
Paretroplus loisellei, Sparks, John S. & Schelly, Robert C., 2011
|
publication ID |
https://doi.org/ 10.5281/zenodo.204397 |
|
DOI |
https://doi.org/10.5281/zenodo.5673938 |
|
persistent identifier |
https://treatment.plazi.org/id/1C78F415-0C26-695C-BED8-DCAACF284051 |
|
treatment provided by |
Plazi |
|
scientific name |
Paretroplus loisellei |
| status |
sp. nov. |
Paretroplus loisellei View in CoL , new species
Figures 5–7 View FIGURE 5 View FIGURE 6 View FIGURE 7 ; Table 1 View TABLE 1
Paretroplus View in CoL nov. sp. “ventitry” — Sparks and Stiassny 2003: Table 9.1. Paretroplus View in CoL sp. Mahanara River — de Rham and Nourissat 2004: 117–118. Paretroplus View in CoL nov. sp. “ventitry” — Sparks and Stiassny 2008: Table 1 View TABLE 1 . Paretroplus damii View in CoL “ventitry” — Sparks 2008.
Holotype. AMNH 251631, 142.8 mm SL; Madagascar: Antsiranana Province: Mahanara River: ca. 4 km northwest of Antsirabe-Nord: 13 57ʹ 18.0ʺ S, 49 56ʹ 12.0ʺ E; P. V. Loiselle and local fishermen, 31-X-2001.
Paratypes. AMNH 231248, 3 ex., 1 ex. C&S, 71.6–139.0 mm SL; Madagascar: Antsiranana Province: Mahanara River: main channel of Mahanara River at Antsirabe-Nord, at bridge on Route N-5: 13 38ʹ 29.4ʺ S, 49 57ʹ 48.6ʺ E; P.V. Loiselle, 9-X-2000. — AMNH 231257, 4 ex., 1 ex. C&S, 74.9–88.5 mm SL; Madagascar: Antsiranana Province: Mahanara River: main channel at Antsirabe-Nord, at bridge on route N-5: 13 58ʹ 29.4ʺ S, 49 57ʹ 48.6ʺ E; P.V. Loiselle, 13-X-2000. — AMNH 232457, 3 ex., 129.5–149.0 mm SL; Madagascar: Antsiranana Province: Mahanara River: ca. 4 km northwest of Antsirabe-Nord: 13 57ʹ 18.0ʺ S, 49 56ʹ 12.0ʺ E; P.V. Loiselle and local fishermen, 31-X-2001. — AMNH 232461, 3 ex., 96.0– 134.1 mm SL; Madagascar: Antsiranana Province: Mahanara River: at Antsirabe-Nord just upstream of bridge over route N-5: 13 58ʹ 29.4ʺ S, 49 57ʹ 48.6ʺ E; P. V. Loiselle and local fishermen, 1-XI-2001. — AMNH 236145, 112.5 mm SL; Madagascar: Antsiranana Province: Mahanara River, main channel at Antsirabe-Nord; P.V. Loiselle. — AMNH 251632, 128.5 mm SL; Madagascar: Antsiranana Province: Mahanara River, main channel at Antsirabe-Nord: 13 58ʹ 51ʺ S, 49 57ʹ 18ʺ E; P.V. Loiselle, 18-X-2007. — UMMZ 249229, 107.1 mm SL; data as for AMNH 231248.
Diagnosis. A species of Paretroplus distinguished from all congeners except its sister taxon, P. d a m i i, by the presence of a triangular, black pectoral-axil patch in combination with the absence of a series of prominent vertical bars on the flank. In life, unstressed individuals of these two species can easily be distinguished from all congeners by the presence of a broad, vertical brick red or pale yellow band on the anterior half of the flank. Paretroplus loisellei is distinguished from P. damii by the presence of a more or less straight gular region in lateral view (vs. rounded and convex in P. damii ), such that the ventral margins of the preopercle and interopercle are essentially horizontal. In P. loisellei the oral jaws are horizontally oriented (vs. strongly oblique in P. d a m i i), the lips are enlarged and fleshy (vs. thin in P. d a m i i), and the snout is elongate and curved, creating a beaked appearance (vs. short and straight in P. d a m i i). The enlarged symphyseal teeth in the upper jaw possess distinct, sharp lateral cusps (vs. poorly developed and rounded in P. damii ). Owing to convex premaxillary pedicels, Paretroplus loisellei exhibits a prominent indentation marking the transition from the dorsal margin of the premaxillary ascending process to the supraoccipital crest. The base coloration in P. loisellei is uniform dark brownish-gray in preservation; whereas P. d a m i i is considerably lighter golden brown overall and frequently retains a faint, pale broad band on the anterior region of the flank. Paretroplus loisellei exhibits a distinctive bright golden breeding coloration (vs. brick red in P. d a m i i).
Description. Morphometric and meristic data presented in Table 1 View TABLE 1 . External morphological characteristics and general pigmentation pattern in life and preservative can be observed in Figures 5 View FIGURE 5 and 6 View FIGURE 6 . A relatively shallow bodied Paretroplus belonging to the “ Paretroplus damii clade” of Sparks (2008: Clade A, Fig. 1 View FIGURE 1 ), which also includes P. damii , P. nourissati , P. tsimoly , and P. lamenabe . Snout pronounced and beak-like owing to elongate and convex premaxillary ascending process. Head pointed, snout curved/convex, and predorsal profile moderately to strongly curved/convex. Prominent indentation present just anterior to orbit, owing to curved premaxillary pedicels. Dorsal body outline slightly curved, and ventral outline mostly straight, except posteriorly. Caudal peduncle short, deep, and strongly laterally compressed. No sexually dimorphic features apparent in material examined.
Total vertebral count 31 or 32, with formulae of: 15 + 16, 15 + 17 (mode), and 16 + 16 precaudal and caudal vertebrae, respectively.
Jaws isognathous and horizontally oriented. Lips of both upper and lower jaws enlarged and fleshy. Single row of anteroposteriorly flattened, spatulate, unicuspid teeth in both upper and lower jaws. Premaxillary symphyseal teeth greatly enlarged with small, yet distinct and sharp lateral cusps (i.e., weakly tricuspid); other upper jaw teeth graded in size and becoming progressively much smaller posterolaterally ( Fig. 7 View FIGURE 7 A). Lower-jaw teeth at symphysis reduced in size compared to adjacent lateral teeth, presumably to accommodate enlarged upper symphyseal teeth when jaws closed. Teeth flattened at crown and procumbently implanted. Teeth in upper jaw number six or seven on each side, and total 12 to 13. Teeth in lower jaw number six or seven on each side, and total 12 to 13. Teeth in both upper and lower jaws in general irregularly spaced, particularly posteriorly in both jaws.
Upper and lower pharyngeal tooth plates well developed and robust. Dentition on lower pharyngeal tooth plates [= lower pharyngeal jaws (LPJ) or fifth ceratobranchial tooth plates] hooked and bicuspid both laterally and anteriorly, becoming progressively enlarged medially. Molariform teeth present only posteromedially on LPJ. LPJ well sutured, with numerous interdigitating sutures on posteroventral margin. Seven to nine robust tooth plates cover majority of dorsal surface of fourth ceratobranchial bones. Fourth ceratobranchial tooth plates not confluent with outer-row gill rakers of these elements. Bases of individual tooth plates exhibit varying degree of fusion depending on the individual. Dentition on fourth ceratobranchial tooth plates unicuspid or weakly hooked and bicuspid laterally, hooked and bicuspid medially (similar to lateral LPJ dentition). Dentition on third upper pharyngobranchial tooth plates molariform only posteromedially, hooked and bicuspid laterally and anteromedially. Dentition on second pharyngobranchial tooth plates hooked and bicuspid, and arrayed in three (rarely four) rows.
Character N Holotype Range Mean SD Standard length (mm) 15 142.8 85.9–142.8 117.5
Caudal peduncle length/width 15 2.5 1.6–2.7 2.2 0.31 Caudal peduncle length/depth 15 0.5 0.3–0.5 0.4 0.05 Scales in lateral line 15 38 (8) (H), 39 (7)
Gill rakers (lower limb 1st arch) 14 12 (5), 13 (9) (H)
Vertebrae (precaudal + caudal) 15 15 + 16 = 31 (2), 15 + 17 = 32 (12) (H), 16 + 16 = 32 (1) Dorsal fin 15 XVIII 12 (1) (H), XIX 12 (7), XIX 13 (2), XX 11 (1), XX 12 (3), XX 13 (1) Anal fin 15 VIII 10 (1), IX 9 (1) (H), IX 10 (7), IX 11 (3), X 9 (3)
Twelve or 13 (mode) triangular and moderately elongate gill rakers arrayed along lower limb of first gill arch. Rakers of first arch weakly denticulate dorsomedially. All other lower-limb rakers (i.e., those on gill arches 2 through 4) short, triangular in shape (not spherical), and strongly denticulate dorsally. Epibranchial rakers elongate, numbering 11 to 12.
Body covered with large, regularly imbricate, cycloid scales. Caudal margin (= posterior field) of flank scales thin, unossified and lacking circuli, creating a thin, flexible “flap”. Well-developed ridges of scales (= scale sheathing, Sparks 2008) present along dorsal- and anal-fin bases. Scale ridges free from spiny dorsal and anal fins, but becoming weakly attached to both soft dorsal and anal fins, such that three or four rows of smaller scales present caudally on both soft dorsal and anal fins. Numerous rows of smaller scales extend onto caudal fin, posterior of hypural flexure, for approximately 1/3 of its length. Pelvic axillary scale well developed and elongate. Lateral-line scales number 38 (mode) or 39. Chest scales reduced in size and embedded, with scales along ventrum extremely reduced in size. Four to five rows of scales on cheek. Opercle, subopercle, and interopercle fully scaled. Snout, lacrimal, and rostral portion of interorbital region asquamate. Preopercle asquamate ventrally, scaled only on anterodorsal portion of shaft where posteriormost cheek scales overlap.
Dorsal fin with XVIII–XX spines and 11–13 soft rays. Anal fin with VIII–X spines and 9–11 soft rays. Origin of dorsal fin at level of vertical through pectoral-fin insertion at its dorsal insertion point. Caudal fin weakly emarginate. Upper and lower lobes quite broad and rounded. Pectoral fin large, broad, and rounded at distal margin. Distal margins of soft dorsal and anal fins becoming increasingly pointed in larger specimens. Distal margins of both fins extend well past caudal origin. Pelvic fin extending to about level of anal-fin origin or just beyond origin.
Miscellaneous osteology and gas bladder anatomy. Members of Paretroplus damii clade (Clade A) exhibit most structurally complex anterior gas bladder diverticula and associated bullae within Etroplinae (Sparks 2008: Fig. 9 View FIGURE 9 B). Expansive and complex exoccipital foramina present, with posterolateral chamber. Paired anterior gas bladder bullae with tough and thickened tunica externa, anteriormost chambers firmly lodged in exoccipital recesses that comprise multiple internal chambers. Excavation (= supraoccipital notch of Stiassny et al. 2001) along posterior margin of supraoccipital generally poorly developed (shallow or narrow) or small and rounded. Supraoccipital extending anteriorly over median frontal pores of neurocranium (NLF0 of Barel et al. 1977). Lacrimal comprising two “plates” - lacrimal proper (= anterior plate; IO1) and plate-like “primitive second infraorbital” (= IO2) of Cichocki (1976) ( Fig. 7 View FIGURE 7 B). Anterior lacrimal plate anteroventrally expanded and paddle-shaped, with elongate lateral line canals bearing constricted foramina ( Figs. 5 View FIGURE 5 & 7 View FIGURE 7 B). Shared marginal canal between lacrimal plates, IO1 and IO2, greatly reduced or absent.
Coloration in life. Base body coloration golden brown, grayish, or dark grayish-black (de Rham & Nourissat 2004: 114–117). Triangular black patch present in pectoral-fin axil and extending dorsal to pectoral-fin base. Unpaired fins olive-grey to grayish-black. Pectoral fin brown to grayish-black. Pelvic fin gray, grayish-black, or black. Breeding coloration bright golden yellow (see color image in de Rham & Nourissat 2004), particularly ventral to dorsal midline, not brick red as in P. d a m i i (P. Loiselle, pers. comm.).
Coloration in preservative ( Figures 5 View FIGURE 5 & 6 View FIGURE 6 ). Dark grayish brown to dark rusty brown; body somewhat darker dorsally, particularly along base of dorsal fin and dorsal portion of head. Triangular black patch characteristic of clade present in pectoral-fin axil and extending dorsal to pectoral-fin base ( Fig. 5 View FIGURE 5 ). Lips pale yellow to light olive, particularly ventrally. Dorsal margin and posterior portion of upper lip dark gray to grayish-black. Unpaired fins gray to grayish-black. Pectoral fin pale yellow to olive. Pelvic fin grayish-black to black.
Distribution and habitat. The new species has only been collected from the middle reaches of the eastward draining Mahanara River, located to the north of Sambava near the town of Antsirabe-Nord, northeastern Madagascar ( Fig. 8 View FIGURE 8 , black circle). Local inhabitants report the occurrence of P. loisellei in adjacent basins; however, no specimens have been collected outside of the Mahanara River to date (P. Loiselle, pers. comm.)
Local name. Ve n ti tr y.
Etymology. Named in honor of Dr. Paul V. Loiselle, Emeritus Curator of Freshwater Fishes at the New York Aquarium (WCS), for directing our attention to this new taxon, and in appreciation of his efforts to document, preserve, and educate the public regarding Madagascar’s unique and severely threatened freshwater ichthyofauna. Relationships and discussion. Clade D, including P. loisellei , new species, and P. d a m i i is recovered as the sister taxon to a clade comprising P. t s i m o l y, P. nourissati , and P. lamenabe ( Fig. 1 View FIGURE 1 ): Clade N. Members of the latter clade are readily distinguished from the former two species by the presence of two prominent and converging midlateral bands (several fainter bands are also present on the flank) and elongate nasal bones, features that are lacking in P. loisellei and P. d a m i i (compare Figs. 2 View FIGURE 2 & 3 View FIGURE 3 and 5 View FIGURE 5 & 6 View FIGURE 6 , with Fig. 4 View FIGURE 4 ). Members of the “ Paretroplus damii clade” of Sparks (2008), comprising P. d a m i i, P. tsimoly , P. nourissati , P. lamenabe , and P. loisellei , new species, are united by the shared presence of five unambiguously optimized apomorphic morphological features [See Diagnosis for “Clade D: Paretroplus damii species group” above, and Sparks (2008)].
Interestingly, our phylogenetic analysis revealed the existence of yet another novel taxon within this clade, referred to herein as P. n. sp. “anjingo” ( Fig. 9 View FIGURE 9 ), which is endemic to the Anjingo and Ankofia river drainage basins, including Lac Andrapongy, in northwestern Madagascar ( Fig. 8 View FIGURE 8 ). Paretroplus n. sp. “anjingo” is recovered as the sister taxon to the clade comprising P. d a m i i and P. loisellei , with which it shares the four apomorphic features that distinguish this group from Clade N: absence of a darkly pigmented vertical barring pattern; a characteristically uniform brown (grayish-, reddish-, or golden-brown) overall coloration as adults (unstressed individuals may exhibit a broad golden yellow to reddish band on the anterior flank); absence of broad, converging black midlateral bars; small lateral cusps on the two enlarged symphyseal teeth in the upper jaw, such that these teeth are weakly tricuspid. As noted by Sparks (2008) and communicated by P. Loiselle (pers. comm.), specimens historically assigned to P. damii ( Fig. 2 View FIGURE 2 ), as well as populations of morphologically similar fishes recently discovered in extreme northern and northeastern Madagascar ( Figs. 3 View FIGURE 3 , 5 View FIGURE 5 , 6 View FIGURE 6 & 9 View FIGURE 9 ), were hypothesized to represent possibly as many as three distinct species, a result corroborated by our phylogenetic analysis ( Fig. 1 View FIGURE 1 ).
It is also worth noting that an identical biogeographic pattern is recovered for the endemic Malagasy cichlid genus Ptychochromis , with distinct species exhibiting the same geographic ranges as Paretroplus damii (viz. Ptychochromis oligacanthus ), P. loisellei (Pty. loisellei ), and P. n. sp. “anjingo” (Pty. inornatus ) ( Sparks 2002; Stiassny & Sparks 2006: Fig. 1 View FIGURE 1 ). The Anjingo/Ankofia drainage basin is characterized by a highly endemic ichthyofauna ( Sparks 2005), and is also home to an endemic aplocheiloid killifish ( Pachypanchax sparksorum ). As for Paretroplus and Ptychochromis , a distinct killifish species ( Pachypanchax omalonotus ) occurs to the north of the Anjingo/Ankofia basin in the Sambirano River and on the satellite island of Nosy Be ( Loiselle 2006) ( Fig. 7 View FIGURE 7 ). The biogeographic disjunction between the Anjingo/Ankofia River system and the Sambirano River directly to the north is intriguing and worth investigating for other aquatic, as well as terrestrial, taxa.
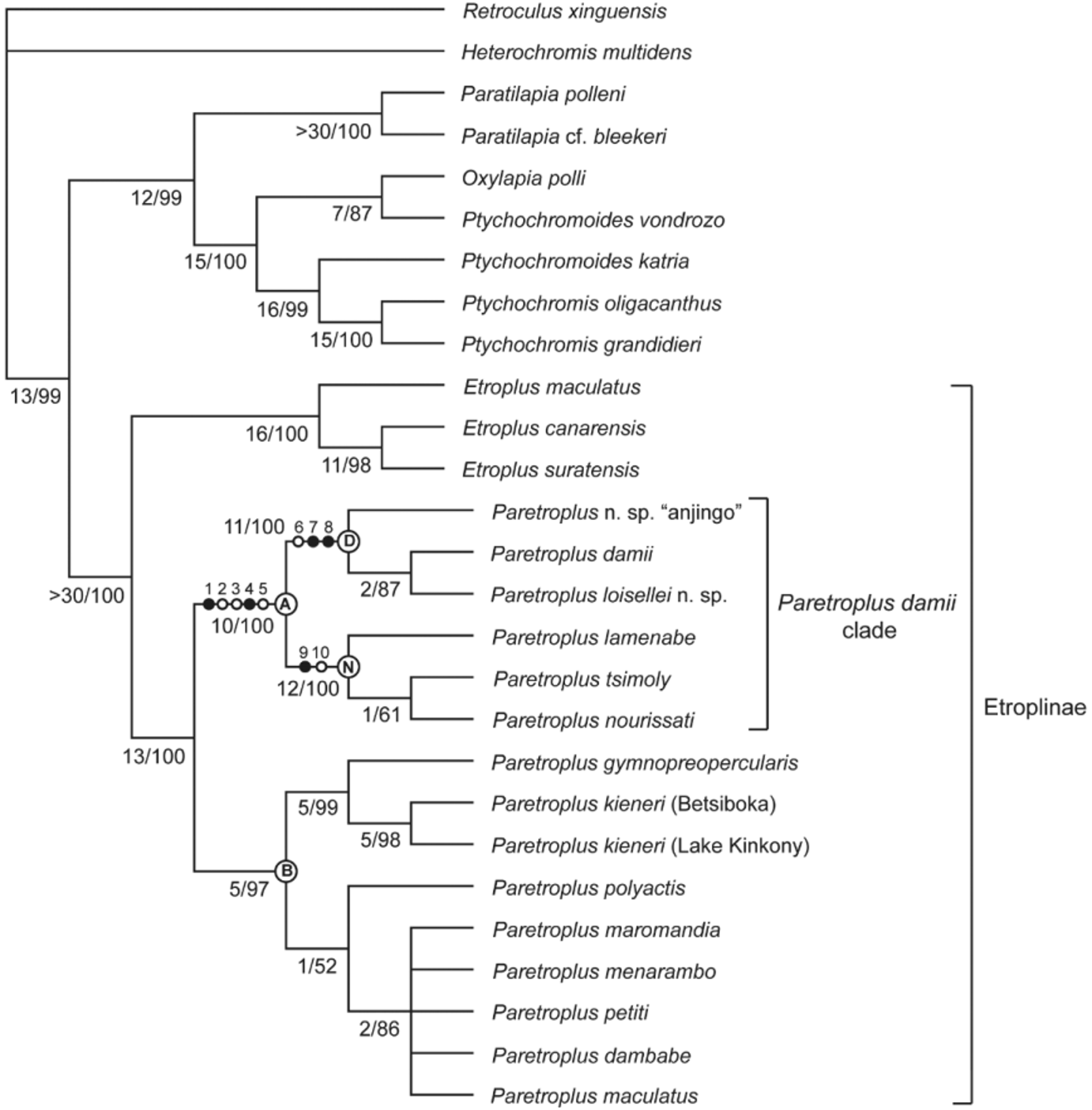





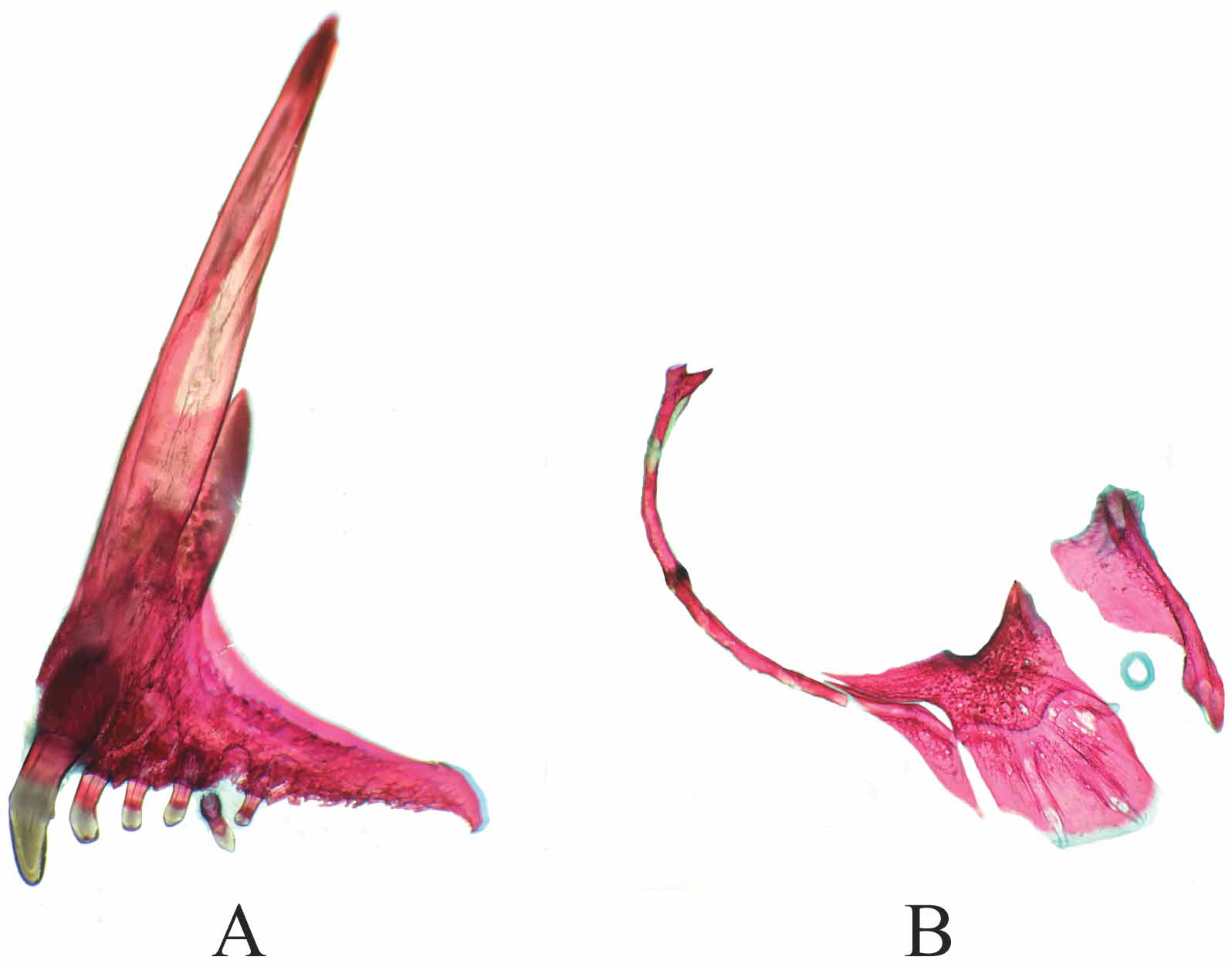
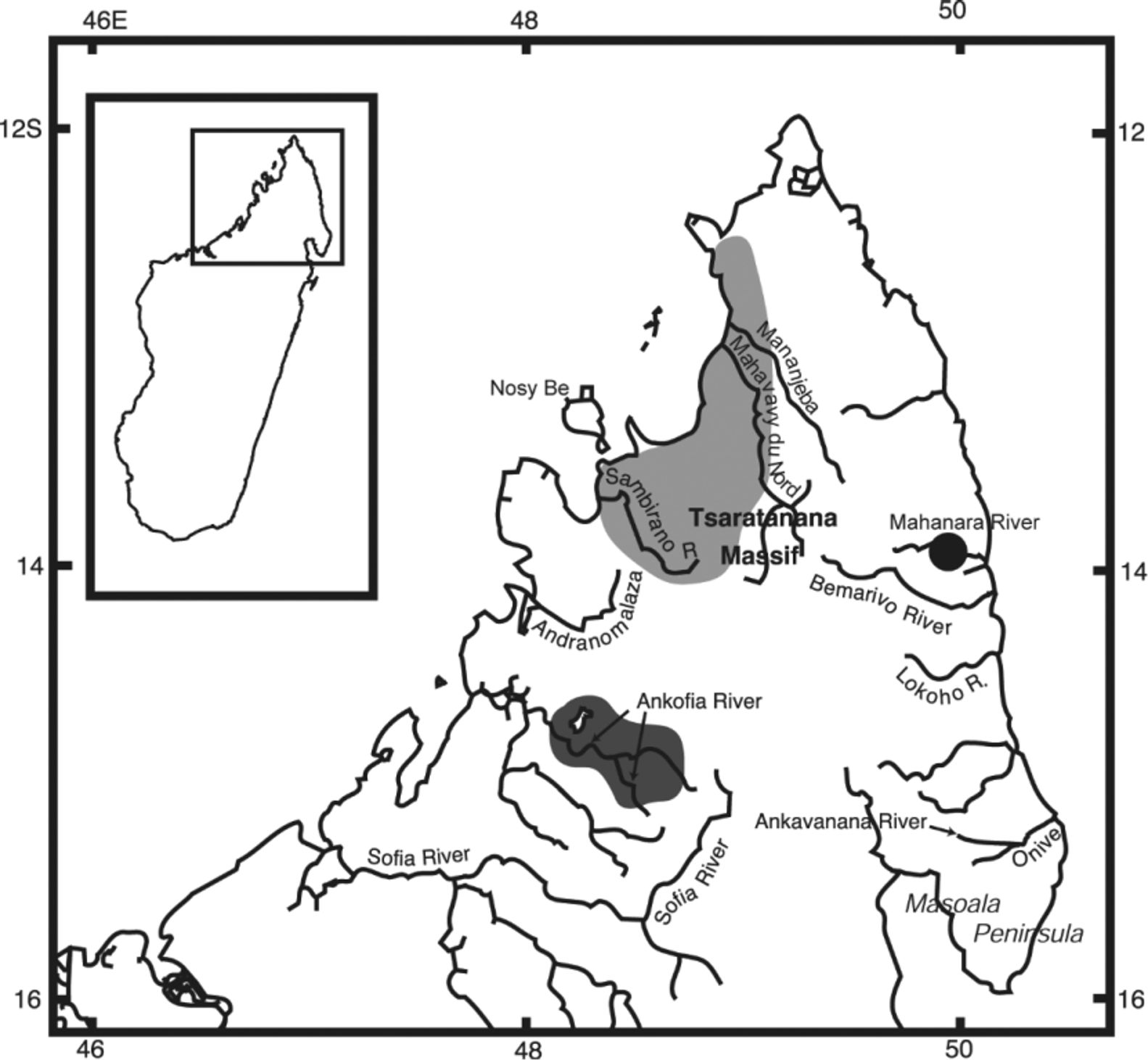

TABLE 1. Morphometric and meristic data for Paretroplus loisellei. For meristics, numerals in parentheses indicate number of specimens examined with that count, and (H) indicates count corresponding to holotype.
| Percentage of SL | |||||
|---|---|---|---|---|---|
| Head width (max.) | 14 | 17.2 | 17.0–18.6 | 17.6 | 0.44 |
| Head length | 15 | 31.5 | 31.5–34.5 | 32.5 | 0.84 |
| Body depth | 15 | 48.7 | 46.9–52.6 | 49.5 | 1.38 |
| Caudal peduncle length | 15 | 8.2 | 5.3–8.9 | 7.2 | 0.90 |
| Caudal peduncle width | 15 | 2.6 | 2.5–4.5 | 3.3 | 0.55 |
| Caudal peduncle depth | 15 | 16.0 | 14.9–17.2 | 16.0 | 0.61 |
| Pectoral–fin length | 15 | 20.7 | 19.3–22.6 | 20.8 | 1.02 |
| Pelvic–fin length | 15 | 20.2 | 19.4–21.7 | 20.7 | 0.72 |
| Pre–dorsal length | 15 | 47.1 | 45.0–52.3 | 46.9 | 1.77 |
| Pre–anal length | 15 | 59.4 | 59.0–62.3 | 60.3 | 1.14 |
| Pre–pelvic length | 15 | 40.1 | 40.1–44.2 | 41.8 | 1.11 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |