
Alpheus viserion, Anker & Leray & Pachelle, 2021
|
publication ID |
https://doi.org/10.11646/zootaxa.4933.3.5 |
|
publication LSID |
lsid:zoobank.org:pub:61F5EC89-729B-4FA7-8337-F438D26D0CCA |
|
DOI |
https://doi.org/10.5281/zenodo.4567307 |
|
persistent identifier |
https://treatment.plazi.org/id/1B03015F-577D-FFC9-FF12-A503FC842F53 |
|
treatment provided by |
Plazi |
|
scientific name |
Alpheus viserion |
| status |
sp. nov. |
Alpheus viserion View in CoL sp. n.
( Figures 1–6 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 )
Type material. Holotype: male (cl 10.9 mm), FLMNH UF 51915 , Panama, Bocas del Toro, Isla Colón, Boca del Drago , 9°24’54.9”N 82°19’52.5”W, seagrass meadows, depth 2–3 m, under rock on sand, coll. M. Leray, 1 April 2019 View Materials GoogleMaps . Paratypes: 1 ovigerous female (cl 11.6 mm), FLMNH UF 51916 , same collection data as for the holotype GoogleMaps ; 1 male (cl 7.5 mm), FLMNH UF 51919 , same collection data as for the holotype GoogleMaps .
Description. Medium-sized species of Alpheus (cl range of type material: 7.5–11.6 mm). Carapace smooth, glabrous, not setose, not pubescent. Rostrum short, subtriangular, about twice as long as wide at base, subacute distally, almost reaching distal margin of first article of antennular peduncle; rostral carina feebly developed, rounded dorsally, flattening and broadening posteriorly between orbital hoods ( Fig. 1A, B View FIGURE 1 ). Orbital hoods well developed, moderately swollen, semi-open frontally, with anterior margin broadly rounded; margin between rostrum and orbital hoods slightly concave; adrostral furrows relatively shallow, margin between post-rostral carina and adrostral furrows relatively smooth, not abrupt ( Fig. 1A, B View FIGURE 1 ). Pterygostomial angle broad, rounded, not protruding anteriorly ( Fig. 1B View FIGURE 1 ); cardiac notch well developed.
Male first and second pleonal sternites each with sharp median process ( Fig. 1D View FIGURE 1 ). Female first and second pleonal sternites smooth ( Fig. 1I View FIGURE 1 ).
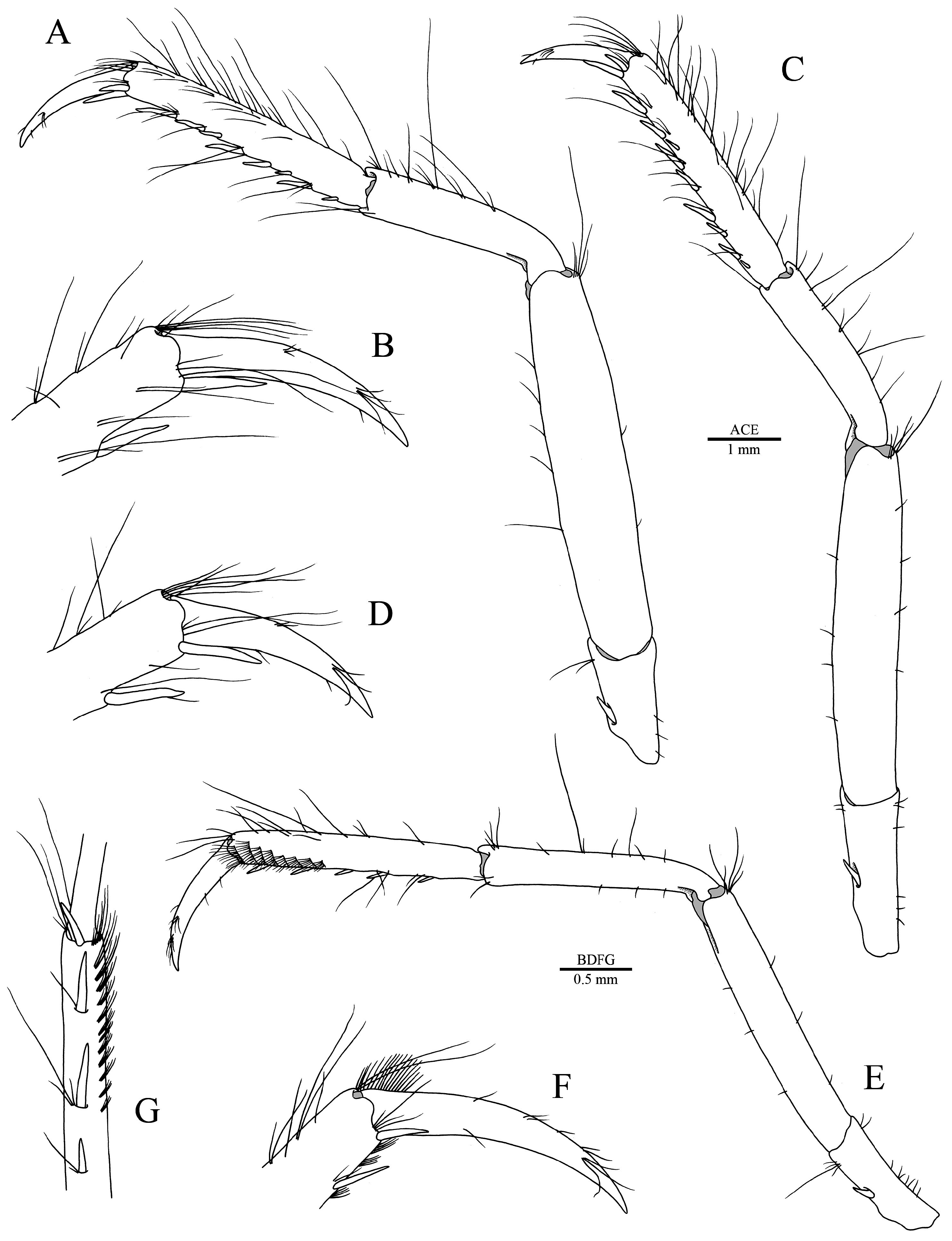
Telson broad, about 1.8 times as long as maximal width, somewhat tapering distally; lateral margins sinuose, without marked constriction; dorsal surface with two pairs of sharp stout cuspidate setae, both inserted at significant distance from lateral margin; proximal pair inserted at about half-length of telson, with cuspidate setae more widely spaced; distal pair inserted at about 0.7 length of telson; distal margin broadly rounded, fringed with long plumose setae and series of small spiniform setae of somewhat variable length; distolateral angles each with one pair of slender spiniform setae, lateral more slender and half as long as mesial ( Fig. 1E, F View FIGURE 1 ).
Antennular peduncle moderately stout; stylocerite slightly overreaching distal margin of first article of peduncle, with acute point; ventromesial carina of first article with small, triangular, distally subacute, anteriorly directed tooth; second article almost twice as long as wide; third article slightly longer than wide; antennular flagella noticeably uneven in length; mesial flagellum longest, diminishing in thickness towards terminal units; lateral flagellum thickened in its basal half, groups of aesthetascs starting at about eighth or ninth unit ( Fig. 1 View FIGURE 1 A–C).
Antenna with basicerite rather small, distolateral margin armed with slender, distally sharp tooth; scaphocerite subovate, tapering distally, about twice as long as its greatest width, lateral margin almost straight, faintly concave proximally, distolateral tooth very stout, sharp, clearly overreaching blade, reaching to or slightly beyond third article of antennular peduncle; carpocerite long, stout, overreaching both scaphocerite and antennular peduncle; flagellum stout, noticeably thicker than antennular flagella ( Fig. 1A, B View FIGURE 1 ).
Mouthparts typical for genus in external view. Third maxilliped relatively stout, pediform; coxa with lateral plate distally produced into small subacute tooth; antepenultimate article longest, lateral margin with longitudinal ridge delimiting shallowly depressed ventrolateral surface, mesial margin somewhat rugose; penultimate article about 0.3 times as long as antepenultimate article, about 2.5 times as long as wide proximally, widening distally; ultimate article about 3.5 times as long as wide, tapering distally, mesial surface furnished with several rows of thick serrulate setae, tip unarmed; much longer setae present on distodorsal margin of antepenultimate article, ventral surface of penultimate article and dorsal surface and apex of ultimate article; exopod long, reaching distal margin of antepenultimate article; arthrobranch very large ( Fig. 2A, B View FIGURE 2 ).
Major cheliped (= first pereiopod) with chela of edwardsii - type, with both dorsal and ventral notches of palm well developed and lateral grooves extending from dorsal notch on both mesial and lateral surfaces of palm; ischium very short; merus stout, about twice as long as wide; distodorsal angle blunt; mesial margin smooth, with short, simple, non-spiniform setae; distomesial angle with stout sharp tooth; distolateral angle smooth; carpus very short, cup-shaped, distodorsal margin with setae; chela slightly compressed, relatively slender, about 1.8 times as high (at maximal height) as long; surface smooth, not granulate, with numerous setae, especially on distal half of palm and fingers; dorsal shoulder rounded, not overhanging adjacent dorsal notch, latter deep, moderately broad; dorsomesial and dorsolateral surface of palm each with shallow longitudinal groove extending from dorsal notch posteriorly, reaching linea impressa on lateral surface; ventral shoulder rounded, not protruding forward, adjacent ventral notch deep; ventromesial and ventrolateral surface of palm each with broad, fairly deep oblique groove; fingers slightly longer than half-length of palm; distomesial margin of palm forming almost 90° angle, without tooth; distolateral margin smooth, without tooth; pollex shorter than dactylus, without oblique ridge on mesial surface; dactylus with distal margin broadly rounded; plunger stout, rounded anteriorly feebly bulging distally, with tuft of stamen-shaped sensillae; adhesive disks on dactylus and palm very small ( Fig. 3 View FIGURE 3 A–E). Male and female major chelipeds generally very similar, that of female slightly less robust and smaller (cf. Figs. 5A View FIGURE 5 , 6A View FIGURE 6 ).
Minor cheliped with chela simple, non-balaeniceps in both sexes. Male minor cheliped much smaller and weaker than major cheliped; ischium short; merus stout, about twice as long as wide; distodorsal angle blunt; mesial margin smoth, with simple, non-spiniform setae, distomesial angle with stout sharp tooth; distolateral angle smooth; carpus cup-shaped, more elongate than that of major cheliped; distodorsal margin setose; chela moderately stout; surface smooth, not granulate, very setose, especially on distal half of palm and fingers; palm about twice as long as high, devoid of notches or grooves, distodorsal angle slightly elevated; fingers subequal in length, approximately as long as palm, without balaeniceps ridges, not particularly expanded or modified otherwise; cutting edges bladelike, simple; adhesive disks on dactylus and palm very small ( Fig. 3 View FIGURE 3 F–H; see also Fig. 5A View FIGURE 5 ). Female minor cheliped similar to that of male, somewhat more slender (cf. Figs. 5A View FIGURE 5 , 6A View FIGURE 6 ).
Second pereiopod slender; ischium long, about six times as long as wide; merus slightly longer than ischium; carpus subdivided into five units, first longest, almost seven times as long as wide; ratio of carpal units approximately equal to 3.5 – 2.5 – 1 – 1 – 1.7; chela slightly longer than distal-most carpal unit, with fingers slightly longer than palm, furnished with several clusters of simple setae ( Fig. 2C View FIGURE 2 ).
Third pereiopod moderately slender; ischium about 1.6 times as long as wide, ventrolateral surface with stout cuspidate seta; merus about 4.2 times as long as wide; distomesial margin unarmed; distodorsal margin furnished with long setae; carpus about five times as long as wide, about half as long as merus, much slenderer than merus; propodus about six times as long as wide; ventral margin with row of eight or so spiniform setae in addition to one pair of stouter spiniform near articulation with dactylus; dactylus simple, conical, somewhat elongate, with gentle curvature, about half-length of propodus ( Fig. 4A, B View FIGURE 4 ). Fourth pereiopod similar to third, noticeably slenderer; ischium about 2.5 times as long as wide, ventrolateral surface with stout cuspidate seta; propodus with ventral margin armed with row of eight or so spiniform setae in addition to one distal pair of spiniform setae near dactylar base; dactylus somewhat shorter and stouter than than of third pereiopod, otherwise similar ( Fig. 4C, D View FIGURE 4 ). Fifth pereiopod slenderest of all; ischium with ventrolateral surface armed with stout spiniform seta; merus almost six times as long as wide; carpus slightly shorter than merus, about 5.5 times as long as wide; propodus with row of five spiniform setae in addition to one distal spiniform seta near dactylar base, on ventromesial side; distal half of propodus with about eight transverse rows of serrulate setae forming cleaning brush; dactylus almost 0.6 times as long as propodus, otherwise similar to that of third pereiopod ( Fig. 4 View FIGURE 4 E–G).
First to fifth pleopods without spiniform setae on ventrolateral margin of protopod in both sexes ( Fig. 2D, E, G, H View FIGURE 2 ). Male first pleopod with endopod reduced, much smaller than exopod ( Fig. 2D View FIGURE 2 ). Male second pleopod with endopod and exopod similar in size; appendix masculina small, about 0.6 times as long as appendix interna, with stiff setae along most of its surface ( Fig. 2E, F View FIGURE 2 ). Female first and second pleopods similar to those of male; protopod with long flexible setae for egg retention ( Fig. 2G, H View FIGURE 2 ). Developing embryos moderately large, about 0.8 mm long and 0.6 mm wide ( Fig. 1J View FIGURE 1 ).
Uropod with endopod and exopod broadly rounded; exopod broader and slightly longer than endopod, both fringed with plumose setae; protopod with distolateral and distomedial lobes each produced into strong triangular tooth; exopod with sharp distolateral tooth and stout distolateral spiniform seta; diaeresis sinuous, with broadly rounded tooth laterally, near base of distolateral spiniform seta; endopod with row of small spiniform setae of variable length on distal margin, interrupted by cluster of few long setae ( Fig. 1G, H View FIGURE 1 ).
Colour pattern. Body and appendages generally appearing as olive-green; carapace, pleon, telson and uropods with olive-green to brown-greenish background, speckled with pale greenish-yellow chromatophores, many interconnecting and forming nets of greenish-yellow spots and chains; antennular peduncles dark brown-green, stylocerite with pale green spots; antennal scaphocerite greenish, with some larger pale green blotches; antennular and antennal flagella pale olive-green; major chela almost uniformly olive-green on mesial surface, except for small cluster of paler blotches between linea impressa and dorsal notch, one large pale green area on proximal half of palm, and contrasting pinkish fingertips; minor cheliped uniformly olive green on mesial surface, with paler palm; second to fifth pereiopods pale olive-green to olive-yellow ( Figs. 5 View FIGURE 5 , 6 View FIGURE 6 ).
Etymology. The new species’ name refers to Viserion, one of Daenerys Targaryen’s three dragons of the popular TV series “Game of Thrones”. The dragon-related name was originally inspired from the Spanish name of the collection site, Boca del Drago (= dragon’s mouth). Used as a noun in apposition.
Distribution. West Atlantic: presently known only from the type locality on the Caribbean coast of Panama.
Ecology. All three specimens were collected under large rocks on fine white sand approximately 1–2 m away from a dense meadow of turtlegrass ( Thalassia testudinum Koenig ), at a depth of about 2– 3 m.
Remarks. Alpheus viserion sp. n. is morphologically closest to several species of the transisthmian, taxonomically challenging A. armillatus H. Milne-Edwards, 1837 species complex (see below), which was studied with three molecular markers by Mathews & Anker (2009). The western Atlantic members of the A. armillatus complex were subsequently revised by Anker (2012). The eastern Pacific species of the A. armillatus complex, among them two taxonomically problematic species and at least two undescribed ones ( Mathews & Anker 2009), will be revised in the near future (A. Anker, in prep.).
All members of the A. armillatus species complex are characterised by the rostral carina conspicuously flattened between the orbital hoods, forming a more or less well-defined, V-shaped or U-shaped post-rostral plate, rising above the usually deep adrostral furrows; the major chela of a basic edwardsii - type (see above); the merus of the cheliped armed with a strong distomesial tooth; the minor chela non-balaeniceps in both sexes; the ambulatory pereiopods with simple, conical dactyli; and the third and fourth pereiopods with distomesially unarmed meri (modified from Anker 2012). Additionally, in several species, the pleon is armed with sternal processes, usually on the first two sternites, sometimes on the first four sternites ( Kim & Abele 1988; McClure 2002; Anker 2012). The morphological features of A. viserion sp. n. are generally consistent with the characteristics of the A. armillatus complex, although the post-rostral plate of the new species is less obvious and not abruptly delimited from the adjacent adrostral furrows as in most species of the A. armillatus complex ( Fig. 1A View FIGURE 1 ).
Among the western Atlantic members of the A. armillatus complex, only two species are characterised by the feebly demarcated, roughly V-shaped post-rostral plate, viz. A. carlae Anker, 2012 and A. angulosus McClure, 2002 ( McClure 1995, 2002; Anker 2012), both of them also occurring in Bocas del Toro (Anker 2012). All other western Atlantic species of the A. armillatus complex have abruptly delimited, V- or U-shaped post-rostral plates and present additionally a series of other morphological differences and/or have different colour patterns (Anker 2012).
Alpheus carlae has a wide distribution in the western Atlantic, ranging from Florida through the Caribbean Sea to southern Brazil (Anker 2012). The species seems to be genetically strongly structured ( Mathews & Anker 2009) and exhibits a great deal of variation in both morphology and colouration, although the colour pattern type is essentially the same (Anker 2012). Alpheus viserion sp. n. seems to share more morphological characters with A. carlae than with any of the other species of the A. armillatus complex. These include the identical armature of the pleonal sternum (with a median process on each of the first two pleonites, although with some variation); the similar proportions of the major chela; the presence of a cuspidate seta on the ischium of the fifth pereiopod; and the appendix masculina being distinctly shorter than the appendix interna ( Figs. 1D View FIGURE 1 , 2F View FIGURE 2 , 3A, D View FIGURE 3 , 4E View FIGURE 4 ; cf. Anker 2012: figs. 41H–J, 42A, B, 43O). The new species can be separated morphologically from A. carlae by at least three relatively subtle characters, viz. the shape of the post-rostral area (with an anteriorly wide post-rostral carina, broadening and flattening between the eyes in A. viserion sp. n. vs. with an anteriorly narrow post-rostral carina, widening and flattening only posterior to the level of the eyes in A. carlae ); the major chela dactylus longer and with a much less bulging plunger; and the male minor chela without a trace of a ventral notch on the palm (vs. with a slight sinus in A. carlae ) ( Figs. 1A View FIGURE 1 , 3E, H View FIGURE 3 ; cf. Anker 2012: figs. 41A, 42D–F, 43F, K). In addition, the colour patterns of A. viserion n. sp. and A. carlae are radically different. The new species has uniformly olive-green background colour, speckled with pale green-yellow spots, and a homogenously coloured major and minor chelae ( Figs. 5 View FIGURE 5 , 6 View FIGURE 6 ). In contrast, in A. carlae , the pleon displays transverse bands of red-brown, green-brown or dark brown colour, with at least two pairs of large black lateral spots, and the chelae are brownish speckled with white blotches or dots (Anker 2012: figs. 44–48).
Alpheus angulosus is another genetically highly structured western Atlantic snapping shrimp, occurring from South Carolina to southern Brazil and throughout the Caribbean Sea and Gulf of Mexico ( Mathews & Anker 2009; Anker 2012; Heuring et al. 2017). As in the previous species, the populations of A. angulosus exhibit some variation in the colouration of the body and chelipeds (Anker 2012), which apparently is not directly linked to the genetic structure of the species ( Heuring et al. 2017). Alpheus viserion sp. n. differs from A. angulosus by the presence of a strong median sternal process on each of the first two pleonites in males (vs. with a weak median sternal process on the first pleonite and without such a process on the second pleonite in males of A. angulosus ); the telson noticeably narrower, about 1.8 times as long as maximally wide (vs. broader, about 1.5 times as long as wide, in A. angulosus ); the blade of the antennal scaphocerite distally wider and approaching more the point of the distolateral tooth (vs. distally narrower and with the distal margin greatly exceeded by the distolateral tooth in A. angulosus ); the male major chela with the fingers longer relative to the palm and with the dactylar plunger shorter and more truncate (i.e. less bulging distally); the male minor chela without trace of a ventral notch (typically present in larger specimens of A. angulosus ); the ischium of the fifth pereiopod armed with a stout cuspidate seta (vs. unarmed in A. angulosus ); and the appendix masculina much shorter than the appendix interna (vs. the two being subequal in length in A. angulosus ) ( Figs. 1A, D, E View FIGURE 1 , 2F View FIGURE 2 , 3A, H View FIGURE 3 , 4E View FIGURE 4 ; cf. Anker 2012: figs. 19D, H–K, 20A, D, E). In addition, the two species present clear differences in their colour patterns, despite a generally similar, uniform colouration of the carapace and pleon. In A. viserion sp. n., the background of the body is olive-green to olive-brown, speckled with pale green-yellow dots ( Figs. 5 View FIGURE 5 , 6 View FIGURE 6 ), whilst in A. angulosus , in contrast to the new species, the background varies from pale greenish-grey to bluish-grey and is speckled with darker brownish or dark greenish-red dots (Anker 2012: figs. 22–26). Furthermore, in A. viserion sp. n., the palm of the major chela is uniformly olive-green in dorsal view (= mesial side of the chela), except for a paler patch on the proximal half of the palm ( Figs. 5A View FIGURE 5 , 6A View FIGURE 6 ), whereas in A. angulosus , the palm of the major chela is generally more patterned, for instance, the dorsal and ventral notches and adjacent oblique grooves are always conspicuously marked with white or pale yellow (Anker 2012: figs. 22–26).
The only eastern Pacific species of the A. armillatus complex, which has a smoothly delimited post-rostral plate, is A. tenuis Kim & Abele, 1988 , known from Panama to Mexico ( Kim & Abele 1988; Villalobos Hiriart et al. 1989; Mathews & Anker 2009). The taxonomic identity of A. tenuis is presently being investigated due to the presence of two genetically distinct and apparently geographically at least partly overlapping clades in the eastern Pacific ( Mathews & Anker 2009). Nevertheless, A. viserion sp. n. can be easily separated from A. tenuis sensu Kim & Abele (1988) by the major chela palm with the dorsal shoulder not protruding and not overhanging the dorsal notch (vs. clearly overhanging it in A. tenuis ); the ventromesial carina of the first article of the antennular peduncle bearing a subacute, anteriorly directed tooth (vs. with a low blunt tooth in A. tenuis ); the longer stylocerite, slightly overreaching the distal margin of the first article of the antennular peduncle (vs. not reaching this margin in A. tenuis ); and the minor chela with the fingers subequal to the palm (vs. with the fingers much longer than the palm in A. tenuis ). Finally, the two species also have different colour patterns ( Figs. 5 View FIGURE 5 , 6 View FIGURE 6 ; cf. Mathews & Anker 2009: fig. 2D) and are allopatrically isolated by the Isthmus of Panama.





| FLMNH |
Florida Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
InfraOrder |
Caridea |
|
Family |
|
|
Genus |