
Ichthyscopus pollicaris, Vilasri & Ho & Kawai & Gomon, 2019
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4702.1.10 |
|
publication LSID |
lsid:zoobank.org:pub:42D46A1B-1827-4971-9CF6-F46F291969A0 |
|
persistent identifier |
https://treatment.plazi.org/id/1B49E033-FB25-5863-FEE8-9F01AFEA480F |
|
treatment provided by |
Plazi |
|
scientific name |
Ichthyscopus pollicaris |
| status |
sp. nov. |
Ichthyscopus pollicaris sp. nov.
English name: Oriental Fringe Stargazer; Japanese name: ṷỵtìÿ̚ (Satsuo-mishima); Taiwan name: AEḧā üḃ (Dong-fang-pi-jian-teng)
Figs. 1 View FIGURE 1 A–B, 2A–D, 3A, 4A, 5A; Table 1 View TABLE 1
Ichthyscopus lebeck View in CoL (non Bloch & Schneider, 1801): Chen, 1969: 430; Shen, 1984a: 116, fig. 378-2; Jordan & Snyder, 1901:
109; Jordan et al., 1913: 369; Ui, 1924: 257; Okada & Matsubara, 1938: 384; Kamohara, 1950: 261; Matsubara, 1955: 707;
Shen, 1984b: 378; Chen & Yu, 1986: 701, fig.6-148; Shen, 1993: 481, fig.162-4; Chen, 2003: 181; Chen, 2004: 126. Ichthyscopus lebeck lebeck (non Bloch & Schneider, 1801): Shen & Wu, 2011: 610. Ichthyscopus lebeck sannio (non Whitley, 1936): Mees, 1960: 52–53 (in part); Kishimoto, 1984: 293, pl. 263-B; Kishimoto,
1997: 557; Yamada, 2002: 1076. Ichthyscopus sp.: Gomon & Johnson, 1999: 616 (in part). Ichthyscopus sannio (non Whitley, 1936): Yamada & Yagishita, 2013: 1279.
Holotype. NMMB-P24460 (187 mm SL) Daxi , Yilan, northeastern Taiwan, western Pacific Ocean, 22 Mar. 2015.
Paratypes. HUMZ 221796 View Materials (239 mm SL) Dong-gang Fishing Port, Taiwan ; HUMZ 222077 View Materials (161 mm SL) Nanfang-ao Fishing Port, Taiwan ; NMMB-P6314 (161 mm SL), Kaohsiung, Taiwan, 16 June 1960 ; NMMB-P6363 (223 mm SL) Chin-quen-son, Tainan, Taiwan, June 1963 ; NMMB-P31287 (ex. NMMB-P24460 , 2, 211–212 mm SL) Daxi , Yilan, northeastern Taiwan, 22 Mar. 2015 ; NMMB-P25632 (171 mm SL) Ke-tzu-liao, Kaohsiung, Taiwan, 7 Dec. 2016 ; NMMB-P28076 (308 mm SL) Ke-tzu-liao, Kaohsiung, southwestern Taiwan, 18 Jan. 2018 ; NMV A31864 View Materials -001 About NMV (216 mm SL) and KAUM–I. 128242 (230 mm SL) Nan-fang-ao, Yilan, NE Taiwan, NW Pacific, 5 Apr. 2018 ; NSMT-P 34566 (169 mm SL) Sagami Bay , off Oiso, Kanagawa Prefecture, Japan, 25 Mar. 1991 .
Diagnosis. A species of Ichthyscopus with caudal vertebrae 17; upper lip fimbriae 20–27; ventral midline of belly forming three prominent prolonged skin flaps, one midline flap and two sandwiching midline flap; shape of posterior nostril oval to circular; pectoral-fin base uniformly pale brown, lacking distinct blotches; 1 or 2 white spots present on uppermost pectoral-fin ray.
Description. Morphometric and meristic data are presented in Table 1 View TABLE 1 . Description is based on the holotype followed by the paratypes in parentheses when variation present.
Body elongated, moderately deep and compressed posteriorly. Head large and slightly compressed, mostly encased in armoured bones; bony surface sculptured with fine low vermiculations; dorsal surface slightly convex. Interorbital fossa rounded posteriorly, width posteriorly rather less than between orbits. Orbit slightly oval with diameter distinctly greater than that of eyes. Nostrils with margin edged by bushy branched fimbriae; anterior nostril small, short, circular and tubular; posterior nostril small oval to circular, confined posteriorly in groove at anterior corner of orbit ( Fig. 3A View FIGURE 3 ). Mouth large, opening distinctly superior; both lips with numerous long, compressed and brush-like fimbriae externally; lateral surface of lower jaw forming concave skin flap that covers ventrolateral rim of lower lip. Teeth on prevomer and palatine numerous, minute and close-set in oval and elongate patches respec- tively; teeth on dentary conical caniniform, uniserial and distinctly separated from each other; teeth on premaxilla small, caniniform, numerous and close-set. Respiratory valve inside lower jaw broad, fleshy, with short appendage medially. Chin smooth, lacking barbel. Opercular bones enlarged; dorsal half of opercular membrane edged by flaplike fimbriae with several arborescent branches; ventral margin of membrane almost smooth, but weakly crenulated ventrally ( Fig. 4A View FIGURE 4 ).
*Data from Gomon & Johnson (1999) including AMS IA6309 (holotype) and 23 non-type specimens.
Cleithral spine short, fully enclosed in flattened-fleshy skin flap; length of spine1.8 (1.6–2.0) in length of skin flap. Ventral margin of cleithral flap fimbriate with 22 (17–22) arborescent-branched fimbriae, extending from anterior edge inside of opercular opening to posterior tip of flap. Narrow fleshy groove developed between anteroventral margin of cleithral flap and dorsal margin of pectoral base; ventral border of groove forming a fimbriate ridge from inside of opercular opening to about 1/3 length of uppermost pectoral ray, with widely separated tiny fimbriae anteriorly, that gradually become larger and arborescently branched posteriorly. No spines associated with subopercle, opercle, preopercle or pelvic girdle exposed from skin.
Dorsal fin rather long-based and low, with anterior and posterior segments continuous; first two spines relatively short with subsequent soft rays becoming progressively longer. Anal fin long-based and low. Pectoral fin knifeshaped and rounded posteriorly with 5th ray longest, membrane incised slightly between tips of rays. Pelvic fins large and fleshy; pelvic spine reduced, embedded under skin, following branched rays long, innermost ray longest; interradial membranes obviously incised between adjacent branched rays distally. Caudal fin slightly rounded with tips of branched caudal rays extending slightly beyond membrane. Ventromesial surfaces of throat and belly with three prominent skin flaps, comprising long medial flap and two shorter lateral flaps sandwiching medial flap; medial flap prominent and large, extending from anterior tip of basipterygium to rear of pelvic-fin base, subsequently becoming inconspicuous and low on anterior half of belly, flap separated (continuous) on middle of belly, again becoming gradually prominent and deep posteriorly to anus; lateral flaps developed on posterior half of belly with posterior end approaching middle of second anal-fin ray, anterior portion of flaps low and thin but becoming deeper and forming blade-like flap posteriorly ( Fig. 4A View FIGURE 4 ).
Scales small, cycloid, rectangular, arranged in oblique rows directed downward and backward on side of body, but sometimes several posterior rows directed downward and forward dorsally; occasionally two dorsal rows fused into one and continuing ventrally; additional 3 (3 or 4) rows of small scales medially on upper half of caudal base. Ventral side of body naked. Lateral line embedded in skin with several angled rows of pores, short and narrow rows of pores emerging with 1 or 2 pores at dorsal end of each oblique scale row on side of body ventrally and ventral end of those on nape dorsally; irregularly enlarged and plate-like rows becoming wedge shaped except largest lowermost row oval shaped, and emerging numerous pores at base of caudal rays in all upper and 2 middle of lower elements; lateral line running from posterior surface of posttemporal to middle of caudal fin, anteriorly rising to just beneath base of 3rd dorsal-fin ray and running horizontally adjacent to dorsal-fin base, to dorsal midline of caudalfin base where those of the two sides fuse, posteriorly curving downward and terminating at middle of caudal-fin base. Nape between lateral lines mostly naked with up to 9 very short, oblique scale rows directed downward and backward above lateral line (NMMB-25632 variously covered by fully developed scales).
Color when fresh ( Figs. 1A View FIGURE 1 , 2 View FIGURE 2 A–D). Upper half of body dark brown. Blotches dorsally on head and nape pale brown surrounded by dark brown ring; those on trunk, cleithral skin flap, dorsal fin, uppermost pectoral ray and caudal fin vivid creamy white. Lower half of body mostly pale brown, except base of pectoral fin, belly and area above anterior 5 (4–6) anal fin rays pale white. Exposed opercular bones flesh pink, with irregular pale brown pattern. Pectoral fin brown with lower portion yellow. Pelvic fin yellow (yellow to olive brown). Anal fin yellow. Caudal fin brown basally and becoming yellow distally.
Color in alcohol ( Fig. 1B View FIGURE 1 ). Dark brown on upper half of head and body. Upper half of body marked with about three longitudinal series of circular, oval to irregular conspicuous large blotches, small anterodorsally and becoming larger posteriorly and ventrally; nape between lateral lines, top of head, cheek, lips and chin scattered with oval to irregular pale blotches surrounded by dark ring, less conspicuous on exposed opercular bones; blotches of somewhat moderate size posteriorly and becoming smaller anteriorly, those on chin smallest; blotches on nape and cheek conspicuous (fading or absent in larger specimens); no dark or dusky dots scattered on these areas. Lower half of head and body pale, with minute scattered melanophores. Cleithral skin flap brown, occasionally with pale rim ventrally; 2 (1 or 2) moderately pale blotches without dark ring between central and posterior portions of flap. Dorsal fin dark brown with moderately sized pale blotches forming 2 irregular longitudinal stripes, dorsal spines and adjacent interspace membrane dark, some with 1 (1 or 2) small pale blotches. Anal fin pale. Caudal fin brown usually with 5 (1–5) pale irregular blotches or longitudinal bars on basally, centrally and distally upper half of fin, some fused with corresponding marking on opposite. Pectoral fin uniformly brown with base lacking distinct blotches; uppermost ray and adjacent intervening membrane with 2 (1 or 2) white spots (NMV A31864 View Materials -001 variously with additional 2 white spots on 2nd and 4th pectoral fin rays, respectively); ray tips and incised membrane distinctly pale ( Fig. 5A View FIGURE 5 ). Pelvic fin pale.
Distribution. Restricted to the coastal waters of East Asia between about Japan Sea and East China Sea from Jeju Province in South Korea through Niigata Prefecture in Japan to the western coast of Kyushu and Pacific Ocean from Suruga Bay to the southern coast of Kyushu ( Yamada & Yagishita, 2013) in the north and the northeast region of South China Sea from the northern Taiwan ( Shen, 1984a) through Penghu Islands ( Chen, 2003) to the southern Taiwan (present study) in the south.
Etymology. The specific name pollicaris from the Latin meaning “thumb” in reference to the distinctive uppermost pectoral fin ray adorned with 1 or 2 white spots characterizing this species.
Remarks. Ichthyscopus pollicaris is similar to I. lebeck and I. sannio among its congeners in having a prominent medial skin flap on ventral midline extending from the anterior tip of the basipterygium to the rear end of the pelvic-fin base, and continuing posteriorly to the anus ( Fig. 4 View FIGURE 4 A–B); and the dorsal side of body dark brown with several distinctly pale blotches, without distinct transverse bands (vs. a ventral midline with a prominent medial skin flap extending only from the anterior tip of the basipterygium to the rear end of the pelvic-fin base; and with dark transverse bands, saddles or spots present in the other 5 species of Ichthyscopus in Gomon & Johnson, 1999 ).
Ichthyscopus lebeck was originally described from Tranquebar in India, but the status of its type is currently unknown. The Latin description in Bloch & Schneider (1801) is very informative in determining the possible identity of this species, which can be translated as follows: Branchiostegal membrane 5 rays, opercula double layered under the chest 1.5 soft protrusions (pollicis in Latin means thumbs that may indicate protrusion of these elements) broadly spread out, double lobed, split in the center but joined; the other lower unarmed opercula end in soft lobes; towards the opposite upper side are broad protrusion with a moss-like fringe. Posterior to the operculum and above the pelvic fins has a semilunar protrusion. The arch of the upper aperture of the mouth bears 2 broad protrusions. The lips fleshy, transversely striated, with fringed edges. Nostrils between the eyes and the lips; between them a triangular region with mossy fringe ascends above the eyes (may mean the posterior nostril extending the interorbital fossa above the orbit). Tongue cartilaginous, soft, immobile; teeth of the lower jaw short, sharp, gradually lengthening posteriorly; teeth of the upper jaw resemble a rough file. Caudal fin rounded; dorsal and anal fins elongated; rays of all fins soft. Eyes small, greenish, prominent, covered by nictitating membrane, black pupil enclosed by a golden circle, no vestige of a lateral line. Above the paired blades of the upper jaw there opens a soft oval body in the shape of a bladder. The exposed belly surrounds 3 length protrusions (may mean the three prominent skin flaps on belly). Color of the back reddish becoming green, a pair of white spots in a row manifest; similarly spots on the pectoral fin; nape of neck unspotted, glossy; crown bony, rough resembling written Arabic characters; color of the belly and fins citrine, becoming white caudally.
The comparative specimens of the present study collected from Ranong Province of Thailand and Singapore agree well with the description in having the posterior nostril positioned in the interorbital fossa above the orbit, the three prominent skin flaps on the belly and body coloration ( Fig. 6 View FIGURE 6 ). We conclude they are I. lebeck .
Ichthyscopus sannio was originally described by Whitley (1936) based on the holotype (AMS LA6309) collected from Patonga, Broken Bay, New South Wales. He distinguished I. sannio from I. lebeck by the posterior nostril (the preocular fringes “probably mean the posterior nostril” restricted to the anterior part of the interorbital depression only, vs. the fringes extending backwards half-way along each side of the depression), the granulation of the opercular bones and the vertex (less granulated, vs. much more granulated) and variations of fin rays and teeth. The identity of this species is clear from Gomon & Johnson’s (1999) redescription that characterizes it by the dorsal fin with a shallow concavity preceding soft rays, the posterior nostril oval to circular situated in the anterior gap in bony orbit, the belly with a low blade-like skin flap along ventral midline from anus to midbase of pelvic fins, the upper body with 2 poorly defined broad dark brown saddles positioned below dorsal fin anteriorly and posteriorly and with numerous irregular large white spots and blotches, and the upper part of head with many small creamy white spots.
Although sharing mostly similar in morphometric and meristic ranges as shown in Table 1 View TABLE 1 , the number of the caudal vertebrae is 17 in I. pollicaris and 16–17 in I. lebeck , but 18 in I. sannio . The number of the upper lip fimbriae is relatively greater in I. sannio and I. lebeck , 27–36 and 25–34, respectively, whereas it is least in I. pollicaris , 20–27.
The posterior nostril in I. pollicaris and I. sannio has caused confusion ( Mees, 1960), because they both have a small circular posterior nostril set in a groove at the anterior corner of the orbit ( Figs. 3 View FIGURE 3 A–B). In contrast, I. lebeck have a posterior nostril that is distinctly produced posteromesially in the interorbital fossa along the mesial side of the exposed frontal, extending beyond the center of the orbit ( Fig. 3C View FIGURE 3 ).
However, I. pollicaris is more similar to I. lebeck than to I. sannio in sharing with it a ventral midline of the body having two lateral skin flaps sandwiching the prominent medial skin flap that are well developed on the posterior half of the belly, their posterior ends attached to the middle of the second anal fin ray (vs. the two lateral skin flaps reduced and just restricted to the anterior portion from the anus to the base of the second anal fin ray in I. sannio )( Figs. 4 View FIGURE 4 A–B), no broad dark saddles across the upper body (vs. 2 poorly defined broad dark brown saddles across the upper body), an indistinct notch barely separating the anterior portion of the dorsal fin with the fin almost uniformly pale anteriorly and posteriorly (vs. a distinct shallow notch defining a somewhat separate anterior section of the dorsal fin that is noticeably darker than the fin posteriorly) and crenulations or tab-like cirri on the ventral opercular margin anterior to the pelvic fin bases (vs. a smooth edge on the opercular margin)( Figs. 4 View FIGURE 4 A–B).Although coloration may vary in these species, some details are consistent. Ichthyscopus lebeck is distinguishable from I. pollicaris in having a distinct transverse pale blotch on the pectoral base (vs. lacking a blotch) and the uppermost pectoral ray uniformly dusky (vs. the uppermost ray with 1–2 white small spots)( Figs. 5 View FIGURE 5 A–B).
Discussion. Gomon & Johnson (1999) proposed a hypothesis for the evolutionary relationships and biogreographical distributions of recognized species of Ichthyscopus and their ancestral populations, although at that time the northern hemisphere species (i.e., I. lebeck and I. pollicaris ) were not included in their analysis. However, based on a superficial examination of Indian Ocean and Northwest Pacific specimens, they hypothesized that northern hemisphere species likely constitute a monophyletic line that diverged after the isolation of I. insperatus , but prior to the divergence of I. sannio . This assumption was supported by the absence of any detectable development of a separate anterior segment of the dorsal fin in I. insperatus and the northern hemisphere species (vs. presence of an anterior segment in I. sannio and subsequently diverging I. barbatus , I. spinosus and I. nigripinnis ) and the presence of a unique crenulated margin on the opercular membrane anterior to the pelvic fin bases in only the northern hemisphere species (vs. crenulations absent in Australian species). Notably, the specialized medial skin flap that is well developed from the anterior tip of the basipterygium to the anus was not included in that analysis, because it may have been considered an autapomorphy in I. sannio . As observed above, the character is present in I. sannio and northern hemisphere species, but absent in other Australian species ( Gomon & Johnson, 1999). In additional, the presence of two lateral skin flaps sandwiching the medial skin flap that are well developed on the posterior half of the belly in the northern hemisphere species (vs. flaps reduced in I. sannio and other Australian species) supports the monophyly of the northern hemisphere species and a sister relationship with the I sannio Australian clade. The longitudinally elongate posterior nostril in I. lebeck is likely to be the even more specialized form that was derived from the primitive circular nostril in I. pollicaris and all Australasian species.
The genus Ichthyscopus is considered to be a highly evolved group of uranoscopids adapt to live in sand as ambush predators aided by several unique, adaptive characters ( Pietsch, 1989; Vilasri, 2013). Some of these like the presence of a palatine concavity contiguous with the nasal and oral cavities, fimbriae on the nostrils large and well developed, fimbriae on the lips long and compressed, fimbriae on the edges of the dorsal half of the opercular membrane and the ventral margin of the cleithral flap imply that species of Ichthyscopus specifically evolved to live in nearshore waters impacted by high concentrations of suspended sediment. This has been reinforced by reports of their distributions ( Gomon & Johnson, 1999; Bray & Hoese, 2006; Gomon, 2008; some collection data of speci- mens examined in the present study). As a consequence, paleoenvironmental changes in coastal areas could conceivably influence speciation of Ichthyscopus historically. This supports Gomon & Johnson’s (1999) hypothesis that temperature (i.e., warm or cold temperature waters) was at least in part a factor affecting the extent of distribution around the Australian continental coastline during climatic fluctuations that occurred at approximately 100,000 year intervals through the late Quaternary, the last major interglacial period taking place about 122,000 years ago. The common ancestor of the northern hemisphere species (i.e., I. pollicaris and I. lebeck ) likely dispersed and crossed the equatorial barrier separating the South China Sea from northern Australia, allowing a northern hemisphere population to differentiate from the I. sannio population. The separation of I. pollicaris in the temperate region from I. lebeck in the tropical region may have resulted from the combined effects of a geological barrier (e.g., Wallace’s Line; Huxley, 1868, with modification) and current dispersal (mainly Kuroshio Current), that is, the Wallace’s Line providing the geological barrier/separation during the ice age (with low temperature) and their larvae dispersed by the Kuroshio Current to both sides of Taiwan and northward to southern Japan.
Comparative materials. Ichthyscopus lebeck : CAS-SU 14135 View Materials (261 mm SL), Singapore, 7–12 May 1937 . THNHM-F11561 (349 mm SL), F11562 (368 mm SL), F11563 (269 mm SL), F11564 (277 mm SL), F11565 (287 mm SL), Ranong Fish Market , Ranong, Thailand, Andaman Sea, 6 Dec. 2014 . THNHM-F11997 (217 mm SL), Suksamran, Ranong, Thailand, Andaman Sea , 27 July 2015 . THNHM-F12531 (191 mm SL), F12533 (314 mm SL), F12534 (210 mm SL), Ban Tha Klang Pear , Suksamran, Ranong, Thailand, Andaman Sea, June 2016 . THNHM- F12677 (252 mm SL), Ranong Fish Market , Ranong, Thailand, Andaman Sea, 29 Aug. 2016 . Ichthyscopus sannio : AMS I.16796-100 (256 mm SL), Fam Beach , Queensland, Australia .

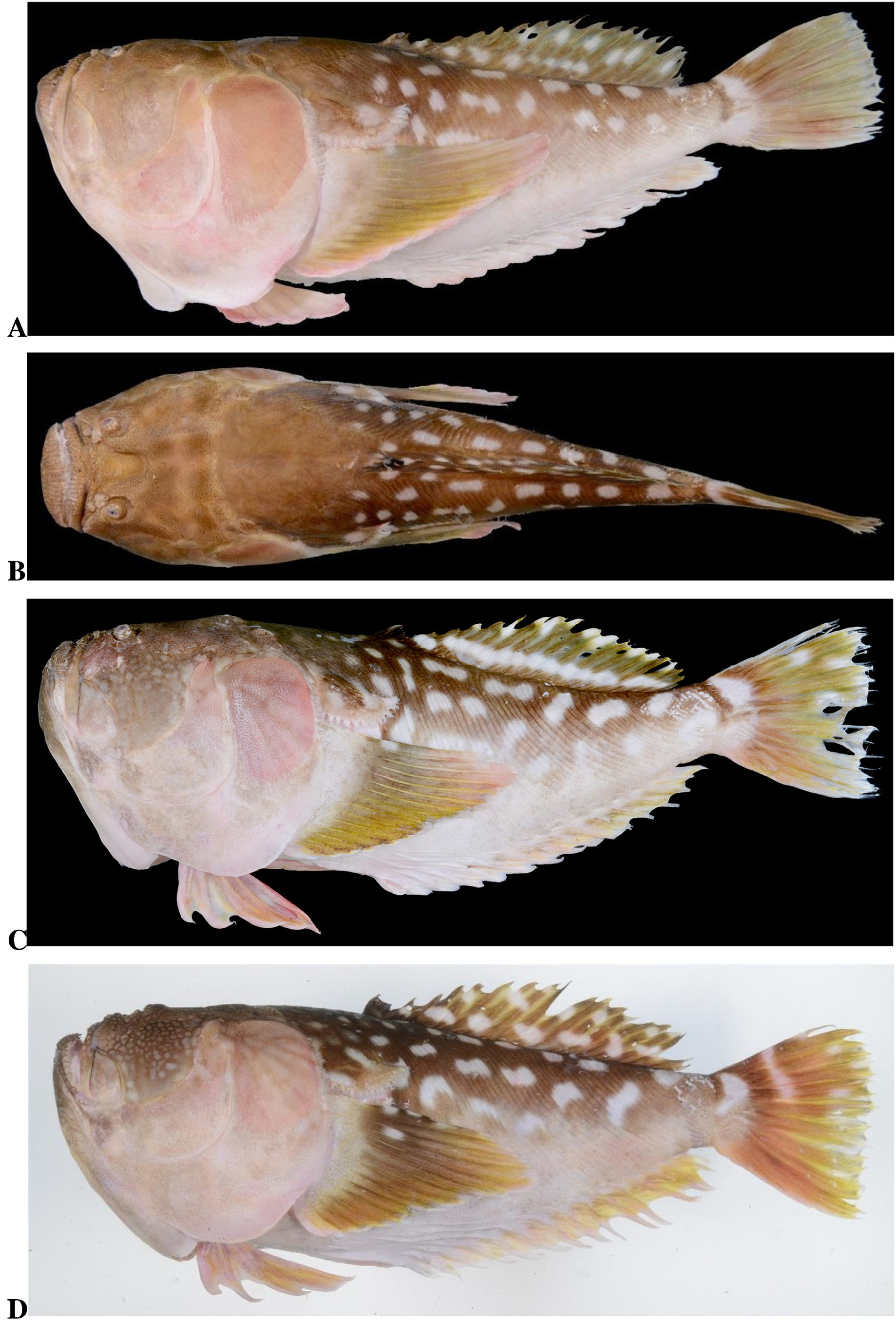
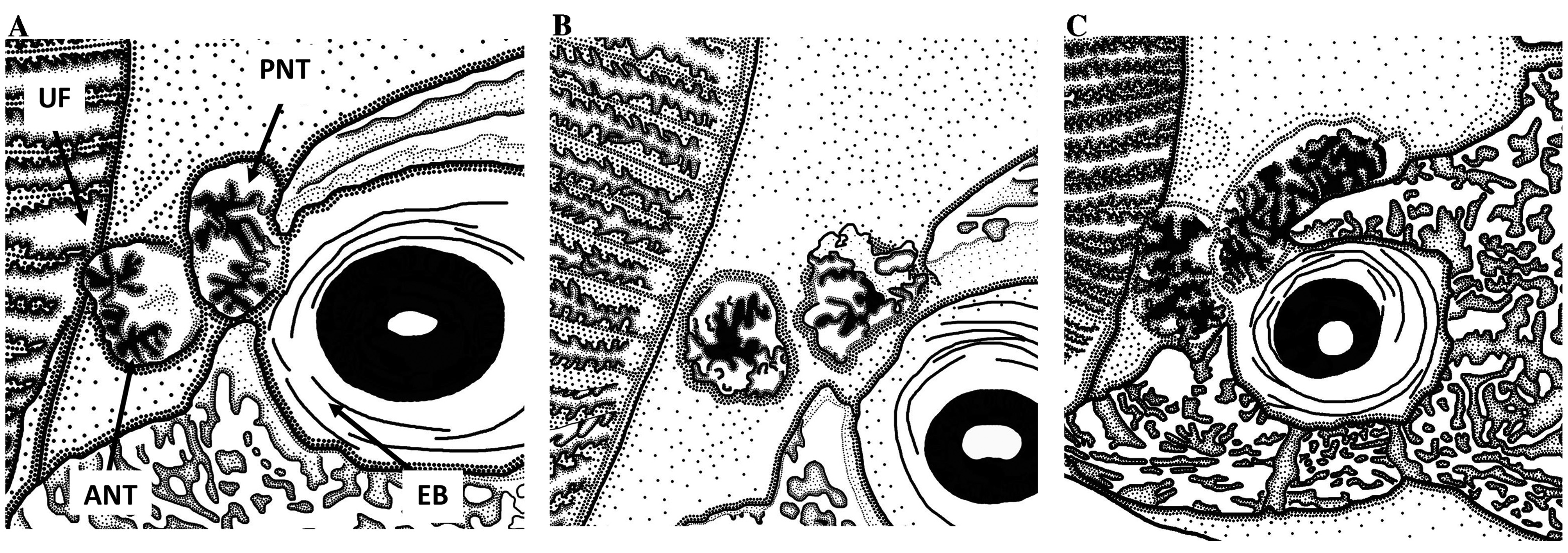
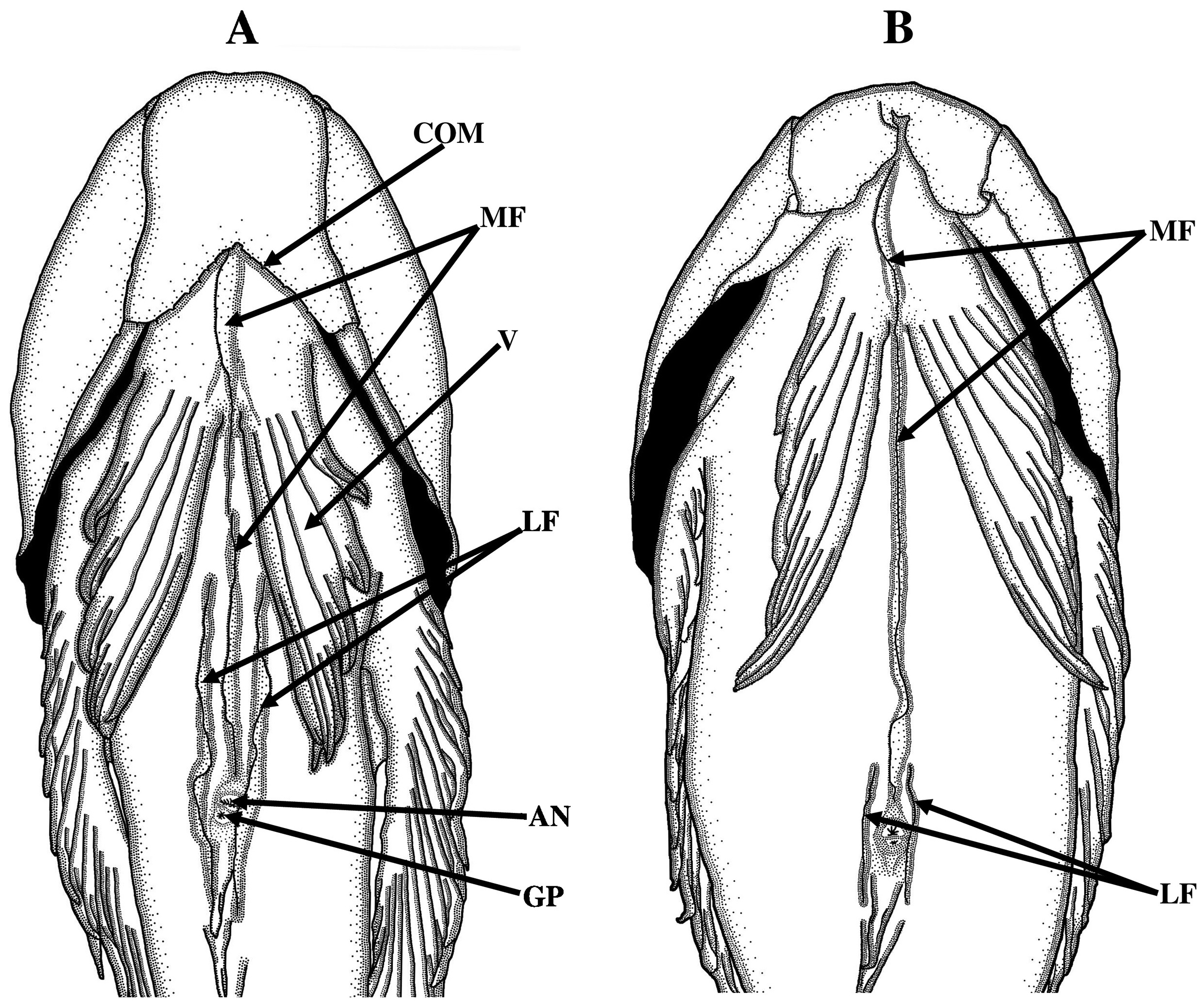
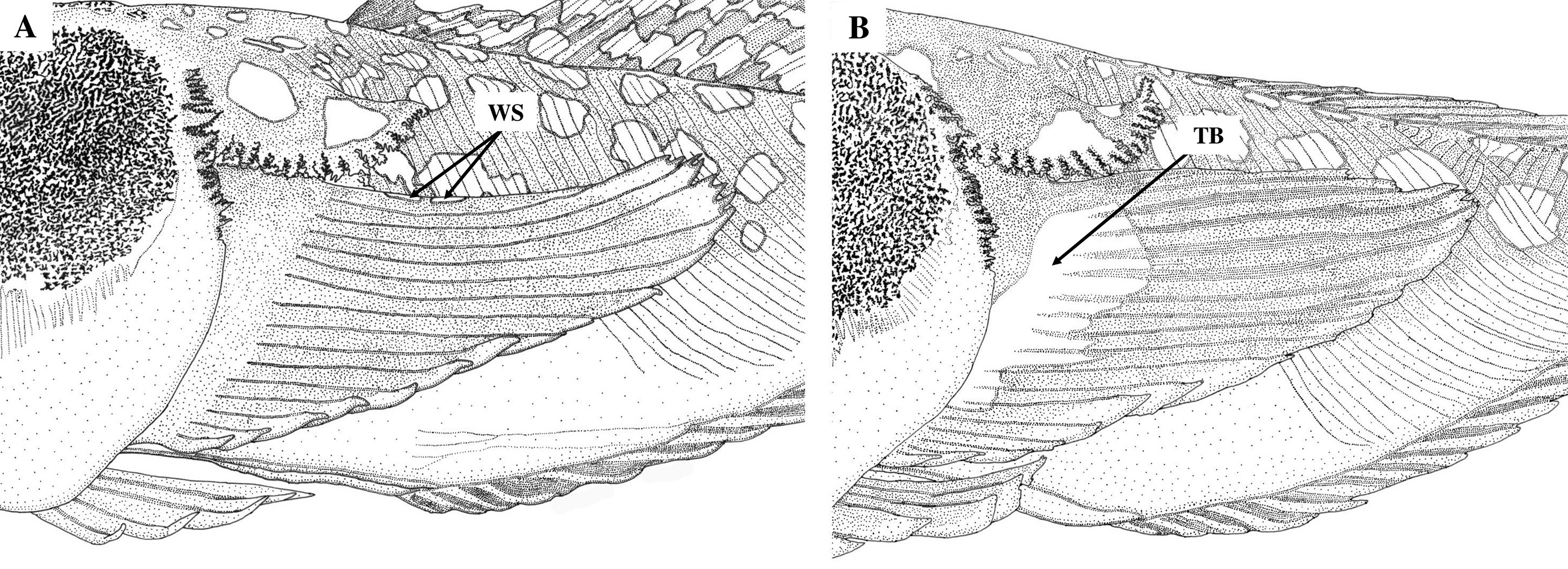

TABLE 1. Morphometric and meristic data of Ichthyscopus pollicaris sp. nov., comparing with the data of I. lebeck and I. sannio.
| I. pollicaris sp.nov. | I. lebeck | I. sannio * | ||
|---|---|---|---|---|
| Holotype | Paratypes (n =11) | (n =11) | (n =24) | |
| Standard length (mm) Measurements (% SL) | 187 | 160–308 Mean (Range) | 191–368 | 45.0–431 |
| Body depth | 35.8 | 34.5 (29.2–36.6) | 29.8–36.6 | 29.5–37.1 |
| Head length | 42.3 | 42.2 (40.2–43.8) | 39.7–43.3 | 30.9–36.9 |
| Head width | 31.6 | 32.1(28.6–35.1) | 29.3–34.3 | 31.0–38.1 |
| Pectoral fin length | 30.0 | 30.0(26.1–33.7) | 26.4–33.3 | 27.0–33.6 |
| Pelvic fin length | 24.1 | 25.0(23.9–26.9) | 21.6–28.1 | 22.4–26.2 |
| Measurements (% HL) | ||||
| Orbital diameter long | 12.7 | 13.4(12.5–15.1) | 10.1–13.6 | 13.1–18.7 |
| Orbital diameter trans | 10.1 | 10.7(9.6–12.3) | 9.1–10.6 | 10.2–16.1 |
| Eye diameter | 7.6 | 7.7(6.3–10.3) | 6.4–9.1 | 5.7–12.7 |
| Interorbital distance | 20.3 | 19.4(17.2–22.9) | 20.2–24.5 | 20.6–27.4 |
| Interorbital fossa width | 12.7 | 11.9(10.0–13.5) | 14.9–19.5 | 11.2–19.8 |
| Cleithral spine length | 21.5 | 20.0 (17.8–23.5) | 16.0–27.4 | 23.8–36.5 |
| Meristics | ||||
| Dorsal fin rays | II,18 | II, 17–18 | II, 17.5–19 | II–III, 16–17.5 |
| Anal fin rays | 16 | 16–17 | 17–19 | 16.5–17.5 |
| Pectoral fin rays | 17 | 16–18 | 18–19 | 17–18 |
| Vertebrae (abdominal+caudal) | 9+17 | 9+17 | 9+(16–17) | 9+18 |
| Epipleural ribs | 9 | 9–10 | 8–11 | 7–9 |
| Upper lip fimbriae (both sides) | 24 | 20–27 | 25–34 | 27–36 |
| Lower lip fimbriae (both sides) | 45 | 40–49 | 43–53 | 41–51 |
| Lower jaw teeth (both sides) | 11 | 10–18 | 11–20 | 12–21 |
| Opercular fimbriae | 17 | 15–21 | 15–22 | 12–19 |
| Cleithral flap fimbriae | 22 | 17–22 | 17–22 | 15–22 |
| Scale rows | 59 | 48–69 | 50–61 | 47–59 |
| NMV |
Museum Victoria |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Ichthyscopus pollicaris
| Vilasri, Veera, Ho, Hsuan-Ching, Kawai, Toshio & Gomon, Martin F. 2019 |
Ichthyscopus lebeck
| Shen, S. - C. 1984: 116 |
| Chen, J. T. - F. 1969: 430 |