
Torrenticola trimaculata Fisher, 2015
|
publication ID |
https://doi.org/ 10.1051/acarologia/20152155 |
|
persistent identifier |
https://treatment.plazi.org/id/2319C663-2147-ED14-ACF3-6294FBDEF98D |
|
treatment provided by |
Marcus |
|
scientific name |
Torrenticola trimaculata Fisher |
| status |
sp. nov. |
Torrenticola trimaculata Fisher n. sp.
(http://species-id.net/wiki/ Torrenticola _trimacula ta)
LSID — urn:lsid:zoobank.org:act:BE021914-89AE-4DE7-A0DC-18CF80BA78AF
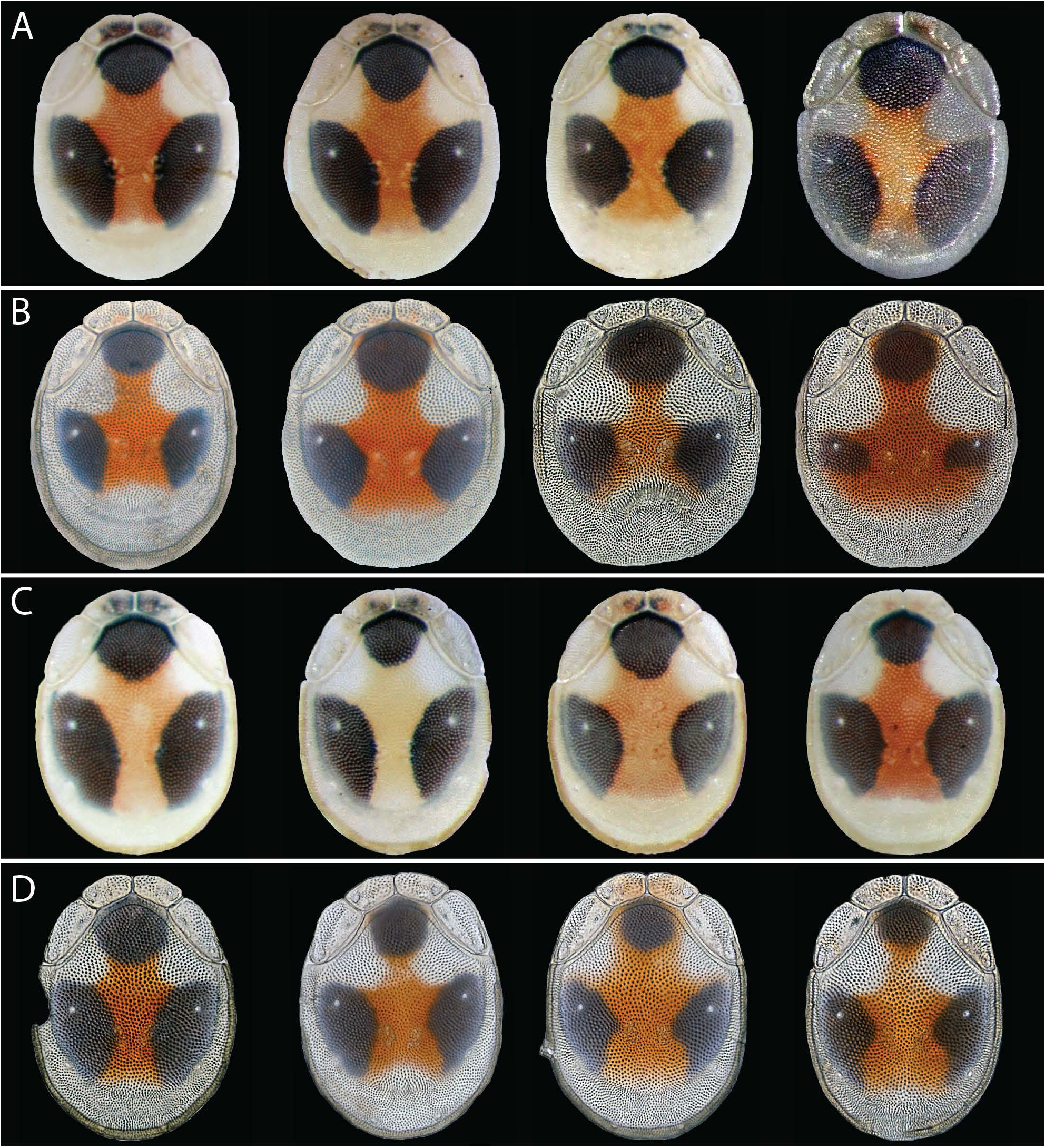
IMAGO GENERAL FEATURES. Color variable across two distinct color morphs that do not coexist within a stream ( Fig. 2 View FIGURE , 9 View FIGURE ). For both morphs, the integument is yellowish, with a central red structural coloration on the dorsal plate. Both morphs also have pigmentation in the form of three dark spots on the dorsal plate that do not extend beyond the area of primary sclerotization; this pigmentation is destroyed during clearing ( Fig. 3 View FIGURE ). This adequately describes the less pigmented morph (Morph-2: Fig. 2B, D, F View FIGURE ), although some specimens express light pigmentation on the genital plates ( Fig. 2B View FIGURE ) and hind coxae (males) ( Fig. 2D View FIGURE ), and size and shape of the dorsal spots varies considerably ( Fig. 9B, D View FIGURE ). The more pigmented morph (Morph-1: Fig. 2A, C, E View FIGURE ) is marked by the following: 1) area of primary sclerotization on the venter is darkly pigmented; 2) the three dorsal spots are larger; 3) entire gnathosoma, including pedipalps, is pigmented; 4) dorsal pigmentation can extend to the anteriomedial platelets; and 5) legs are pinkish-orange. Although noticeable variability is present in the size and shape of dorsal spots, it is much less extreme in Morph-1 ( Fig. 9A, C View FIGURE ). As with Morph-2, Morph- 1 exhibits considerable variability in size and shape of dorsal spots, as well as the extent and vibrancy of all pigmentation.
Sexes are somewhat dimorphic ( Fig. 10 View FIGURE ). Sexual dimorphism consistent with most/all Torrenticola include the following: 1) body slightly smaller in males and consistently ovoid; females larger and vary from round to ovoid; 2) genital fields comparatively small and rectangular in males; female genital plates are larger and trapezoidal; 3) medial suture of males long–comparable to the length of the genital field; female medial suture short–about as long as wide. Additional sexually dimorphic characters that are not shared by most Torrenticola include the following: 1) hind coxae not extending anteriorly beyond hind leg insertions in males but do in females; 2) hind coxae not extending far posteriorly beyond the genital field; female hind coxae extend beyond the genital field by approximately half the length of the genital field; and 3) rostrum directed normally (forward) in females, but is directed downwards in males.
FEMALE (n=49) (holotypic measurements in parentheses when available) with characters described in generic diagnosis and general features, with following specifications.
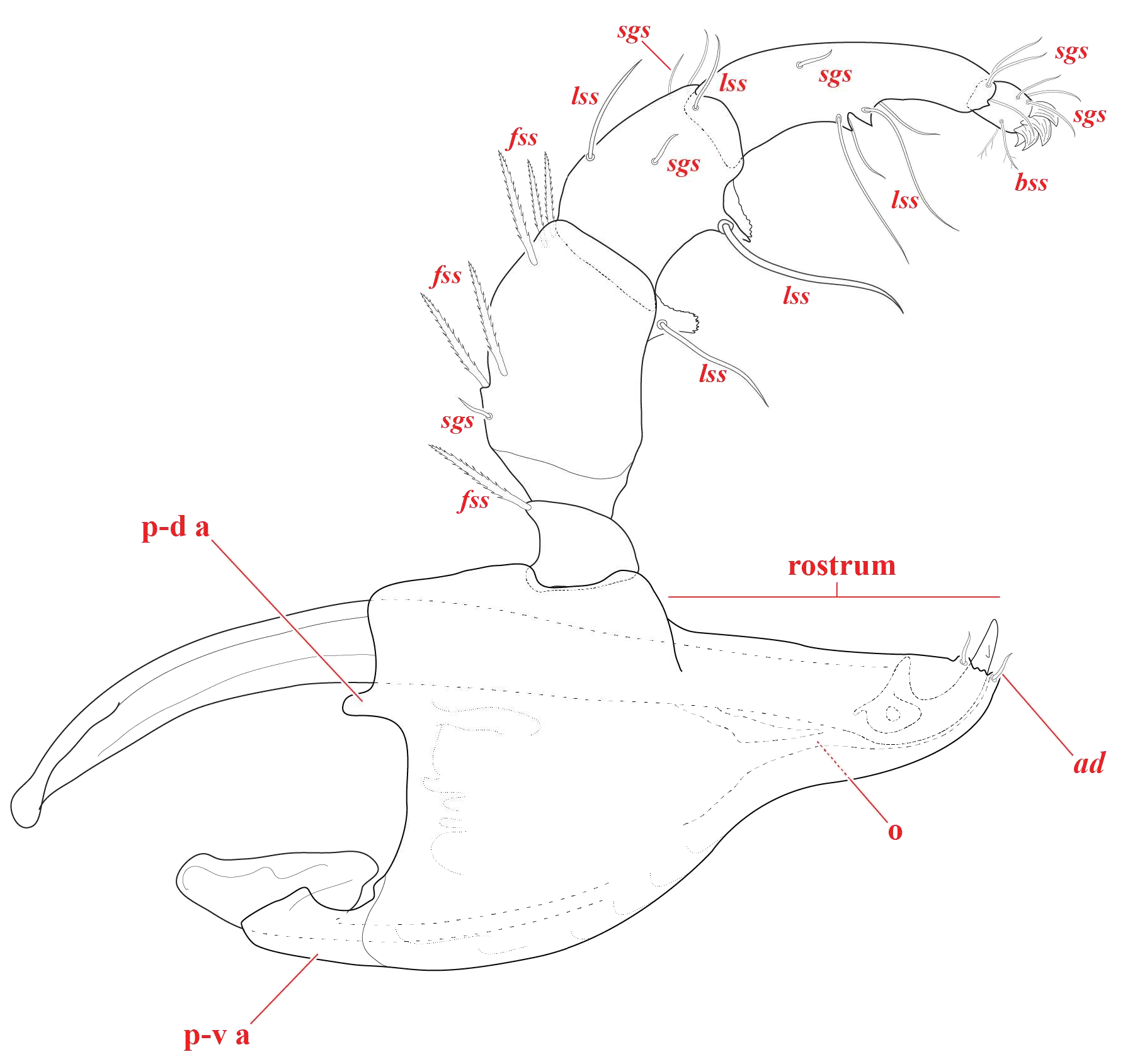
Gnathosoma ( Fig. 11-14 View FIGURE View FIGURE View FIGURE View FIGURE ) — Subcapitulum [250 – 341 (313) ventral length; 185 – 255 (239) dorsal length; 110 – 169 (142) tall] posterior edge nearly vertical, ventral bend depth slight [4 – 20 (5)], and with short rostrum [95 – 133 (129) long] that is directed forwards. Two pairs of adoral setae rim the rostral opening ( Fig. 11-12 View FIGURE View FIGURE ). Chelicerae [230 – 329 (310) long; 15 – 31 (17) high] unmodified with strongly curved fangs [33 – 74 (56) long]. Each fang with lateral and medial teeth presumably used to anchor to prey after puncturing ( Fig. 12 View FIGURE B-D). Pedipalps [248 – 338 (311) long] with dentate ventrodistal projections medially on femora and mediocentrally on genua ( Fig. 11 View FIGURE , 13 View FIGURE ). These projections vary across individuals in thickness, length, and shape. Further, their appearance can vary according to their position in a given slide preparation, sometimes even appearing tuburculate/edentate. Regardless, they are never lamellate as in the T. serratipalpis -group identified by Goldschmidt (2007). Trochanters [24 – 38 (35) long; 28 – 43 (33) wide] with one dorso-distal fringed spatulate seta (fss). Femora [80 – 123 (109) long; 48 – 68 (60) wide] with one long simple seta (lss) associated with the ventral projection and six dorsal setae as follows: proximally one short simple grooved seta (sgs); two central fss, and three distal fss (two medial; one lateral). Genua [55 – 85 (77) long; 40 – 56 (51) wide] shorter than femora with one lss associated with the ventral projection, one short sgs laterally, and four dorsal setae as follows: one central lss, and three setae distally as follows: one sgs medially, one lss medially, and one lss laterally. Tibiae [73 – 124 (107) long; 19 – 37 (34) wide] subequal in length to femora, with two short, spiny tubercles midventrally that are edentate and associated with 3-4 lss ( Fig. 11 View FIGURE , 13E View FIGURE ). Mid-dorsally, there are two sss (one proximo-lateral; one disto-medial). Distally, there is one lss doro-centrally; two lss dorso-medially; two lss dorso-laterally; one lss laterally; and one large, grooved, spine-like seta dorso-medially ( Fig. 11 View FIGURE , 14 View FIGURE A-B). Tarsi [20 – 27 (25) long; 12 – 16 (16) wide] are accompanied by four tarsal claws, with the bottom two paired ( Fig. 14B View FIGURE ), thus appearing as three claws in most slide preparations. Ventrally, there are 2-3 short bifurcating setae (sbs) and dorsally there are three lss ( Fig. 14 View FIGURE A-B).
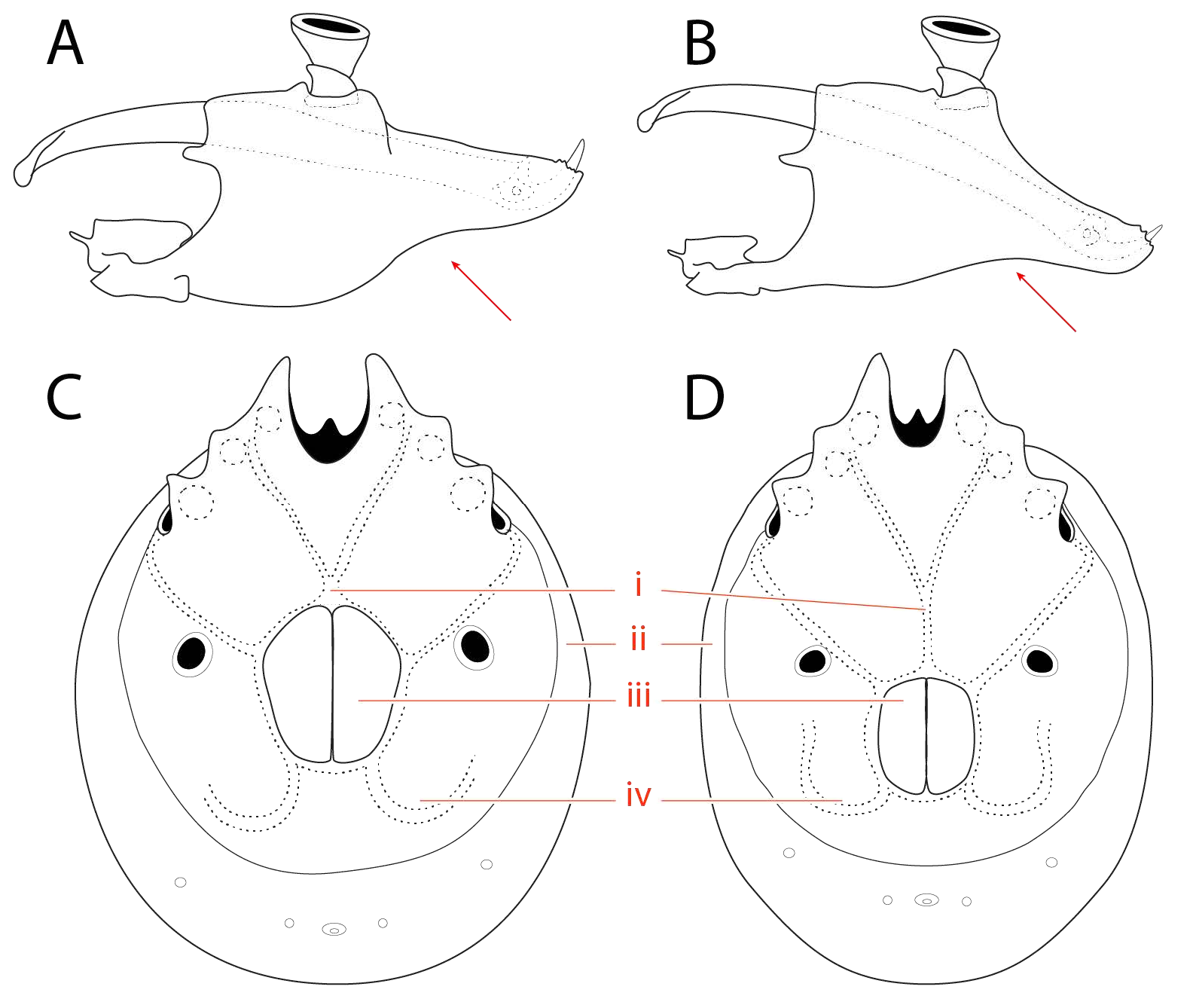
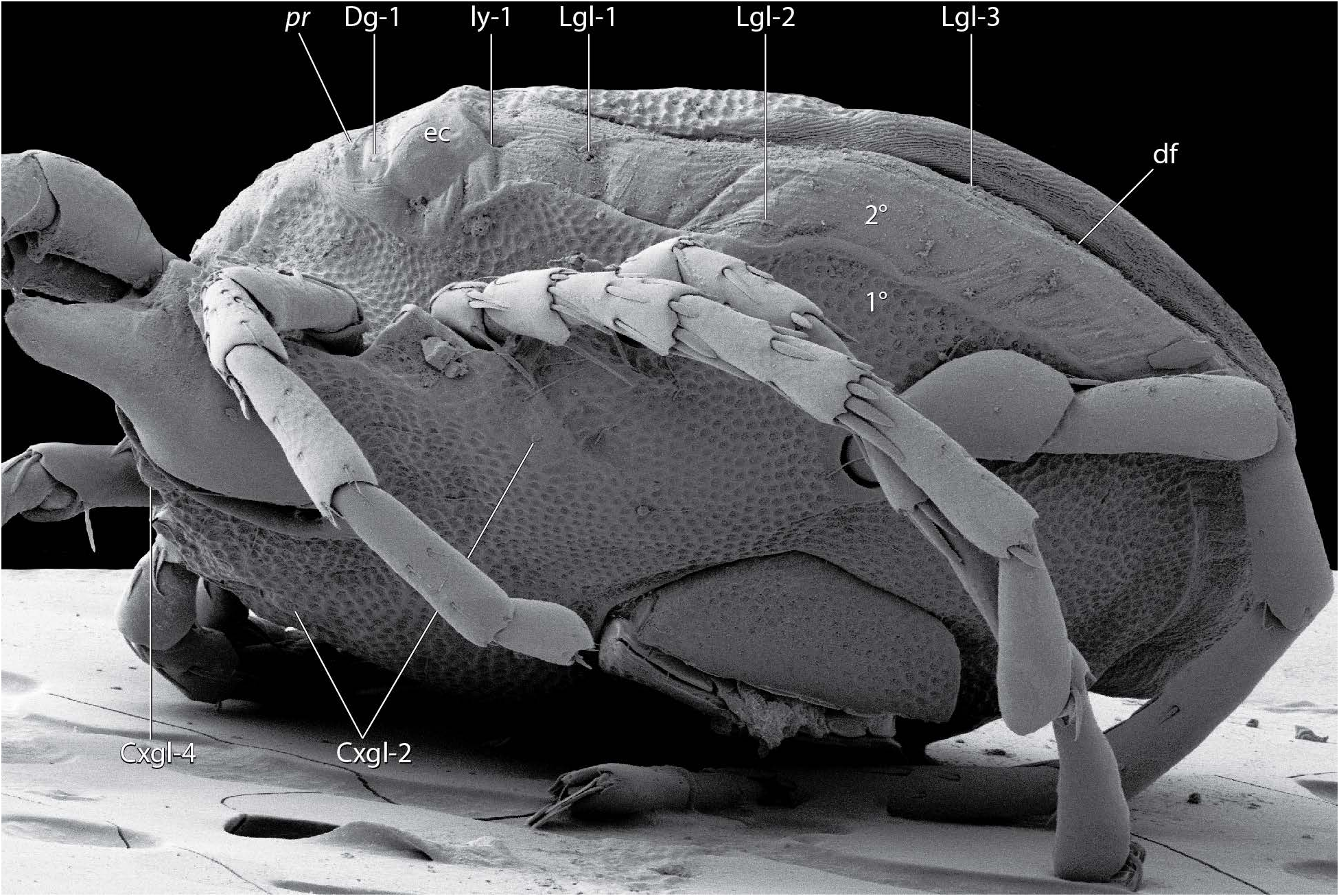
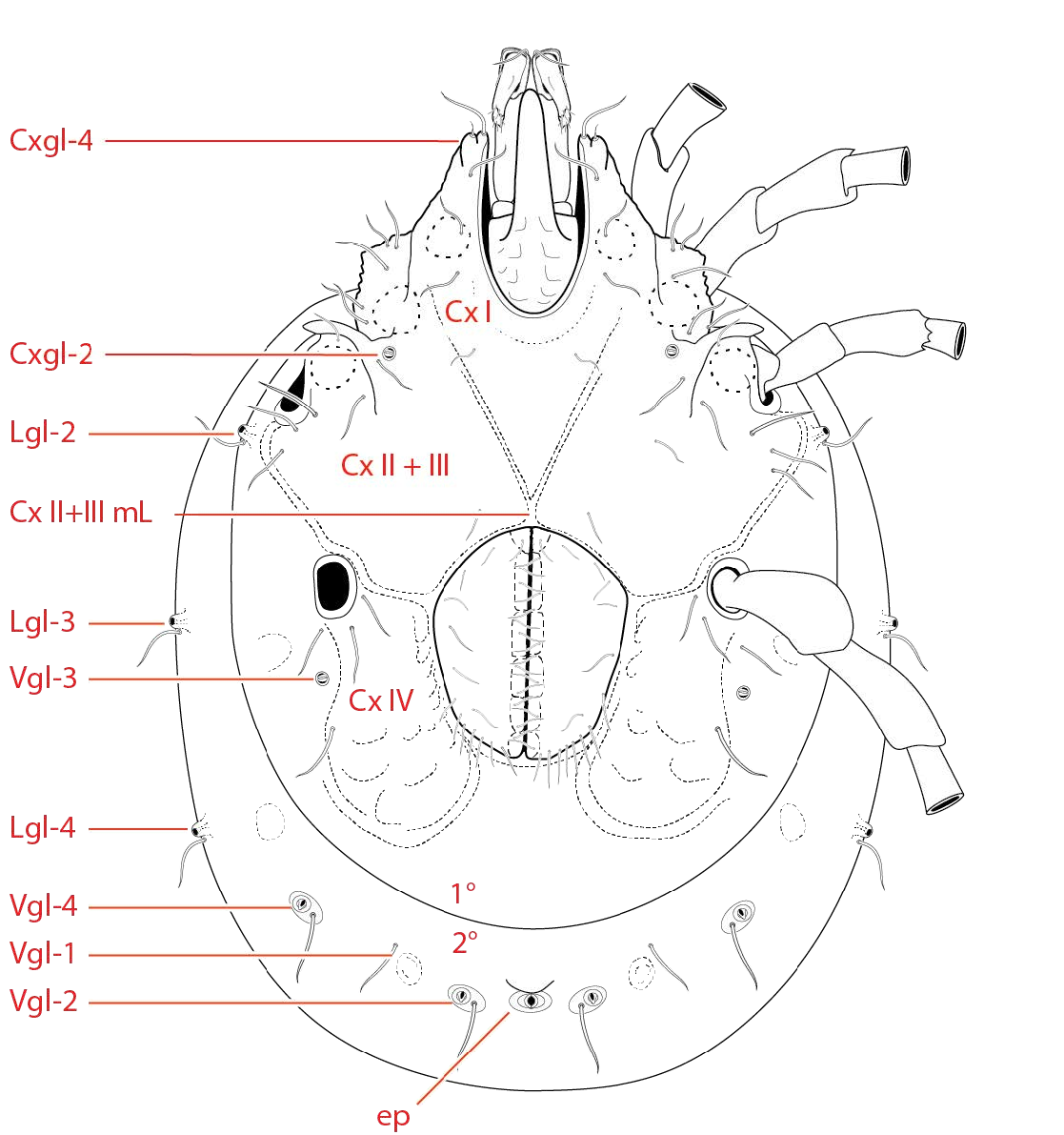
Dorsum ( Fig. 15-19 View FIGURE View FIGURE View FIGURE View FIGURE View FIGURE ) — [560 – 765 (723) long; 415 – 596 (543) wide] round to ovoid; armored with a central dorsal plate that is divided into an area of primary sclerotization [435 – 530 (477) long; 365 – 521 (440) wide] and an area of secondary sclerotization posteriorly [extends dorsal plate length by 87 – 176 for a total dorsal plate length of 522 – 706 (653)]. Specimens that have recently emerged from the imagochrysalis (i.e., teneral) have not yet developed the area of secondary sclerotization ( Fig. 6 View FIGURE ). The dorsal plate is bordered by ten platelets: two anterio-medials [109 – 178 (130) long; 53 – 86 (69) wide]; two anterio-laterals [135 – 220 (178) long; 65 – 101 (86) wide]; and a posterior ring of six smaller platelets in a striated membranous fold (partially visible in Fig. 18 View FIGURE ). Dgl-4 slightly lateral to Dgl-5 and usually in the area of secondary sclerotization, but occasionally near edge of primary sclerotization.
Eyes are apparently paired and located within sclerotized capsules on the margin of the anteriomedial platelets and dorsal covering of the gnathosoma ( Fig. 16 View FIGURE , 17 View FIGURE , 19 View FIGURE , 21 View FIGURE ).
Venter ( Fig. 20-25 View FIGURE View FIGURE View FIGURE View FIGURE View FIGURE View FIGURE ) — [615 – 870 (830) long; 487
– 668 (601) wide] round, fully sclerotized, and di-
Fisher J.R. et al.
vided into primary and secondary areas of sclerotization. Gnathosomal bay [119 – 300 (126) long; 68 – 98 (74) wide] not narrow (length/width <3; 1.9 average). Cx-1 narrowed to blunt tip, bearing Cxgl-4 ventro-apically ( Fig. 19 View FIGURE , 20 View FIGURE , 22 View FIGURE A-B, 23). Medial length of Cx-II + Cx-III short, barely longer than wide [10 – 42 (28)]. Genital plates large [153 – 210 (177) long; 133 – 185 (157) wide] and trapezoidal, extending anteriorly beyond level of Leg IV. Each genital plate rimmed in small setae ranging from simple to slightly barbulate ( Fig. 24C View FIGURE ). Additional measurements as follows: Cx-I total length 190 – 307 (281); Cx-III width 319 – 410 (399); Cx-I medial length 109 – 155 (155); genital field to excretory pore 140 – 240 (202); genital field to cauda 128 – 342 (341). Ovipositor morphology unknown.
Legs — Podomere measurements as follows. Leg I (422 – 536 total length): trochanter 37 – 75, basifemur 74 – 114, telofemur 75 – 119 (88), genu 97 – 120 (116), tibia 105 – 135 (125), tarsus 94 – 123 (106). Leg II [450 – 541 total length (541)]: trochanter 35 – 75 (47), basifemur 70 – 110 (88), telofemur 67 –
(a-m p); dorsal glandularia (Dgl); dorsal furrow (df); dorsal plate (dp); eye capsule (ec) lateral glandularia (Lgl); muscle scars (ms);
pre-ocularial setae (pr); post-ocularial setae (po); and area of primary (1°) and secondary (2°) sclerotization. 119 (91), genu 92 – 128 (101), tibia 107 – 148 (118), tarsus 103 – 153 (130). Leg III [511 – 631 total length (596)]: trochanter 44 – 83 (44), basifemur 70 – 115 (87), telofemur 66 – 98 (74), genu 105 – 143 (122), tibia 123 – 168 (145), tarsus 121 – 175 (150). Leg IV [726 – 867 total length (846)]: trochanter 90 – 138 (111), basifemur 108 – 170 (127), telofemur 103-145 (111), genu 139 – 190 (172), tibia 159 – 213 (184), tarsus 155 – 210 (187).
MALE (n=37) (allotypic measurements in parentheses when available) similar to female, except with sexually dimorphic characters discussed above, and with following specifications.
Gnathosoma — Subcapitulum [225 – 289 (249) ventral length; 168 – 215 (195) dorsal length; 93 – 134 (108) tall] posterior edge nearly vertical, ventral bend depth slight [5 – 15 (6)], and with short rostrum [84 – 128 (102) long] that is directed slightly downwards. Chelicerae [200 – 285 (251) long; 13 – 22 (15) high] unmodified with strongly curved fangs [37 – 58 (49) long]. Pedipalps [222 – 290 (263) long] with ventral projections and chaetotaxy as in female. Podomere measurements as follows: trochanters 23 – 34 (30) long and 24 – 37 (29) wide; femora 73 – 101 (91) long and 33 – 59 (47) wide; genua 57 – 81 (69) long and 36 – 52 (45) wide; tibiae 52 – 106 (87) long and 21 – 35 (30) wide; tarsi 18 – 34 (31) long and 9 – 15 (141) wide.
Dorsum — [520 – 650 (590) long; 366 – 495 (430) wide] ovoid to narrow. Dorsal plate with area of primary sclerotization [369 – 517 (429) long; 315 – 422 (367) wide] and an area of secondary sclerotization posteriorly [extends dorsal plate length by 107 – 148 (115) for a total dorsal plate length of 476 – 570 (545)]. Anterior platelets as follows: anteriomedials 96 – 120 (100) long and 49 – 75 (61) wide; anterio-laterals 148 – 190 (161) long and 65 – 90 (74) wide.
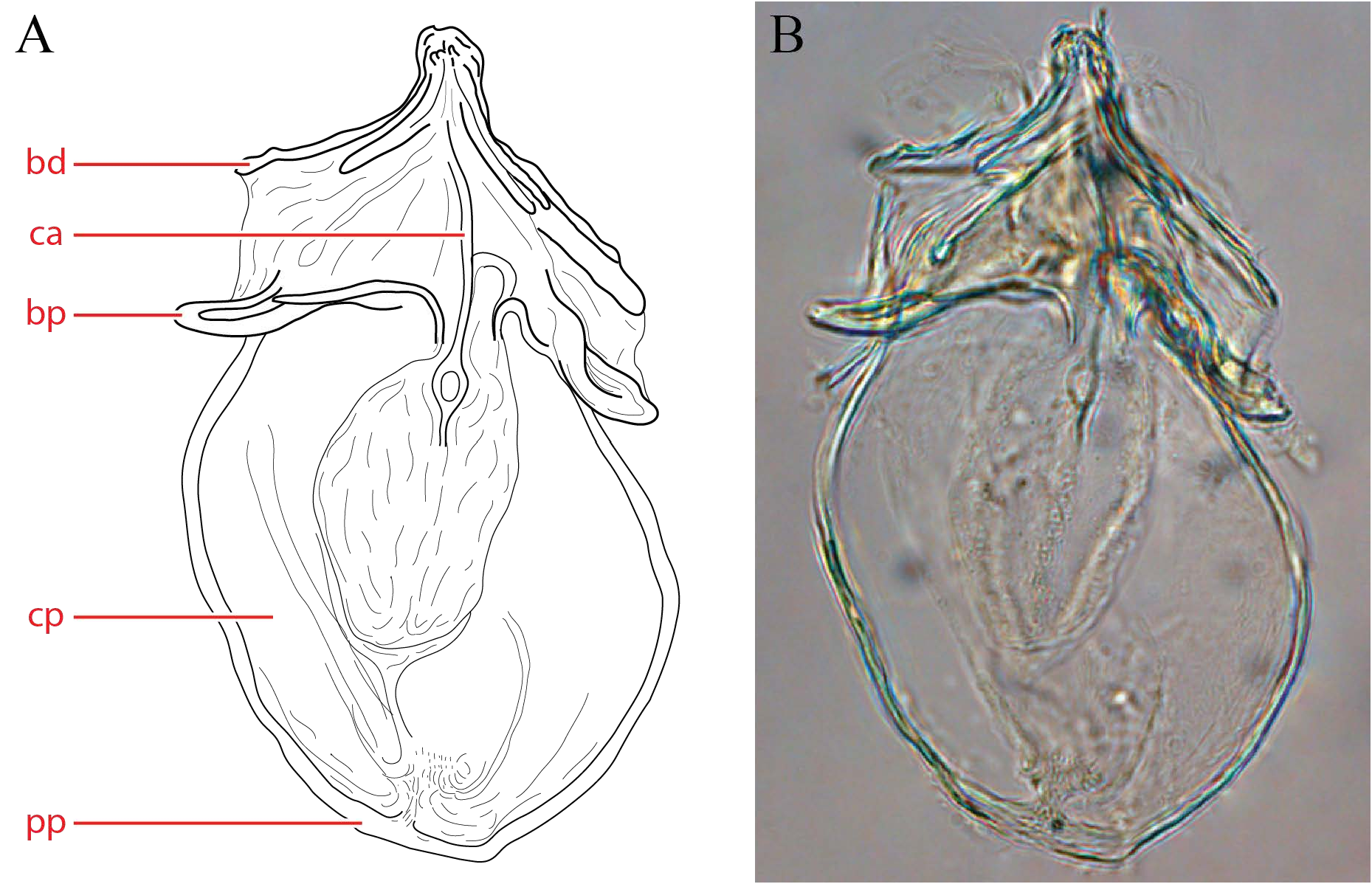
Venter — [589 – 800 (715) long; 432 – 605 (568) wide] ovoid to narrow. Gnathosomal bay [70 – 125 (100) long; 49 – 85 (76) wide] not narrow (length/width <3; 1.6 average). Medial length of Cx-II + Cx-III long [58 – 123 (86)]. Genital plates small [105 – 150 (131) long; 74 – 125 (95) wide] and rectangular, not extending anteriorly beyond level of Leg IV. Additional measurements as follows: Cx-I total length 212 – 297 (261); Cx-III width 387 – 371 (349); Cx-I medial length 126 – 167 (152); genital field to excretory pore 73 – 140 (128); genital field to cauda 150 – 242 (220). Genital skeleton ( Fig. 25 View FIGURE ) apically short, broad, and tapering abruptly. Cella proximalis large, with reduced processus proximalia; branchia proximalia well-developed, but branchia distalia only moderately developed.
Legs — Podomere measurements as follows. Leg I (410 – 515 total length): trochanter 40 – 70, basifemur 60 – 103, telofemur 74 – 96, genu 81 – 114 (90), tibia 85 – 125 (116), tarsus 88 – 110. Leg II [394 – 517 total length (484)]: trochanter 28 – 78 (71), basifemur 56 – 103 (85), telofemur 61-84 (69), genu 85 – 110 (90), tibia 93 – 135 (116), tarsus 101 – 133 (121). Leg III [431 – 578 total length (543)]: trochanter 31 – 73 (56), basifemur 55 – 103 (74), telofemur 64 – 89 (83), genu 90 – 125 (103), tibia 110 – 150 (129), tarsus 112 – 158 (133). Leg IV [678-805 total length (787)]: trochanter 89 – 128 (110), basifemur 93 – 155 (119), telofemur 95 – 128 (117), genu 138 – 170 (153), tibia 151 – 190 (177), tarsus 143 – 188 (173).
IMMATURES — unknown.
Etymology — Torrenticola (torrens -, L. a torrent; - colo, L. inhabitant) translates to "torrent dwellers" and refers to the lotic habitat of most species. The specific epithet trimaculata (tres -, L. three; - maculo, L. spotted) refers to the three dorsal spots of adults.
Habitat — Rocky and sandy areas (especially riffles) of healthy streams.
Distribution — Eastern North America. Given the breadth of material examined from the western US, it can be confidently concluded T. trimaculata is absent west of the 100 th meridian. The species is probably unable to cross the Great Plains, which should be considered the western-most bor- der. Further, it seems to be absent from the southeastern coastal plains. The species is most common in the Appalachian Mountains and Interior Highlands (Ozark and Ouachita Mountains), where it can be the dominant mite in a stream. Morph-1, the more pigmented morph ( Fig. 2A, C, E View FIGURE ), is known only from the Interior Highlands. It is the dominant morph in the Ozark Mountains, but is less common than Morph- 2 in the Ouachita Mountains. Morph- 2, the less-pigmented morph ( Fig. 2B, D, F View FIGURE ), is the only morph east of the Mississippi River. Morphs do not coexist within a given stream.
Common name — Three-spotted torrent mite.
Remarks — Torrenticola trimaculata are easily differentiated from other Torrenticola in eastern North America by the three distinct spots on the dorsal plate ( Fig. 1 View FIGURE , 2 View FIGURE , 9 View FIGURE ). Additionally, the following characters are important in combination: anterior platelets not fused with dorsal plate; rostrum short (less than maximum depth of subcapitulum) and sexually dimorphic (angled downward in males); ventral bend of female subcapitulum slight; pedipalpal femora and genua bear ventral dentate projections that are not lamellate; hind coxae extend posteriorly beyond the genital plates in females (in line with genital plates in males); and medial suture of females short. Torrenticola trimaculata sp. nov. exhibit considerable variability across many character states, such as coloration, especially within Morph- 2 ( Fig. 9 View FIGURE ).
We identified four anomalies that are worth reporting. Occasionally, individuals of Morph-1 were found with reddish coloration of the gnathosoma and surrounding sclerites ( Fig. 26A View FIGURE ). Although size and shape of the dorsal spots varied considerably, two extremes were identified: 1) spots enlarged so much so that the posterior spots merged into a contiguous U-shape, only found in Morph- 1 males ( Fig. 26B View FIGURE ); and 2) spots so much reduced that the posterior spots were merely comma-shaped, found
Fisher J.R. et al.
in both sexes of Morph-2 ( Fig. 26C View FIGURE ). Finally, one individual had an under-developed anterio-medial platelet ( Fig. 26 View FIGURE D-E).
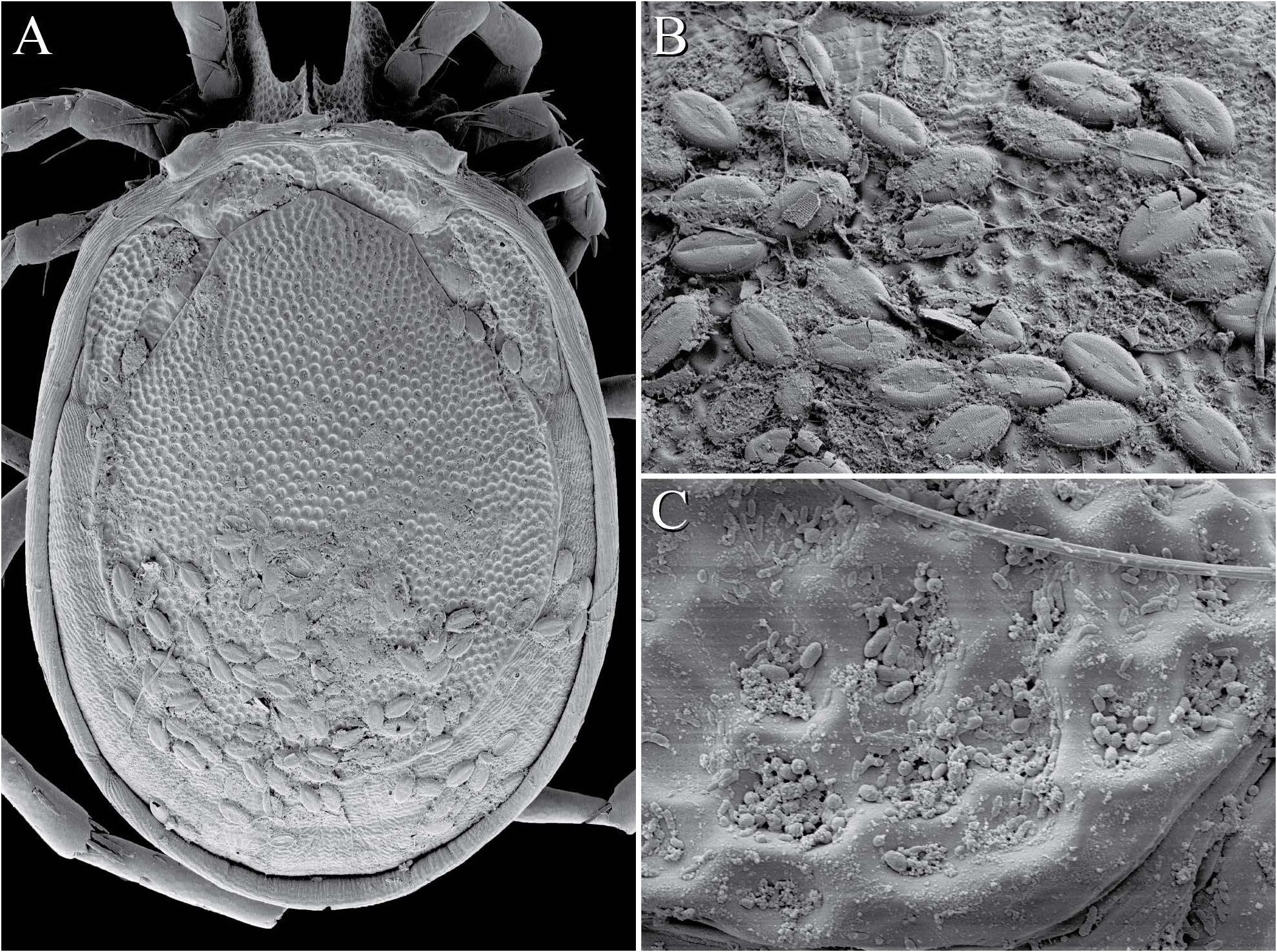
Samples often contained at least a few mites with epibionts. Several suctorian ciliates (Ciliophora: Suctorea) are known epibionts on aquatic arthropods, including mites ( Dovgal and Peši´c 2007, 2012, Dovgal et al. 2008), and our samples occasionally contained small numbers of mites covered in unidentified suctorians. Additionally, unidentified bacteria were surprisingly abundant on the integument surface, especially within depressions ( Fig. 22 View FIGURE B-D, 27C).
Commonly, mites were covered with epiphytic diatoms identified by Andy Alverson, a diatom spe- cialist at the University of Arkansas, as Cocconeis placentula Ehrenberg, 1838 ( Fig. 27 View FIGURE A-B). Cocconeis are common epibionts well-known for adhering to plants and algae (e.g., Sand-Jensen 1977, Ferreira and Seeliger 1985, Hardwick et al. 1992, Siqueiros- Beltrones et al. 2002). However, few records exist of these diatoms adhering to animals; but see Siqueiros-Beltrones et al. (2001) for a record of C. notata Petit, 1877 living inside the body of a hydrozoan that itself was epizootic on giant kelp, Macrocystis pyrifera (L.) CA Agardh. The present report represents the first record of C. placentula as epiphytic on water mites.
Type series — HOLOTYPE (♀): USA, Arkansas, Madison Co., Withrow Springs State Park, War Eagle Creek (36°8’59.3" N, 93°44’26.94" W), 27 Jul 2011, GoogleMaps
by IM Smith, IMS110034.
ALLOTYPE (♂): USA, Arkansas, Madison County, Withrow Springs State Park, War Eagle Creek (36°8’59.3" N, 93°44’26.94" W), 27 Jul 2011, by IM Smith, IMS110034 View Materials . GoogleMaps
PARATYPES (48♀; 36♂): Arkansas, USA : 2♀ and 3♂ from Madison County, Withrow Springs State Park , War Eagle Creek (36°8’59.3" N, 93°44’26.94" W), 27 Jul 2011, by IM Smith, IMS110034 View Materials GoogleMaps 1♀ from Marion County, Crooked Creek ex. Northern hogsucker ( Hypentelium nigricans ) (36°15’9.9" N, 94°26’25.8" W), 22 Jul 2014, by CT McAllister GoogleMaps
3♀ and 2♂ from Montgomery County, Ouachita National Forest , Ouachita River (34°34’53.20" N, 93°53’0.16" W), 5 Oct 2007, by AJ Radwell and HW Robison, AJR070300A GoogleMaps 8♀ and 5♂ from Montgomery County, Ouachita National Forest , South Fork of Ouachita River, 29 Jul 2011, by AJ Radwell and B Crump, AJR110302 2♀ and 1♂ from Montgomery County, Ouachita National Forest , Ouachita River, 27 Aug 2011, by AJ Radwell, AJR110307 4♀ and 4♂ from Montgomery County, Ouachita National Forest , South Fork of Ouachita River, 29 Jul 2011, by IM Smith, IMS110040 View Materials 1♀ from Montgomery County, Caddo River , 29 Jul 2011, by IM Smith, IMS110037 View Materials 1♂ from Newton County, Ozark National Forest , Mill Creek (36°3’42.12" N, 93°8’7.62"W), 20 Jun 2012, by TD Edwards, TDE 12-0620-010 GoogleMaps 2♀ and 2♂ from Newton County, Ozark National Forest , Little Buffalo River, 2 Sep 2012, by TD Edwards, TDE 12-0902- 003 1♂ from Newton County, Buffalo National River , Whiteley Creek (35°59’28.14" N, 93°23’57.24" W), 23 May 2012, by TD Edwards, TDE 12-0523- 002 Illinois, USA GoogleMaps : 2♀ and 1♂ from Union County, Clear Creek (37°33’ N, 89°23’ W), 13 Sep 1991, by IM Smith, IMS910036 View Materials A Indiana, USA GoogleMaps : 1♀ from Wayne County (39°51’13" N, 85°8’4" W), 24 Jul 2014, by MJ Skvarla, MS 14-0731-001 Georgia, USA GoogleMaps : 1♀ from Chattooga County, Johns Creek (34°34’ N, 80°5’ W), 4 Jul 1990, by IM Smith, IMS900076 View Materials Kentucky, USA GoogleMaps : 1♀ and 2♂ from McCreary County, Rock Creek (36°42’ N, 84°36’ W), 8 Jul 1990, by IM Smith, IMS900082 View Materials B Michigan, USA GoogleMaps : 2♀ and 2♂ from Barry County, Thornapple River (42°39’ N, 85°17’ W), 29 Jul 1959, by DR Cook, DRC GoogleMaps 590034 Missouri, USA: 2♀ and 1♂ from Crawford County, Huzzah Creek , 23 Jul 2011, by IM Smith, IMS110029 View Materials
New York, USA: 3♀ and 1♂ from St. Lawrence County, Canton (44°35’ N, 75°10’ W), 15 May 1986, by BP Smith, BPS860508 GoogleMaps 1♀ from USA, New York, Delaware Co. , Roscoe (41°55’ N, 74°54’ W), 11 June 1988, by PW GoogleMaps Schefter and R MacCulloch , IMS880110 View Materials
Nova Scotia, Canada: 1♀ from Victoria County, Baddeck River (44°52’ N, 61°5’ W), 18 Jul 1981, by IM GoogleMaps Smith , IMS810082 View Materials Ontario, Canada: 4♀ and 2♂ from Grey County, Saugeen River (44°10’ N, 80°49’ W), 9 Jun 1989, by IM GoogleMaps Smith , IMS890028 View Materials A 1♀ from Madoc (44°30’ N, 77°28’ W), 4 May 1980, by IM GoogleMaps Smith , IMS800003 View Materials A 1♂ from Renfrew County, Madawaska River (45°21’ N, 76°40’ W), 25 May 1980, by IM GoogleMaps Smith , IMS800012 View Materials 1♀ and 1♂ from Lanark County , Mississippi River (45°3’ N, 76°23’ W), 6 Oct 1983, by IM GoogleMaps Smith and CJ Hill , IMS830093 View Materials A Virginia, USA: 1♀ and 1♂ from Scott County, North Fork of Holston River (36°39’ N, 82°28’ W), 7 Jul 1990, by IM GoogleMaps Smith , IMS0900080 View Materials 2♀ and 4♂ from Alleghany County, Potts Creek (37°44’ N, 80°2’ W), 13 Jul 1990, by IM GoogleMaps Smith , IMS900091 View Materials B 1♀ and 1♂ from Bath County, Jackson River (38°8’ N, 79°46’ W), 16 Jul 1990, by IM GoogleMaps Smith , IMS900100 View Materials West Virginia, USA: 2♀ from Pendleton County, North Fork of South Branch of Potomac River (39°0’ N, 79°22’ W), 17 Jul 1990, by IM GoogleMaps Smith , IMS900104 View Materials .
Type deposition — Holotype (♀), allotype (♂), and 50 (30♀; 20♂) paratypes deposited at the CNC; 4♀ and 4♂ paratypes deposited at the ACUA; 4♀ and 4♂ paratypes deposited at the OSUAC; 4♀ and 4♂ paratypes deposited at the GMNH. The holotype and allotype are slide mounted in Hoyer’s medium; paratypes are a mixture of Hoyer’s and glycerin jelly slide mounts.
















| IM |
Indian Museum |
| AJ |
Central Research Laboratories |
| PW |
Paleontological Collections |
| R |
Departamento de Geologia, Universidad de Chile |
| CJ |
Centro Jambatu de Investigación y Conservación de Anfibios, Fundación Otonga |
| CNC |
Canadian National Collection of Insects, Arachnids, and Nematodes |
| GMNH |
Georgia Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |