
Pontoscolex corethrurus
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4545.1.7 |
|
publication LSID |
lsid:zoobank.org:pub:6FB28E3D-D647-4E9B-BD7D-357677ACC6AB |
|
DOI |
https://doi.org/10.5281/zenodo.5931904 |
|
persistent identifier |
https://treatment.plazi.org/id/1D7C87EF-9E5B-FFD2-15E8-FB67FCA3558D |
|
treatment provided by |
Plazi |
|
scientific name |
Pontoscolex corethrurus |
| status |
|
Lumbricus corethrurus Müller, 1857a: 113 ; 1857b: 13. [Type locality Itajahy (Itajaí), now Blumenau, Santa Catarina, Brazil. Types unknown]. Pontoscolex corethrurus: Taheri et al. 2018 : lineage 1 (L1), p. 66.
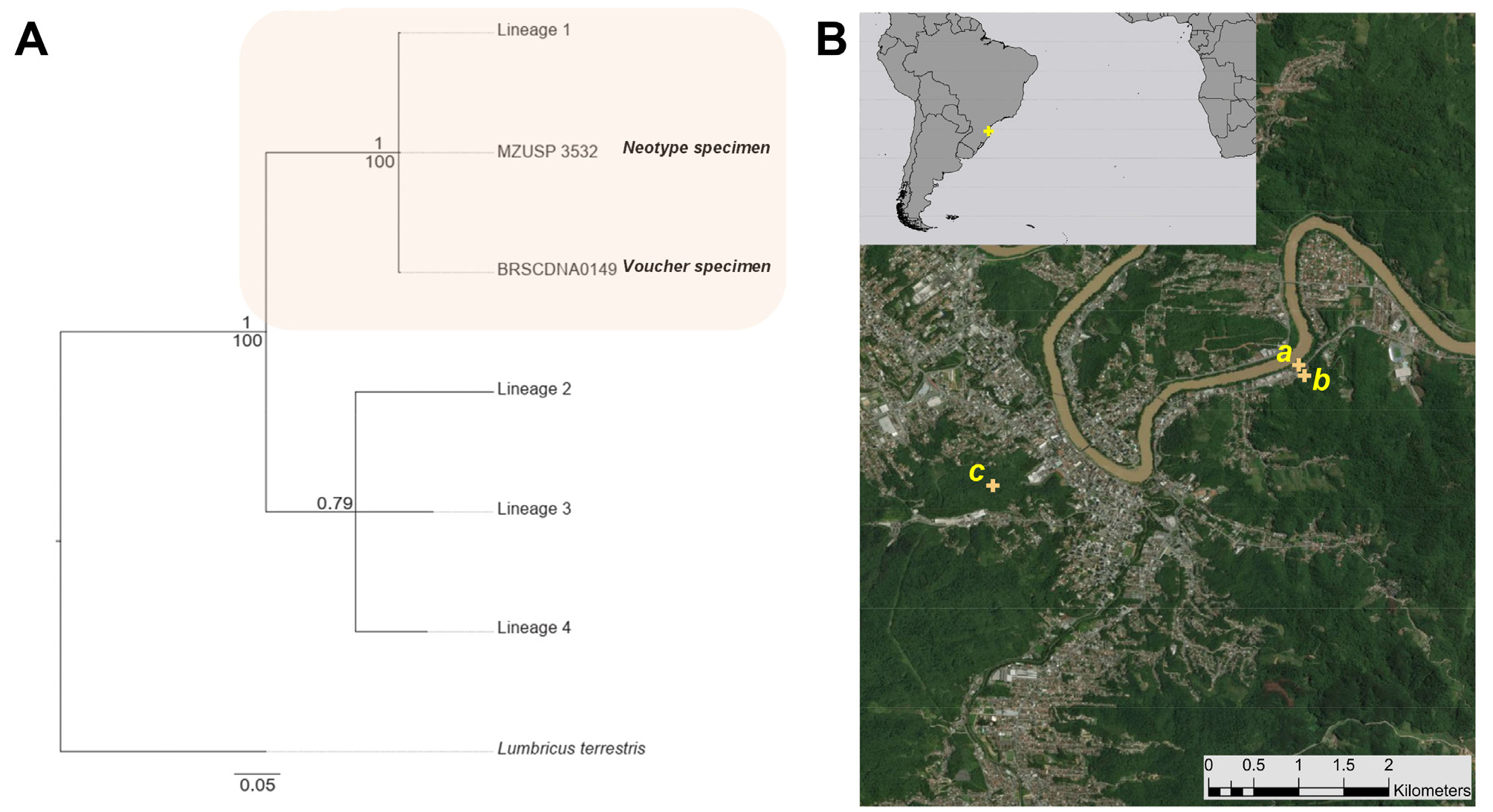
Neotype. MZUSP 3532 View Materials one adult. Type locality. Grass lawn behind the Museu de Ecologia Fritz Müller (former house of Fritz Müller), Blumenau, Santa Catarina, Brazil, 26° 54' 35.499"S 49° 2' 39.3714"W, 37 masl. 22 April 2016, M.L.C. Bartz coll. GenBank MH 717712 View Materials . GoogleMaps
Other material. COFM BRSCDNA0149 : one adult. Same collection data as neotype locality ("voucher"). GenBank MH 717713 View Materials . COFM BRSCDNA0053 : Same collection data as neotype locality, GenBank MH 717714 View Materials . COFM BRSCDNA0046 adult GenBank MH 717710 View Materials , COFM BRSCDNA0047 late juvenile GenBank MH 717711 View Materials : Secondary forest . Other side of the road from the Museu de Ecologia Fritz Müller (former house of Fritz Müller), Blumenau, Santa Catarina, Brazil, 26° 54' 39.3228"S 49° 2' 37.5462"W, 37 masl. 12 November 2015, G.G. Brown, M.L.C. Bartz, L. Cunha, S. Taheri & E. da Silva colls GoogleMaps . COFM BRSCDNA0162 adult GenBank MH 801870 View Materials , COFM BRSCDNA0163 adult GenBank MH 801871 View Materials , COFM BRSCDNA0165 adult GenBank MH 801872 View Materials , COFM BRSCDNA0167 adult GenBank MH 801873 View Materials , COFM BRSCDNA0168 adult GenBank MH 801874 View Materials , COFM BRSCDNA0169 adult GenBank MH 801875 View Materials : Parque Natural Municipal São Francisco de Assis , Blumenau, Santa Catarina, Brazil, 26°55'18.8004"S 49° 4'29.0388"W, 86 masl. 22 April 2016, M.L.C. Bartz coll. GoogleMaps
Description. Dimensions, alcoholic neotype MZUSP 3532: 125 mm by 2.5 mm at x, 3 mm at clitellum, 2.0 mm at xxx, 200 segments; formalin-fixed voucher: 81 mm by 4.0 mm at x, 4.5 mm at clitellum, 3.8 mm at xxx, 210 segments. Body cylindrical. Setae a, b, c and d commence on ii. Setae closely paired in regular series ii–xi, AB distance gradually increasing from xi through clitellar segments; starting in xxiv setae b commence alternation of
position in which the row has a seta displaced dorsally in approximately every other segment. Setae a become irregular in lxvii, with regular alternation of position by lxxv; setae c become irregular in lxxxii with regular alternation by lxxxv; setae d become irregular in xii. In one individual (BRSCDNA0169) rows a and c began alternating in the l to liii range.
All setae attain the regularity of the quincunxial arrangement over segments cxxxiv–cxl and continue in this form to the tail. Caudal setae slightly curved, not hooked, and gradually enlarging starting from cxxxvi until reaching full size characteristic of the tail by cxlviii. Prostomium indistinct, segment i everted, segments ii–xii with equatorial ridge, secondary annulations present after xii until clitellum. Unpigmented/whitish, clitellum white. Spermathecal pores near c in 6/7–8/9, ovipores and male pores invisible; tubercula pubertatis ½ xix–½ xxii, clitellum annular xvi–xviii, saddle to just above b otherwise, ½ xv–½ xxiii. Nephropores in c line or its regular continuation. Special caudal zone of very short segments in cxxiv–cxxxii.
Septa 5/6 thin, 6/7–8/9 equally thick and muscular, septum 9/10 absent and 10/11 thicker. Alimentary canal with large cylindrical gizzard in vi; esophagus valvular in xiv, intestinal origin xvii; typhlosole origin 24/25 lamina in longitudinal diagonal folds. Intestine of xv–xxiii broader, with two or three pairs large blood vessels in intestinal wall of each segment, intestine constricted near 22/23, narrower from xxiii or xxiv posteriorly. Calciferous glands paired in vii, viii, ix, panicled-tubular type with generally parallel tubes following the gland axis, tongue-shaped with slight differentiation like a small crest on the distal end, pedunculated; blood vessels to gland include large branch of dorsal vessel to approximate center of each gland. Holonephric, vesiculate; ducts to body wall near level of c.
Vascular system with ventral trunk, single dorsal trunk, lateral vessels in vii–ix, esophageal hearts in x–xi. Extra-esophageal vessel from pharyngeal glands, along ventral-lateral face of gizzard, esophagus back to calciferous glands; supraesophageal vessel in vii–xi.
Ovaries not found; spermathecae vii, viii, ix, adiverticulate, clavate, empty. Male sexual system generally absent; seminal vesicles atrophied, extending to xvi. When male organs developed, only partially so and atrophied to varying degrees (see remarks).
Remarks. Müller's description (1857) is brief and lacks most of the details now standard. The neotype of 200 segments is at the low end of the range he gives (200–250), and other external characteristics are similar, other than his placement of the clitellum in xiv–xxi. That is explainable by missing an invaginated first segment. He states that setal rows a and c remain linear until about xl to lx, after which the rows become irregular, and even "chaotic," until the tail region, behind the special caudal zone occurring in the region of segments cx to cxx. After that the setae assume the regularity of the quincunxial pattern. Setae a become irregular slightly anterior to the irregularity of row c, which is consistent with our material, although our specimens mostly start irregularity more posteriorly (neotype: a in lxvii, c in lxxxii). The placements of the setae b and d are essentially the same in the neotype and in Müller (1857). For row b, it is generally within the range of xxiv to xxxv, but regular alternation might not begin for several more segments. Only the d row commences alternating positions consistently within a narrow range: xi to xiv.
There is uniformity on the shape and size of the setae within the quincunxial zone as also noted by Müller (1857), and the commencement of quincunx always a few tens of segments behind the special caudal zone of narrow segments. Tail setae are not hooked, in our and Müller's material. The caudal zone itself is with few exceptions in the range 122–135 in the L 1 specimens we viewed. Two had more anterior locations in 110–116 and 112–118, very much as in Müller (1857), with whose material ours agrees in the nature of the caudal zone segments: very short, of slightly greater body diameter, and lacking setae.
The neotype and all specimens named as voucher or other material were from genetic lineage L1 ( Figure 1 View FIGURE 1 ), which was also the most widespread and common lineage found in the study of Taheri et al. (2018a). Examination of these specimens showed that there is considerable variation in many of the setal characters (as detailed in the preceding paragraph), male reproductive characters, and to a lesser extent, in the intestine.
Gates (1973) observed similar variation but a majority of his material had the more anterior caudal zone. However we cannot know to which lineage(s) his material belonged. In our material examined, those of L2 had the caudal zone in 111–114. The L2 setae had very slight displacements, and generally more posteriorly than in L1: a lxxiv, b xxxiv, c lxviii; the last being an uncommon instance of starting alternating placement anterior to row a. However, in an L1 individual (BRSCDNA0169) rows a and c began alternating in the l to liii range.
Müller (1857) gave no internal anatomical information, other than stating that the "stomach is strongly muscular;" we interpret this as referring to the gizzard. Hence all that follows cannot be compared to the original description. A few individuals of L1 had at least one male apparatus developed: an iridescent male funnel enclosed within a testes sac, and connected to a large seminal vesicle extending posteriorly to xxviii or farther, up to xxxiv. Only one had both sides developed, whereas the others only had the left set developed in this way. Still others had the appearance of functioning male gonad(s) but no iridescence and no evident filling of the seminal vesicles. Generally the testes sacs, funnels and vesicles were atrophied or absent. The individual BRSCDNA0163 with the maximum degree of male organ development also had curious small structures attached to the outer body wall of segments iii and iv. These strongly resembled spermatophores but the placement is quite unusual and the size relatively small.
We observed that the anterior intestine is differentiated from the posterior at a constriction somewhere in the range of 22/23 to 23/24. The typhlosole always begins behind this constriction. The anterior section is consistently broader and has a very distinctive pattern of three blood vessels within the intestinal wall on each side, per segment. These are attached to short branches coming from the ventral surface of the dorsal vessel in segments xvii–xxiii, xxiv, as noted also by Righi & Bittencourt (1972). Gates (1973) makes no mention of it, despite his having been a rigorous examiner of somatic morphology. We also observed that in segments xv and xvi, there are paired slender lateral vessels, freely suspended in the body cavity, connecting the dorsal vessel to the ventral vessel, though Righi & Bittencourt (1972) showed these as connecting to the extra-esophageal vessel.
The segment of intestinal origin was usually observed to be xvii, but this was often hard to decide, and was dependent on the condition of the specimens. In some cases it appeared to be in xv or xvi, but then always doubtful because in such cases there was no sharp distinction between the intestine and the esophagus. Where that distinction was clear, the intestinal origin was always xvii.
| MH |
Naturhistorisches Museum, Basel |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Pontoscolex corethrurus
| James, Samuel W., Bartz, Marie L. C., Stanton, David W. G., Conrado, Ana C., Dupont, Lise, Taheri, Shabnam, Silva, Elodie Da, Cunha, Luís & Brown, George G. 2019 |
Lumbricus corethrurus Müller, 1857a : 113
| Muller 1857: 113 |