyes (ly- 1) and the area immediately dorsal to Leg III (ly- 2). Glandularia are as follows: six pairs of dorsal glandularia (Dgl-1 adjacent to the eyes, Dgl-2 on the anterio-medial platelets, Dgl-3 on the anterio- lateral platelets, and Dgl-4, -5, and -6 on the main dorsal plate); four pairs of lateral glandularia (Lgl) on the lateral-most edge of the sclerotized portion of the venter, although Lgl-1 is usually not viewable in most slide preparations as it resides near the eyes; three pairs of ventral glandularia (Vgl-2 – Vgl- 4; Vgl-1 is vestigial and evident only as a small seta); and two pairs of coxal glandularia (Cxgl-2, -4; Cxgl- 1 and -3 are absent).
Dorsum consists of a large dorsal plate occupying most of the dorsum; two anterio-lateral platelets (fused with dorsal plate in some groups); and two
A – dorsum of mature adult depicting area of primary sclerotization (1°) and secondary sclerotization (2°), note dorsal glandularia 5-6 are within 2°; B – dorsum of teneral adult depicting only 1°; C – venter of mature adult depicting 1° and 2°, note ventral glandularia 1,2, & 4 are within 2°; D – venter of teneral adult depicting only 1°, note associated glandularia and excretory pore are not visible.
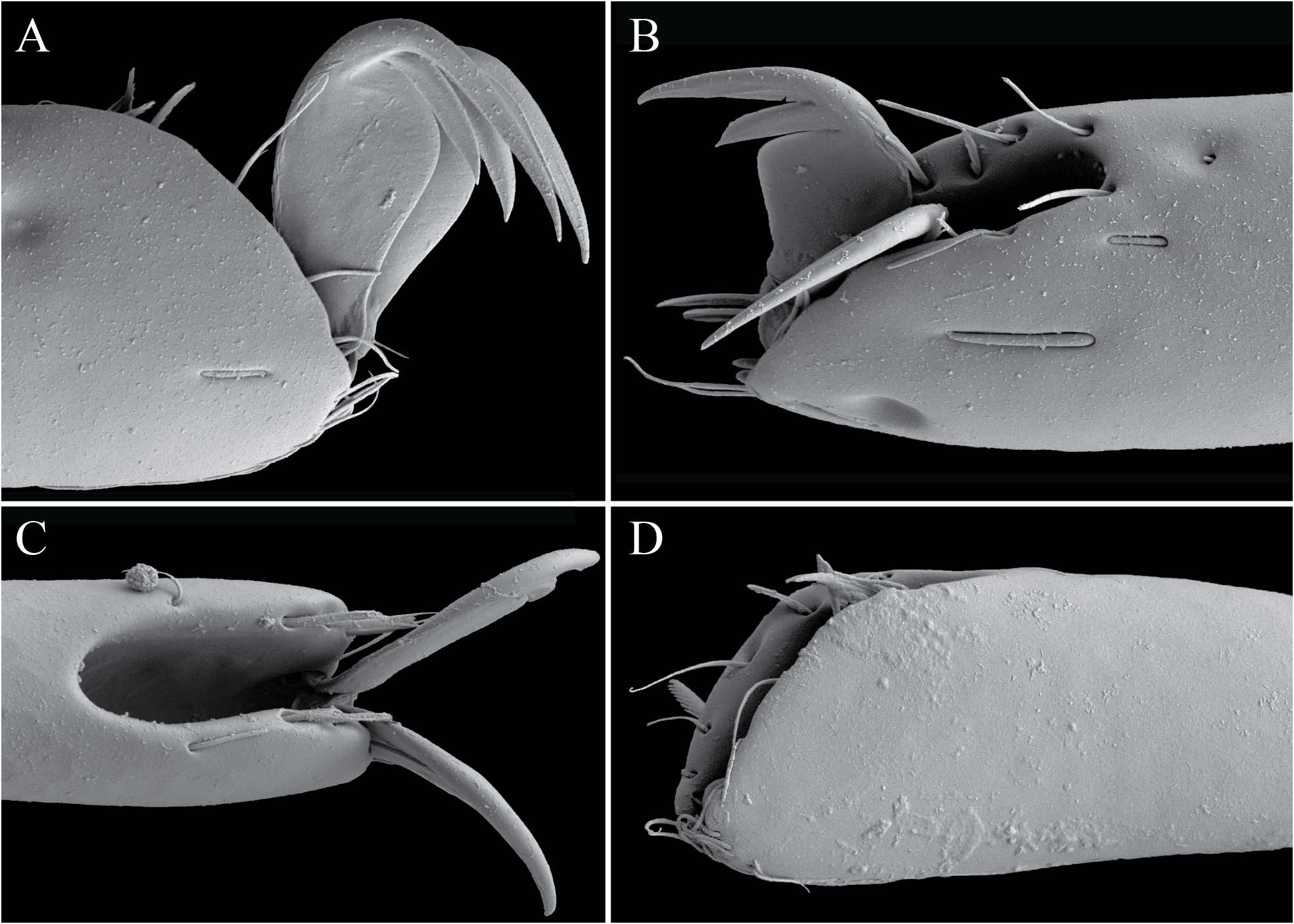
D – fronto-lateral view of claws fully retracted into tibia.
anterio-medial platelets (fused with dorsal plate in some groups). The dorsal plate is divided into area of primary and secondary sclerotization, the latter developing long after emergence from the imagochrysalis, and thus not visible when teneral but increasing in size during adult maturity ( Fig. 6 View FIGURE ). Because the excretory pore resides in the area of secondary sclerotization, teneral adults do not have an external excretory pore. The anterio-lateral platelets bear setae that are not associated with glandularia and are called postocular setae (po). The main dorsal plate centrally contains two sites of irregular circles hypothesized to be areas of muscle attachment.
Venter is completely sclerotized, but divided into an area of primary and secondary sclerotization, the later developing after emergence from the imagochrysalis. Characteristic of the family, sutures between Cx-1/2 and the suture between the medial margin of Cx-2/3 form a Y-shape. Like other lebertioids, the suture between Cx-2/3 is incomplete. Venter with five pairs of glandularia: two pairs on the coxae (Cxgl-2 and -4), three pairs of venteroglandularia (Vgl-2, -3, and -4), although one pair (Vgl-1) has been reduced to a small seta not always visible in slide preparations.
Genital field bears six pairs of acetabula and is covered by two genital flaps rimmed in numerous setae.
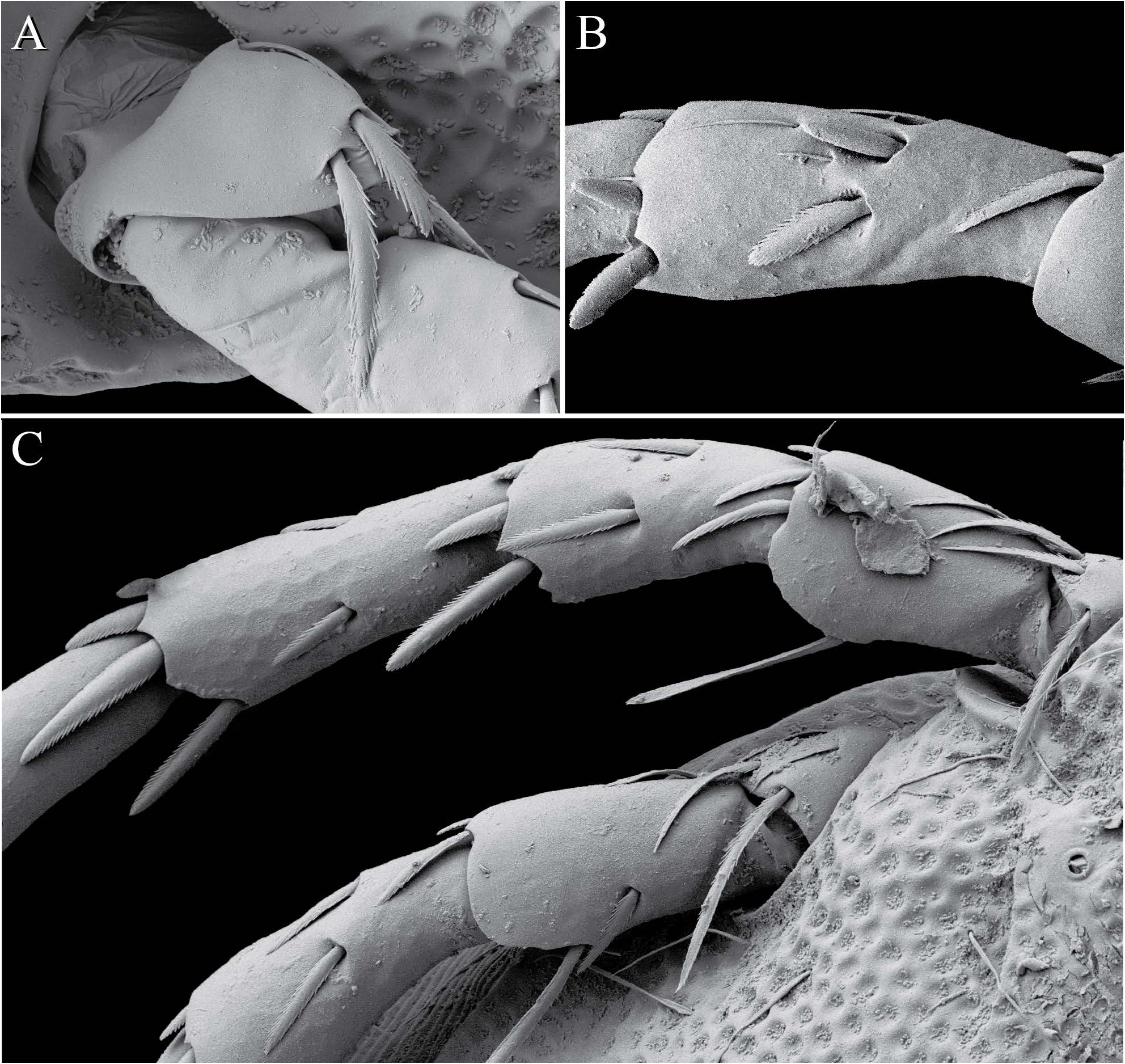
Legs lack swimming hairs and instead have large, fringed spatulate setae clearly used for digging/crawling through sediment ( Fig. 7 View FIGURE ). Legs terminate in two well-developed tarsal claws that fit into a deep tibial groove when retracted ( Fig. 8 View FIGURE B-D). Each claw is broad and shield-like basally and bifid apically ( Fig. 8 View FIGURE A-B). The first three pairs of legs are closely abutting and moved anteriorly, so that they are borne on the anterior portion of the idiosoma made by the first three coxae, and emerge dorsally. The fourth pair of legs is located ventrally, near the genital opening, and are significantly longer than the first three pairs. The insertion of the fourth pair of legs is without condyles.
Sexes are clearly differentiated by the size and shape of the genital field (smaller and rectangular in males; larger and trapezoidal in females) and the length of the medial margin of Cx-2/3 (usually significantly longer in males) referred to merely as the medial suture.
Subgeneric diagnosis. Historically, Torrenticola View in CoL has comprised multiple subgenera (e.g., Angelier 1954, Cook 1974, Bader 1988). In a seminal work that used cladistics to test torrenticolid relationships, Wiles (1997) moved most torrenticolid subgenera to other genera, thereby abolishing subgeneric classification. However, he acknowledged the suggestion by Gerecke and Di Sabatino (1996) to resurrect the subgenus Megapalpis Halbert 1944 without including members of that subgenus in his analysis. As a result, the current system consists of two subgenera: Megapalpis , identified by slender chelicerae, styletiform fangs, and a slender, curved rostrum; and Torrenticola View in CoL identified by the lack of this character combination. The species described herein is clearly not Megapalpis and therefore must be regarded by default as within the subgenus Torrenticola View in CoL . However, we refrain from recognizing subgeneric classification until robustly supported phylogenetic hypotheses corroborate such ranks.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Torrenticola Piersig 1896
| Fisher, J. R., Fisher, D. M., Nelson, W. A., O'Neill, J. C., Skvarla, M. J., Ochoa, R., Bauchan, G. R., Radwell, A. J. & Dowling, A. P. G. 2015 |
Uratractides: Thor 1929: 196
| Thor S. 1929: 196 |
Schizatractides: Thor 1923: 50
| Thor S. 1923: 50 |
Synatractides: Thor 1923: 50
| Thor S. 1923: 50 |
Torrenticola: Piersig 1896: 155 Piersig 1897: 259 Koenike 1898: 376 Piersig and Lohmann 1901: 137 Piersig 1902: 849 Wolcott 1905: 196 Walter 1907: 457 Maglio 1909: 289 Monti 1910: 52 Viets KH 1916: 383 Oudemans 1941: 178 Viets KH 1949: 296 Lundblad 1951: 159 Imamura 1953a: 207 Imamura 1953b: 411
| Lundblad O. 1956: 147 |
| Lundblad O. 1956: 642 |
| Habeeb H. 1955: 2 |
| Angelier E. 1954: 100 |
| Mitchell R. D. 1954: 39 |
| LAEska F. 1953: 292 |
| Piersig R. & Lohmann H. 1910: 155 |
Rusetria: Thor 1897: 20 Thor 1902: 408
| Thor S. 1897: 20 |
Atractides: Koch 1837: 10-11
| Baker E. W. & Wharton G. W. 1952: 294 |
| Angelier E. 1950: 353 |
| Angelier E. 1949: 228 |
| Szalay L. 1947: 289 |
| Halbert J. N. 1944: 72 |
| Marshall R. 1943: 306 |
| Lundblad O. 1941: 99 |
| Enami M. 1940: 213 |
| Sokolow I. 1940: 263 |
| Husiatinschi D. A. 1937: 87 |
| Oudemans A. C. 1937: 1672 |
| Sokolow I. 1934: 310 |
| Marshall R. 1933: 40 |
| Szalay L. 1933: 201 |
| Halik L. 1930: 316 |
| Marshall R. 1929: 317 |
| Szalay L. 1927: 73 |
| Sokolow I. 1926: 72 |
| Thor S. 1923: 50 |
| Walter C. 1922: 105 |
| Halbert J. N. 1911: 16 |
| Koenike F. 1910: 144 |
| Koenike F. 1909: 78 |
| Koenike F. 1908: 231 |
| Walter C. 1908: 352 |
| Thor S. 1899: 29 |
| Koch C. L. 1842: 23 |
| Koch C. L. 1837: 11 |