
Megalomma phyllisae, Capa & Murray, 2009
|
publication ID |
https://doi.org/ 10.3853/j.0067-1975.61.2009.1529 |
|
persistent identifier |
https://treatment.plazi.org/id/2202BF4F-FFC8-5B78-FC4A-50EFFECEF809 |
|
treatment provided by |
Felipe |
|
scientific name |
Megalomma phyllisae |
| status |
sp. nov. |
Megalomma phyllisae View in CoL n.sp.
Figs 2A–E View Figure 2 , 3 View Figure 3 , 4A,B View Figure 4 , 5A View Figure 5
Material examined. Victoria. HOLOTYPE AM W13643, off Townsend Point, Corner Inlet, 38°48'S 146°33'E, 16 Dec. 1976, 1.5 m GoogleMaps . PARATYPES AM W35476 (2 spec., fragment from incomplete spec. on SEM stubs AM M082 and M083), same locality GoogleMaps .
Additional material. Megalomma suspiciens ( Ehlers, 1904) ZMUH PE 1304 , syntypes (4 spec.), French Pass, New Zealand, 1 m, 1897. Megalomma trioculatum Reish, 1968 USNM 38409, paratypes (2), lagoon side of Engebi Island, Eniwetok Atoll, Marshal Islands, 7 Sep. 1956.
Diagnosis. Species characterized by a combination of the following features: subdistal compound eyes in all except ventralmost radioles, radiolar skeleton with more than 25 cells in transverse section, dorsal margins of collar fused to faecal groove forming flanking pockets on each side, caruncle absent, and inferior thoracic chaetae with progressively tapering distal tip (type B).
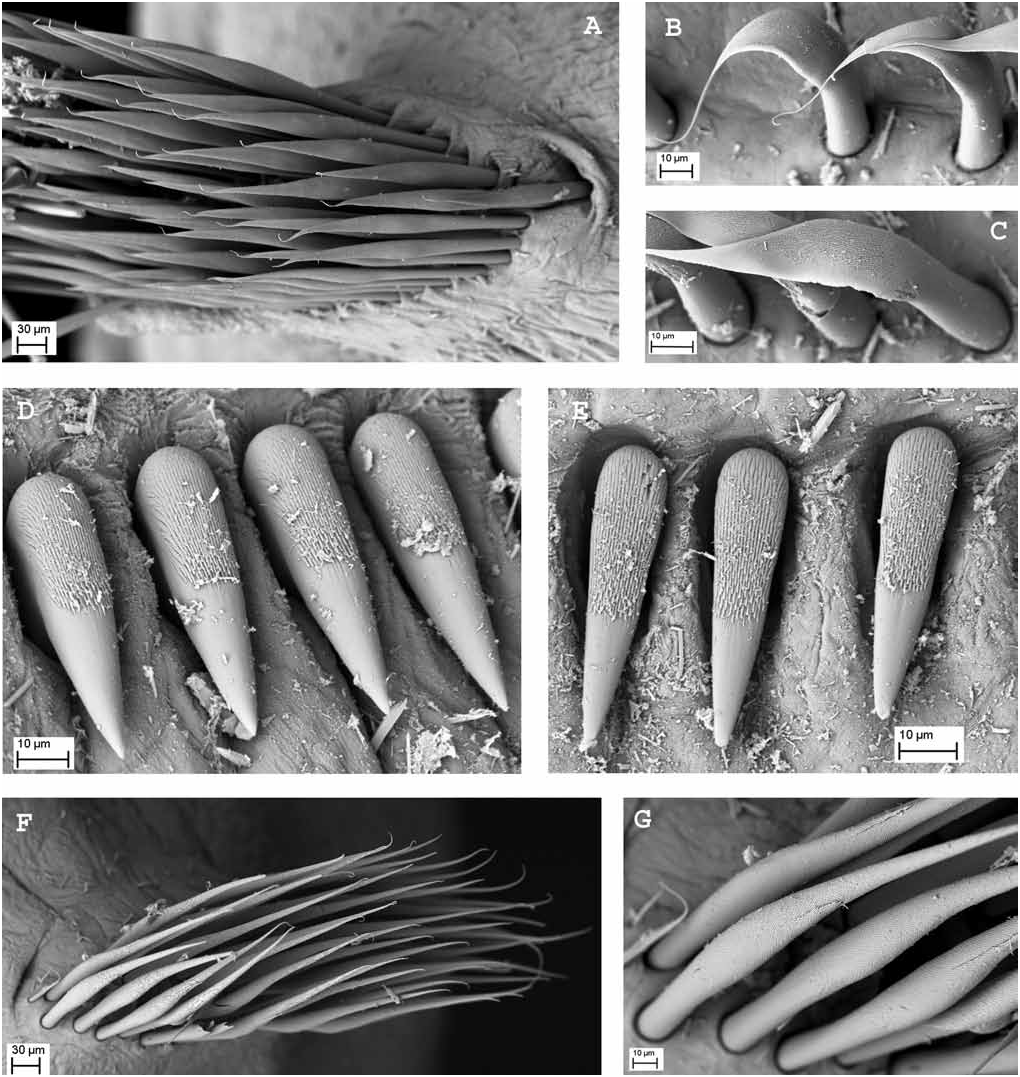
Description. Holotype 135 mm long, 5 mm wide; with eight thoracic and 107 abdominal chaetigers. Crown slightly longer than thorax, 15 mm long, with 30 radioles on left side and 28 on right side arranged in two semicircles. External margin of radioles quadrangular ( Fig. 2A,B View Figure 2 ). Tip of radioles shorter than pinnules ( Fig. 2B View Figure 2 ). Radiolar skeleton composed of more than 25 cells in transverse section ( Fig. 5A View Figure 5 ). All radioles except for last two pairs of ventral radioles with a subdistal compound eye, eyes diminishing in size to ventral radioles, spiral in shape on most radioles (although difficult to determine shape in small eyes). Dorsal lips with radiolar appendages as long as two thoracic chaetigers; with three dorsal pinnular appendages each. Caruncle absent. Low smooth keel (thickened and non-lamellate) projecting ventrally between dorsal lips, arising from raised triangular mound situated mid dorsal to dorsal lips. Ventral lips rounded and well developed; parallel lamellae and ventral sacs present ( Fig. 2D View Figure 2 ). Posterior peristomial collar fused mediodorsally to the faecal groove, with short dorsal lappets equal in length to lateral collar margins, and with shallow dorsolateral U-shaped incisions on both sides forming deep pockets, reaching to second chaetiger ( Fig. 2C View Figure 2 ); lateral collar margins smooth, and ventrally forming overlapping lappets with rounded anterior margins ( Fig. 2D View Figure 2 ). Ventral shields quadrangular, separated from the neuropodial tori, all similar in width. First ventral shield longer than the rest, with m-shaped anterior margin ( Fig. 2D View Figure 2 ). First chaetiger with superior and inferior elongate narrowly hooded notochaetae; superior longer than inferior. Rest of thoracic chaetigers with about 20 elongate narrowly hooded superior chaetae and several irregular rows of broadly hooded inferior notochaetae progressively tapering distally (type B) ( Fig. 3A View Figure 3 ), in fascicles separated by large lamellate interramal lobe. Neuropodial tori slightly diminishing in width posteriorly. Uncini with around 13 rows of small and similarly sized teeth above main fang ( Fig. 3D View Figure 3 ), well developed breast, handle about 3.5 times the length of the distance from breast to main fang ( Fig. 4A View Figure 4 ). Companion chaetae with asymmetrical membrane with most of surface covered with fine teeth ( Fig. 3B,C View Figure 3 ). Abdominal neuropodia with slender broadly hooded chaetae ( Fig. 3F,G View Figure 3 ). Abdominal notopodial uncini with crest of teeth similar to thoracic uncini ( Fig. 3E View Figure 3 ) but with handles half the length of thoracic ones ( Fig. 4B View Figure 4 ). Pygidium damaged ( Fig. 2E View Figure 2 ). Tough chitinous tube present, embedded externally with shell fragments.
Colour pattern. Body unpigmented, radioles purple, with around three-four irregular broad transverse bands; proximal band most intense and largest, with more irregular patches distally on radioles ( Fig. 2A View Figure 2 ).
Variation. Complete paratype, 95 mm long, 6 mm wide, with eight thoracic and 120 abdominal chaetigers; pygidium damaged. Branchial crown with 30 pairs radioles, 24 pairs with eyes. Incomplete paratype, with eight thoracic and c. 10 abdominal chaetigers. Branchial crown with 25 and 27 radioles in right and left lobes; with subdistal eyes missing from ventralmost 5–7 pairs of radioles. Colour pattern similar in all three specimens.
Reproductive features. Incomplete paratype with oocytes in the ten anterior abdominal segments (rest of abdominal chaetigers missing).
Etymology. The name of this species is given in dedication to Dr Phyllis Knight-Jones in recognition of her great contribution to sabellid taxonomy.
Remarks. This new species shares some characters with others described from the Indo-Pacific: M. suspiciens ( Ehlers, 1904) recorded from New Zealand, and M. multioculatum Fitzhugh, 2002 , from Thailand. These characters are the dorsal margins of collar fused to the faecal groove, dorsolateral collar pockets present, and subdistal eyes present on most branchial radioles.
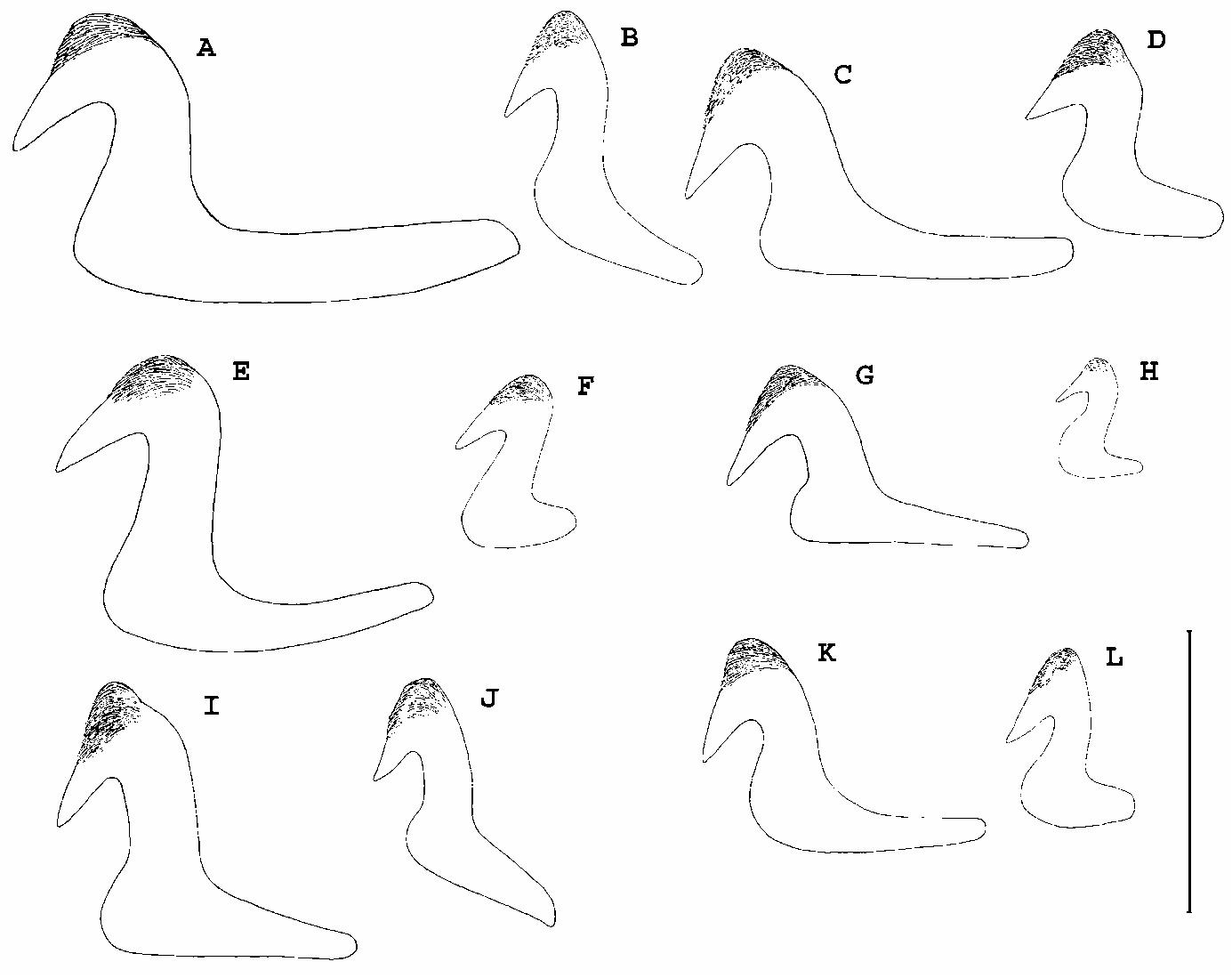
Megalomma phyllisae n.sp. and M. suspiciens ( Ehlers, 1904) both possess the same type of inferior thoracic chaetae (type B) as well as long-handled thoracic uncini. Furthermore, they share the presence of a smooth, rounded and dorsoventrally-directed “keel” between the dorsal lips, a feature observed after examining the types of M. suspiciens and which Tovar-Hernández & Salazar-Vallejo (2008) misinterpreted as a caruncle (p. 1957). Both taxa also lack interramal “eyespots” although they were recorded as present in M. suspiciens by Tovar-Hernández & Salazar-Vallejo (2008 p. 1957), probably mistakenly, as they subsequently state these are absent on p. 1961. Megalomma phyllisae n.sp. possesses eyes on most of the radioles but in M. suspiciens they are only present in 8–9 of 16–18 pairs of radioles (less than half) and the eyes are smaller in the new species. Collar margins have more shallow dorsolateral incisions and deeper pockets compared with those of M. suspiciens , and short dorsal lappets are present in M. phyllisae n.sp. and absent in M. suspiciens . The new species differs from M. suspiciens in the length of the thoracic uncini handles, with approximately 3.5 times the length of the distance from main fang to breast in M. phyllisae n.sp., compared with about twice the length form breast to fang in M. suspiciens ( Fig. 4K View Figure 4 ), and also in the length of abdominal uncini handles ( Figs 4B,L View Figure 4 ). Other minor differences include the presence of three pinnular appendages per dorsal lip in M. phyllisae n.sp. compared with a single pinnular appendage in M. suspiciens , the development of the ventral lappets which are long and overlapping in M. phyllisae n.sp. and short and non-overlapping in M. suspiciens , and the relative position of the thoracic ventral shields to the neuropodial tori with obvious gaps between tori and shields in M. phyllisae n.sp. and which abut in M. suspiciens .
Megalomma phyllisae n.sp. and M. multioculatum differ by the shape of the inferior thoracic notochaetae type B in the new species and type A in M. multioculatum ). Other differences include the number of rows of cells in the radiole structure, with more than 25 in M. phyllisae n.sp. whereas there are four rows of cells in M. multioculatum ; and the development of dorsal lappets, low in M. phyllisae n.sp. and absent in M. multioculatum .
Another species recorded in the Indo-Pacific that shares similar features is M. trioculatum Reish, 1968 , from Marshall Islands, but after reviewing the paratypes ( Fig. 2F–I View Figure 2 ) there is no doubt that there is a misinterpretation of morphology in the original description of this species. The dorsal margins of collar are clearly separated from the faecal groove, ending laterally and abruptly, and not producing any pockets on the sides ( Fig. 2G View Figure 2 ). Therefore, M. trioculatum should now be included with those from Knight-Jones’ Group 2C ( Table 2). Megalomma phyllisae n.sp. also is distinguished from M. trioculatum in the shape of the ventral lappets, rounded in M. phyllisae n.sp. and pointed in M. trioculatum ( Fig. 2F,H View Figure 2 ), and in the length of the thoracic uncini handles (which are much shorter in M. trioculatum ). Also, the relative number of radiolar eyes is greater in M. trioculatum as all radioles of the described holotype and the two paratypes examined possess subdistal eyes in all radioles. The number of rows in the radiolar skeleton of M. phyllisae n.sp. is higher than in M. trioculatum which only bears four ( Fig. 5A,G View Figure 5 ). It also has to be mentioned that M. trioculatum does not show three types of eyes, as stated in the original description; the two paratypes examined possess radiolar eyes only, and these are of two sizes, those in dorsalmost radioles are large and almost surrounding the whole radiole ( Fig. 2I View Figure 2 ), whereas the remaining radioles bear small eyes, all similar in size.
Other Megalomma species resemble the new species in the shape of collar dorsal margins and the presence of eyes in most radioles. Some of these species can be distinguished from M. phyllisae n.sp. by the presence of a caruncle, absent in the new species. These are M. lobiferum ( Ehlers, 1904) and M. carunculata Tovar-Hernández & Salazar-Vallejo, 2008 . The new species has short dorsal lappets equal in length to lateral collar margins, while other species possess long dorsal lappets. These are M. acrophthalmos ( Grube, 1878) , M. fauchaldi Giangrande et al., 2007 , M. vesiculosum ( Montagu, 1815) and M. claparedii ( Gravier, 1908) as described by Giangrande and Licciano (2008). Megalomma phyllisae n.sp. is distinguished from M. circumspectum ( Moore, 1923) in the arrangement of radiolar eyes which are lacking in some ventral radioles in the new species and present in all radioles in M. circumspectum ( Hartman, 1969) ; thoracic uncini also differ in the two species—uncini handles of M. phyllisae n.sp. are approximately twice as long as those of M. circumspectum .


| AM |
Australian Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |