
Maechidius Macleay, 1819
|
publication ID |
https://doi.org/10.5852/ejt.2020.721.1127 |
|
publication LSID |
lsid:zoobank.org:pub:89E62EF8-2E45-4C59-94B7-6A5603E8939B |
|
DOI |
https://doi.org/10.5281/zenodo.4344293 |
|
persistent identifier |
https://treatment.plazi.org/id/1A3787F6-9D11-9254-FDEA-FA14CCEF2DB1 |
|
treatment provided by |
Valdenar |
|
scientific name |
Maechidius Macleay, 1819 |
| status |
|
Maechidius Macleay, 1819 View in CoL
Maechidius Macleay, 1819: 14 View in CoL .
Epholcis Waterhouse, 1875: 192 View in CoL . syn. nov.
Paramaechidius Frey, 1969: 503 View in CoL . syn. nov.
New and restored combinations
The following new combinations are proposed: Maechidius acutus ( Narakusumo & Balke, 2019) comb. nov., M. arcuatus ( Narakusumo & Balke, 2019) comb. nov., M. bilobiceps ( Fairmaire, 1877) comb. nov., M. cakalele ( Narakusumo & Balke, 2019) comb. nov., M. divergens ( Waterhouse, 1875) comb. nov., M. gracilis ( Waterhouse, 1875) comb. nov., M. longior (Blackburn, 1898) comb. nov., M. moluccanus Moser, 1920 comb. rest., M. obiensis ( Narakusumo & Balke, 2019) comb. nov., M. uniformis ( Britton, 1957) comb. nov. (all from Epholcis ), M. agnellus ( Prokofiev, 2018) comb. nov., M. clypeatus ( Frey, 1969) comb. nov., M. helleri ( Frey, 1969) comb. nov., M. heterosquamosus Heller, 1910 comb. rest., M. opatroides Arrow, 1941 comb. rest., M. pauxillus Heller, 1910 comb. rest., M. perlatus ( Frey, 1969) comb. nov., M. popei ( Frey, 1969) comb. nov. and M. speciosus ( Frey, 1969) comb. nov. (all from Paramaechidius ).
Sexual dimorphism
The taxa of Maechidius are sexually dimorphic. Arrow (1941: 449) was the first to describe sexual dimorphism in Maechidius and noted that differences in external morphology are considerable between the two sexes. He listed several sexually dimorphic features: “greater development of the clypeal lobes of the males”, “more slender front tibiae with feebler teeth” [in males], “longer tarsi, especially in the hind legs” [in males], “slightly longer lamellae of the antenna” in male ( Britton 1957).
According to my own observations, the best feature for sexing certain Maechidius sensu novo species besides a considerable difference in length of male and female antennal lamellae, is the shape of the inner distal spur of the protibia, which in many Papuan and Wallacean taxa is straight and pointed in females but less slender and distally curved inward in males (cf. Figs 449–450 View Figs 436–450 ). However, at least in male M. lineatopunctatus Frey, 1969 , the protibial terminal spur is straight and pointed, whereas in M. nanus Arrow, 1942 , M. nepenthephilus sp. nov., M. rugicollis Moser, 1920 and M. woodlarkianus Heller, 1914 protibial terminal spurs are absent in both sexes. The same is true for the lower terminal spur of the metatibia, which appears curved inward in males but straight in females ( Figs 438–440 View Figs 436–450 ). In other morphological groups the male terminal spur of the protibia is absent or short and hardly curved distally, but straight and longer in the female or basally curved and afterwards straight in both sexes ( Figs 447–448 View Figs 436–450 ). In many males of Papuan species (both with the male protibial terminal spur curved or straight) the lower meso- and metatibial terminal spurs are also curved ( Figs 449–450 View Figs 436–450 ). Metatibial terminal spur(s) can be longer in the female than in the male in certain species ( Figs 441–442 View Figs 436–450 ). In many species, the male and female pygidium is different as well (e.g., flat or convex dorsally in male, concave, humped posteromediad or longitudinally impressed in female) ( Figs 451 View Figs 451–468 –534). The shape of the labroclypeus is often considerably different between males and females ( Figs 9–10 View Figs 8–13 , 97–196 View Figs 97–108 View Figs 109–120 View Figs 121–132 View Figs 133–144 View Figs 145–156 View Figs 157–168 View Figs 169–180 View Figs 181–192 View Figs 193–207 ), in most cases males having more stronger protruding anterolateral angles of the labroclypeus and the anterior margin is more deeply emarginate, but opposite cases are known (see descriptions below).
Ecology
Generally, Australian Maechidius are considered silvicolous and connected with diverse forest types ( Williams 2002). The Papuan Region species are reported from lowland up to mid-montane rainforests. Many (if not all) Australian taxa are termitophiles and can be found in termite nests ( Lawrence & Ślipiński 2013). Adults of the Australian Maechidius cavus Britton, 1957 were observed feeding on palm trunks and trunks of “native trees” in Northern Territory, “causing severe damage at a property” ( Northern Territory Government 2011). Australian Maechidius caviceps Blackburn, 1888 , M. fissiceps Macleay, 1888 , M. rugosicollis Macleay, 1873 and M. bilobiceps ( Fairmaire, 1877) comb. nov. were reported to assemble under the loose bark of dead or alive termite-invaded Eucalyptus spp. in the forest ( Dodd 1921), sometimes in thousands of specimens. The same author reported M. gracilis ( Waterhouse, 1875) comb. nov. from flowers ( Dodd 1921, as “ Epholcis pacilis ” [sic♂]). Most Papuan species were sampled at light (e.g., Arrow 1941; Telnov personal observations) and Australian species are also known to be attracted to lights ( Dodd 1921; Britton 1957). As already pointed out by Arrow (1941: 449), and I can confirm it from my own observations, some of the night-collected specimens have clay adhering on both the dorsum and venter, which points to their burrowing behaviour (hiding during the daytime). In my study, Maechidius lobaticeps Frey, 1969 and M. nepenthephilus sp. nov. were observed (see Descriptions) on flowers of Nepenthes sp. pitcher plants ( Figs 751–753 View Figs 751–753 ) and many were found being digested inside pitchers situated below the racemes. Nepenthes are known to be insect-pollinated plants. In this particular situation, M. lobaticeps were eudominants among all other insects observed both on Nepenthes sp. flowers and in pitchers in the Lake Sentani area (only two specimens of M. nepenthephilus sp. nov. were collected). It is worth mentioning that the area around Doyo Lama village in the Sentani area is a semi-dry eucalypt forest on very poor soil (woody vegetation consists of bushy Eucalyptus sp. only and a grass layer – mainly of ferns and pitcher plants with scarce bunches of hard grass). According to my knowledge, this is the first evidence of such an interaction between Maechidius and Nepenthes .
Dodd (1921) reported copulation of Australian Maechidius bilobiceps comb. nov. during the daytime under loose bark of Eucalyptus spp. This author reported a noteworthy passive defence mechanism of the same M. bilobiceps comb. nov. observed in crevices and under loose bark of eucalypt: “… a sudden movement will cause them to drop suddenly; first one will fall to the ground, then others, until in a few seconds hundreds are raining down and in the course of a couple of minutes not one is left on the treetrunk where just before they were in such numbers” ( Dodd 1921: 62).
Surprisingly, the Sulawesi species M. babyrousa sp. nov. and M. deltouri sp. nov. were attracted to yellow pan traps (visiting blossoming plants is not yet recorded for Maechidiini ), except for M. lobaticeps and M. nepenthephilus sp. nov. record mentioned above.
Species groups
Based on the studied specimens and new generic synonymy, the following four informal species-groups are herewith established to include most morphologically uniform taxa:
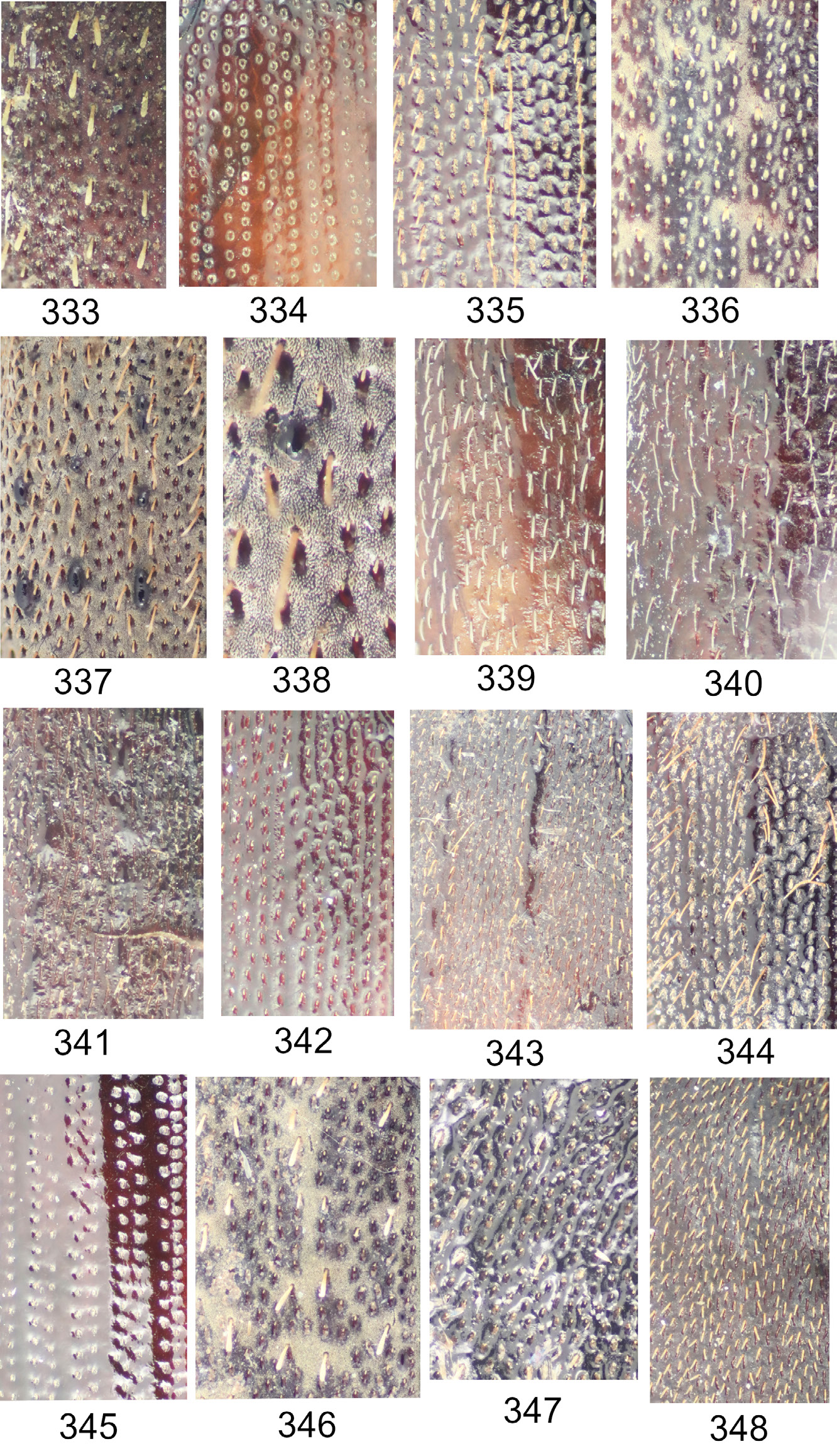
Maechidius sturnus group, based on strongly irregularly arranged punctures forming conspicuous fingerprint-like pattern on dorsal pronotum and elytra ( Figs 297 View Figs 286–301 , 347 View Figs 333–348 ).
Maechidius dani group, based on ventrally dentate male protarsomeres 1–2 and humped visible abdominal sternite III. This group includes the Baliem Valley ( Papua) species M. dani sp. nov. and M. hamatus sp. nov.
Maechidius pauxillus group, based on the vestiture of the body (clavate scale-like setae, microscopical velvety pubescence), the shape of the pronotum and the type of dorsal punctures. This group includes the Moluccan M. agnellus (Prokofiev) comb. nov. and the Papuan M. bintang sp. nov., M. crypticus sp. nov., M. heterosquamosus Heller comb. rest., M. dendrolagus sp. nov., M. lapsus sp. nov., M. merdeka sp. nov., M. owenstanleyi sp. nov., M. pauxillus Heller, 1910 comb. rest., M. popei (Frey) comb. nov., M. speciosus comb. nov. and M. weigeli sp. nov.
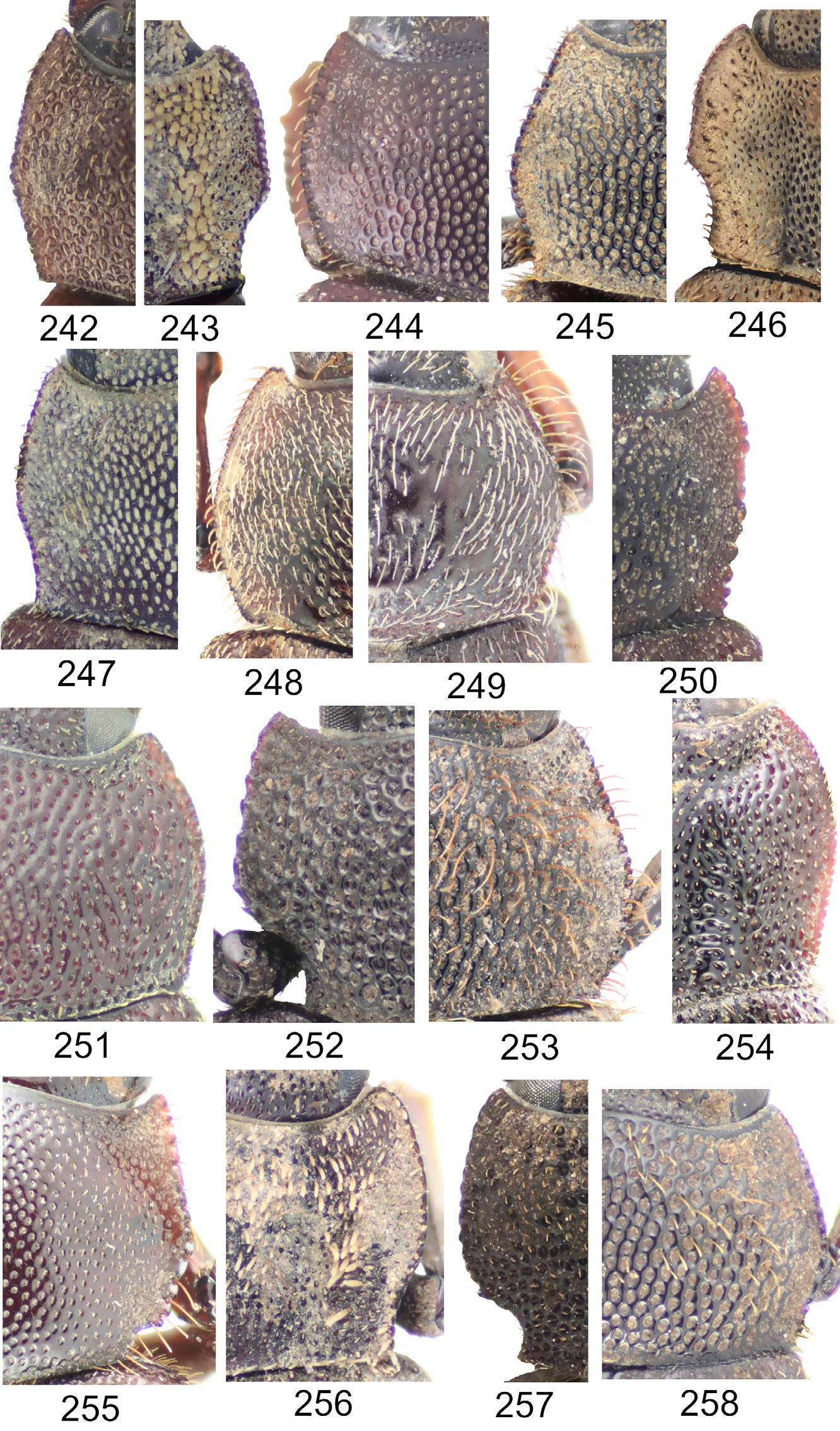
Maechidius ursus group, based on the flattened and laterally expanded pronotum and brush-like clustered setae of the dorsum ( Figs 238 View Figs 226–241 , 246 View Figs 242–258 , 261 View Figs 259–265 , 357 View Figs 349–358 ). This group includes the Papuan species M. kazantsevi sp. nov., M. opatroides Arrow, 1941 comb. rest., M. perlatus ( Frey, 1969) comb. nov. and M. ursus sp. nov.
Descriptions
In this chapter redescriptions of previously known species are generally less comprehensive than those of new taxa, intentionally omitting differential diagnoses, ecology and distribution. Species recently (re)described by Narakusumo & Balke (2019) are not redescribed again, and for those of Arrow (1941) and Frey (1969) only additional features or corrections are given. For the previously described species measurements are only given when necessary, e.g., where they provide additional data.

















No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Melolonthinae |
|
Tribe |
Maechidiini |
Maechidius Macleay, 1819
| Telnov, Dmitry 2020 |
Paramaechidius
| Frey G. 1969: 503 |
Epholcis
| Waterhouse C. O. 1875: 192 |
Maechidius
| Macleay W. S. 1819: 14 |