
Bujurquina omagua, Říčan & Říčanová, 2023
|
publication ID |
https://doi.org/ 10.5852/ejt.2023.870.2127 |
|
publication LSID |
lsid:zoobank.org:pub:60245C9C-34EE-4F68-A346-F7A684E18EB6 |
|
DOI |
https://doi.org/10.5281/zenodo.8006645 |
|
persistent identifier |
https://treatment.plazi.org/id/8E478708-58F1-4E97-960C-9FCD93107C9A |
|
taxon LSID |
lsid:zoobank.org:act:8E478708-58F1-4E97-960C-9FCD93107C9A |
|
treatment provided by |
Felipe |
|
scientific name |
Bujurquina omagua |
| status |
sp. nov. |
Bujurquina omagua sp. nov.
urn:lsid:zoobank.org:act:8E478708-58F1-4E97-960C-9FCD93107C9A
Figs 6–10 View Fig View Fig View Fig View Fig View Fig ; Table 1 View Table 1
Bujurquina sp. Oran Říčan et al. 2022.
Bujurquina sp. Peru 1 Říčan et al. 2022.
Diagnosis
Bujurquina omagua sp. nov. is highly apomorphic compared to other described species in the genus because it is unique in several character states. These include: 1) shape and pattern of the suborbital stripe in adults, which is bend and with a blotch at the postero-ventral end (vs straight or absent, with or without a blotch), with an ontogenetic change with juveniles and subadults having a straight stripe running diagonally postero-ventrally and starting from below the eye in juveniles (i.e., not bend as in adults) and without a blotch, 2) orange cheek and lower head coloration with alternating opalescent lines and series of spots on snout and spotted on cheek and lower head, 3) spinous portion of dorsal fin with a single thin diagonal black line per membrane, or two lines per membrane in holotype and largest adults (vs unpatterned in all Southern group species, or with circular blotches, with several thick diagonal black lines per membrane, or two thick horizontal bands across the dorsal fin in all other Northern group species), 4) longest snout of all species without overlap (mean 15.5%, 14.4–16.4% of SL; followed by B. oenolaemus with a mean of 11.4%, 10.3–13.1 of SL), and 5) most delicate LPJ, expressed by being shortest of all species with widest horns (length/width ratio 0.42–0.45 vs 0.50–0.64 in all other species), one of the smallest dentigerous areas (length of dentigerous area/width of LPJ ratio 0.33–0.34 vs 0.34–0.44 except B. tambopatae , B. moriorum and B. huallagae where also 0.33–0.34), and the least robust dorso-ventrally (bone at posterior end of dentigerous area less thick or equal to length of largest posterior teeth vs much thicker than teeth in other species).
Bujurquina omagua sp. nov. is additionally diagnosed by the following unique combination of morphometric characters, the most pronounced of which characterize the robust head of the species: 1) second to third longest head (after B. oenolaemus and together with B. labiosa ; both with a mean 37.0%, 36.3–38.0% of SL in B. omagua vs mean 39.1%, 38.6–40.4% of SL), 2) third longest preorbital distance (after B. robusta and B. oenolaemus ; mean 9.3%, 8.5–10.0% of SL), 3) large interocular distance (largest mean value of all species, 12.5% of SL vs 9.7–12.4% however with large overlap of ranges with the other species), 4) rather wide head (mean 19.9%, range 19.1–20.7% of SL), 5) rather long preorbital distance (mean 9.3%, range 8.5–10.0% of SL, third longest based on mean value), 6) robust head which has rather small eyes (mean 11.3%, range 10.3–12.0% of SL), 7) very deep body (mean 45.1%, range 43.2–46.8% of SL, based on mean the second after B. vitatta ), but 8) with rather long and shallow caudal peduncle (CPL mean 13.8%, range 12.6–15.6% of SL; CPD mean 16.6%, range 15.5–17.1% of SL), and 9) rather short last spine in the dorsal fin (mean 15.6%, range 12.3–17.1% of SL).
Etymology
The specific epithet ‘ omagua ’ is a noun in apposition given after the Omagua people, which were at the time of first contact with Europeans in the 16 th century the dominant people along the banks of the Amazon River upstream from the mouth of the Negro River well into Peru.
Type material
Holotype PERU • adult ♂, 97.4 mm; Amazonas river basin, Department Loreto, Maynas Province , District Las Amazonas , close to Oran village, Quebrada Sabalillo , cabeceras upper locality P18-17; 3° 27´08.4˝ S, 72° 29´11.8˝ W; 97 m a.s.l. based on GPS, 115 m a.s.l. based on Google Earth; 28 Jun. 2018; Říčan leg.; clear-water stream with blackish tinge; ID tag 1189; MUSM 70225 ( Figs 6 View Fig , 8–9 View Fig View Fig ). GoogleMaps
Paratypes
All from Peru, Amazonas river basin, Department Loreto, Maynas Province , District Las Amazonas , close to Oran village, 9 ex.
PERU • 1 adult, 78.6 mm; same collection data as for holotype; ID tag 1188; GenBank: OP436199 ; MUSM 70224 • 1 adult, 98.9 mm; same collection data as for holotype; ID tag 1190; GenBank: OP436200 ; MUSM 70226 ( Figs 7–9 View Fig View Fig View Fig ) • 1 adult, 69.7 mm; same collection data as for holotype; ID tag 1191; GenBank: OP436201 ; MUSM 70227 ( Fig. 7 View Fig ) • 1 adult, 77.1 mm; same collection data as for holotype; ID tag 1192; GenBank: OP436202 ; MUSM 70228 ( Fig. 7 View Fig ) • 1 adult, 78.7 mm; Amazonas river basin, Department Loreto, Maynas Province , District Las Amazonas, close to Oran village , Qebrada Sabalillo on trail from mouth, P18-15; 3° 27´45.6˝ S, 72° 29´22.5˝ W; 90 m a.s.l. based on GPS, 110 m a.s.l. based on Google Earth; 27 Jun. 2018; Říčan leg.; clear-water stream with blackish tinge; ID tag 1182; GenBank: OP436196; MUSM 70221 ( Figs 7 View Fig , 9 View Fig ) GoogleMaps • 1 adult, 79.9 mm; Amazonas river basin, Department Loreto, Maynas Province , District Las Amazonas, close to Oran village , cabeceras of Quebrada Sabalillo , lower loc., P18-16; 3° 27´17.5˝ S, 72° 29´22.8˝ W; 75 m a.s.l. based on GPS, 117 m a.s.l. based on Google Earth; 28 Jun. 2018; Říčan leg.; clear-water stream with blackish tinge; ID tag 1185; GenBank: OP436197; MUSM 70222 ( Figs 7 View Fig , 10 View Fig ) GoogleMaps • 1 adult, 92.6 mm; same collection data as for preceding; ID tag 1186; GenBank: OP436198 ; MUSM 70223 ( Fig. 7 View Fig ) GoogleMaps • 1 adult; Amazonas river basin, Department Loreto, Maynas Province , District Las Amazonas, close to Oran village , small unnamed quebrada just before entering Oran creek , P18-21; 3° 27´21.2˝ S, 72° 30´43.2˝ W; 82 m a.s.l. based on GPS, 104 m a.s.l. based on Google Earth; 3 Jul. 2018; Říčan leg.; clear-water stream with blackish tinge; ID tag 1200; GenBank: OP436204; MUSM 70229 ( Fig. 7 View Fig ) GoogleMaps • 1 juvenile; same collection data as for preceding; GenBank: OP436205 ; MUSM 70230 GoogleMaps .
Description
Compound, based on eight specimens from Sabalillo creek, 69.7–98.9 mm. Refer to Figs 6–10 View Fig View Fig View Fig View Fig View Fig . Measurements summarized in Table 1. View Table 1
SHAPE. Moderately elongate, rather robust, deep bodied (body depth mean 45.1%, range 43.2–46.8% of SL). Head especially robust, long (head length mean 37.0%, 36.3–38.0% of SL), wide (head width mean 19.9%, range 19.1–20.7% of SL), with large interocular distance (mean 12.5%, range 11.6–14.2% of SL). Frontal outline straight, nape and snout curved, anterior half of dorsal-fin base contour curved. Ventral outline curved. Snout long (mean 15.5%, 14.4–16.4% of SL). Preorbital distance long (mean 9.3%, range 8.5–10.0% of SL), preorbital bone much longer than high. Robust head with rather small eyes (mean 11.3%, range 10.3–12.0% of SL). Lips rather narrow, not turgid. Corner of mouth not reaching vertical from anterior margin of orbit, especially in largest specimens. Rather long and shallow caudal peduncle (CPL mean 13.8%, range 12.6–15.6% of SL; CPD mean 16.6%, range 15.5–17.1% of SL), and rather short last spine in dorsal fin (mean 15.6%, range 12.3–17.1% of SL).
SCALES. Squ. long. (E1) scales 24 (5), 25 (3); 2 scales between L1 and dorsal fin anteriorly, 2 scales between L1 and L2. Upper lateral line (L1) with 15 (1), 16 (7) scales. Lower lateral line (L2) with 9 (4), 10 (3), 11 (1) scales; canal bearing scales on caudal fin: 0 in dorsal, 0-1 in median and 0 in ventral sequence. Cheek scales in 3 (8) series. Anterior portion of caudal fin scaly.
FINS. Dorsal fin XII.11 (1), XIII.10 (1), XIII.11 (1), XIV.9 (1), XIV.10 (4). Soft anal fin pointed, 3 rd ray longest, reaching to middle of caudal fin or slightly beyond. Anal fin III.8 (8). Pectoral fin with rounded tip, reaching to above genital papilla to anal-fin origin (to 2 nd spine in one specimen); Pectoral fin 13 (8). Pelvic fin pointed, first ray slightly produced, reaching to origin of anal fin or base of 1 st spine of anal fin. Caudal fin rounded, without well-developed streamers or with only short streamers.
GILL-RAKERS. 1 epibranchial, 1 in angle, and 5 (3), 6 (4), 7 (1) ceratobranchial rakers externally on first arch.
JAW TEETH. An outer series of stronger, conical, sharply pointed, slightly recurved teeth, slightly increasing in size anteriorly, not worn, 15–22 in outer hemiseries in upper jaw.
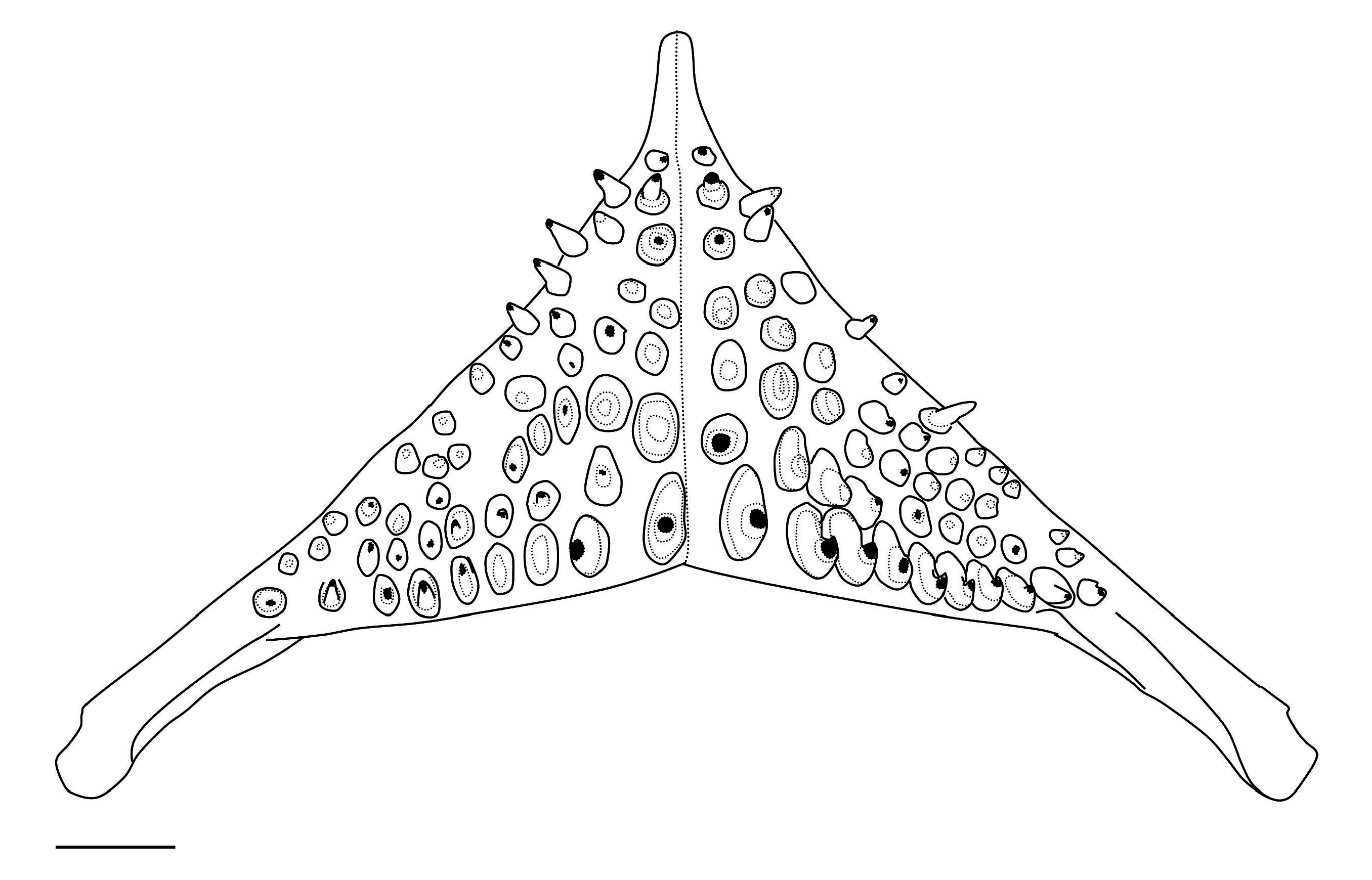
LOWER PHARYNGEAL JAW AND TEETH. Lower pharyngeal jaw (LPJ) as examined in two specimens (79.8– 98.9 mm) ( Fig. 10 View Fig ) very delicate, most delicate LPJ of all described species of Bujurquina , very shallow vertically dorso-ventrally (bone at posterior end of dentigerous area less thick than length of largest posterior teeth in the smaller specimen, almost so in the larger specimen vs thicker than the teeth in all other species). LPJ also shortest and widest of all species, length/width ratio 0.42–0.45 (vs 0.50–0.64 in all other species). LPJ tooth-plate (due to the longest arms within Bujurquina ) also one of the shortest in genus, length of dentigerous area/width of LPJ ratio 0.33–0.34 (vs 0.34–0.44 except B. tambopatae , B. moriorum and B. huallagae where also 0.33–0.34), Teeth slightly laterally compressed (most pronounced in posterior row), slightly bicuspid with main cusp posterior, central and posterior teeth largest, smaller towards outside, central teeth with flat-worn apex, others with pronounced tip.
Coloration in alcohol
Coloration limited to melanin patterns in preserved state, ground colour light yellowish-brownish, lighter towards whitish on chest and anterior abdomen, dusky greyish on anterior head, snout, and dorsum. Dark diffuse and wide interorbital band. Operculum silvery greyish.
Up to four clearly defined grey lines between eye and mouth corner and on anterioir cheek interspersed with spots, rest of cheek as well as opercular series richly spotted. Suborbital stripe rather marrow and of uniform width, diffuse and ghost-like, bend from behind eye forward to cheek and then postero-ventrally to a variously developed suborbital stripe blotch in adults (never ocellated and with diffuse borders). In one collected juvenile specimen and in subadults straight (i.e., not bend), running from below eye (vs from behind eye) diagonally postero-ventrally.
Midlateral band continuous from head to penultimate vertical bar on body, strongly pigmented and black, very wide anteriorly and gradually narrowing posteriorly. No interruption in vertical bar 5 (sensu Říčan et al. 2005), midlateral blotch (usually present and dominant in bar 5 in cichlasomatine cichlids) is absent. Lateral band continued on head across nape close behind eye.
Body with five vertical bars plus one caudal peduncle bar and a caudal peduncle blotch (usually vertically elongated). The five body bars correspond to ontogenetic bars 2–5 with the anteriormost ontogenetic bars 6–7 fused into one wide bar below midlateral band but visible as two above it. Posterior body bars (2–4) narrow and tightly spaced, anterior bars (5, 6+ 7) wide and widely spaced. Bars darkest dorsally, reaching ventrally approximately to level of lower edge of caudal peduncle. Bars do not extend onto dorsal fin.
Body scale centres (below midlateral band) with dark blotches, more pronounced in zones of vertical bars.
Dorsal fin light grey, lappets indistinct, with a single narrow grey diagonal stripe per membrane in spinous part, with blotches on ventral portion of soft part. Anal fin light grey, with spots on last about four membranes. Caudal fin light grey with minute slightly elongate dark dots on anterior half, distally membranes hyaline with grey edges. Pelvic fin light grey, edge and produced portion of first ray white.
Coloration in life
Melanin patterns as in preserved. Lower head, cheek, and opercular series yellow-orange, with opalescent golden to azure blue lines and spots, arranged as up to four clearly defined lines between eye and mouth corner and on anterioir cheek interspersed with spots, rest of cheek as well as opercular series richly spotted. Fins orange, pelvic fin yellow, edge and produced portion of first ray white (opalescent). Spinous portion of dorsal fin with a single thin diagonal black line per membrane in juveniles and subadults, and with two lines per membrane in holotype and large adults. Lower lip pale greyish to indistinctly light azure blue. Midlateral blotch present only as a paler reflective area within the dark midlateral band (hence absent from coloration of preserved specimens).
Habitat
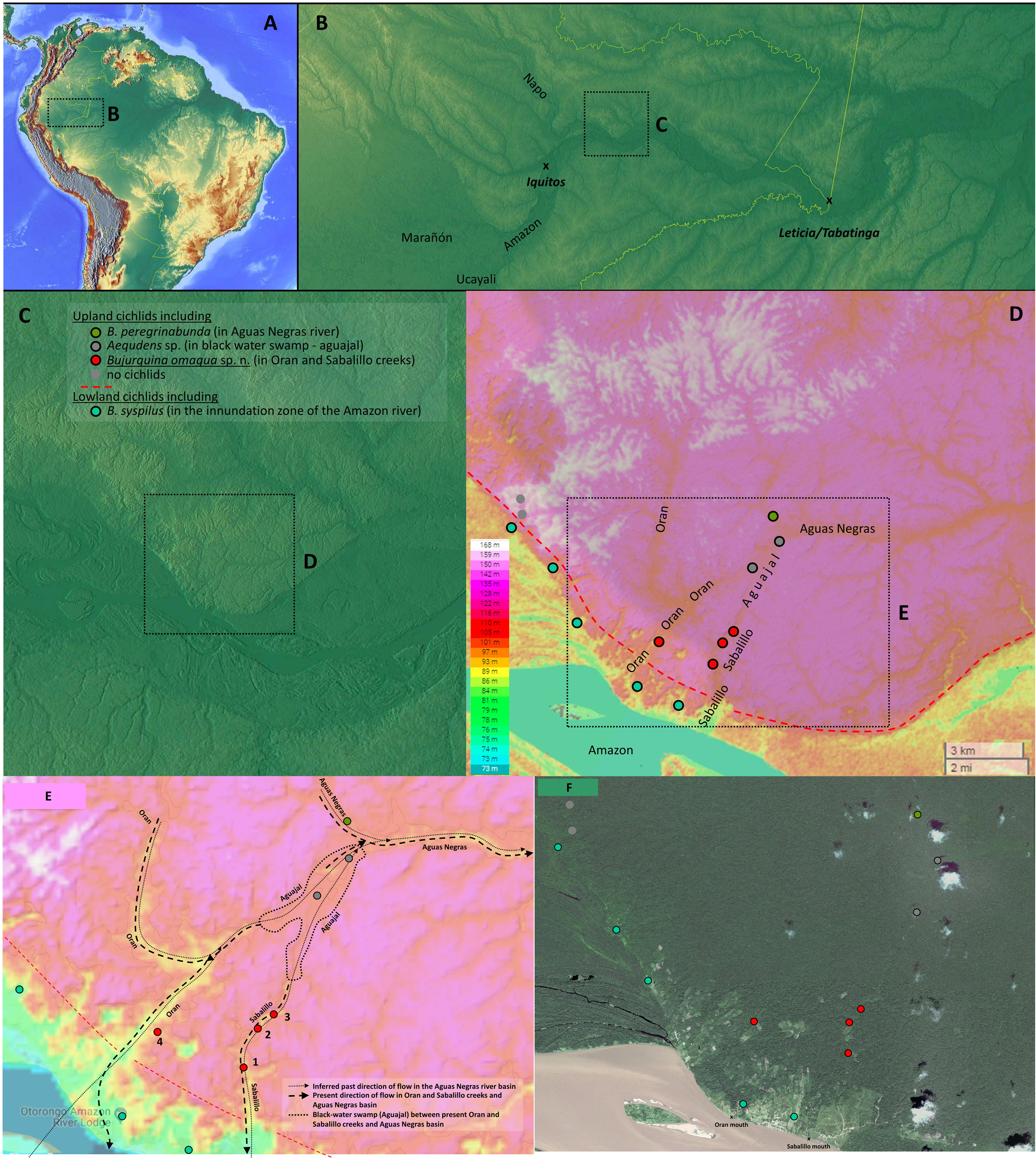
Bujurquina omagua sp. nov. was found only in small rainforest streams within the basins of the Sabalillo and Oran creeks. The localities ( Fig. 11 View Fig ) were around 2 m wide streams, with shallow water (less than 1 m except in deeper pools) but deep mud and accumulated debris, blackish water, and only weak or moderate current. Bujurquina omagua has not been collected from the main streams of the Oran creek or from the lower sections of the Oran and Sabalillo creeks ( Fig. 12 View Fig ), which have muddy water and where B. syspilus was found.
Distribution
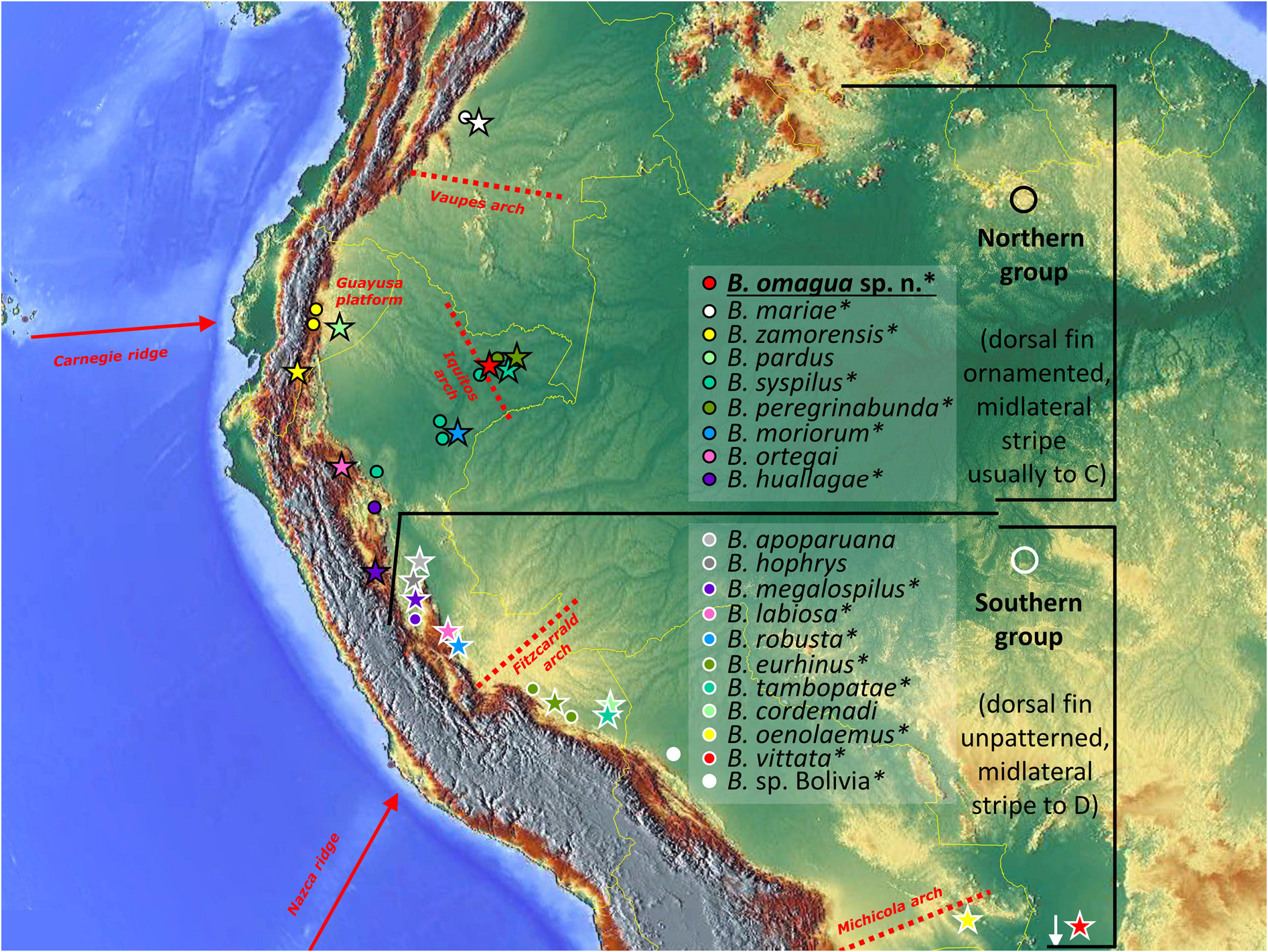
Bujurquina omagua sp. nov. is only known from the Sabalillo and Oran creeks which empty into the Amazon River on either side of the village of Oran located on the first high ground just downstream from the mouth of the Napo into the Amazon, District Las Amazonas, Maynas Province, Department Loreto, Perú ( Figs 1 View Fig , 13 View Fig ).
Morphometric differentiation of Bujurquina omagua sp. nov.
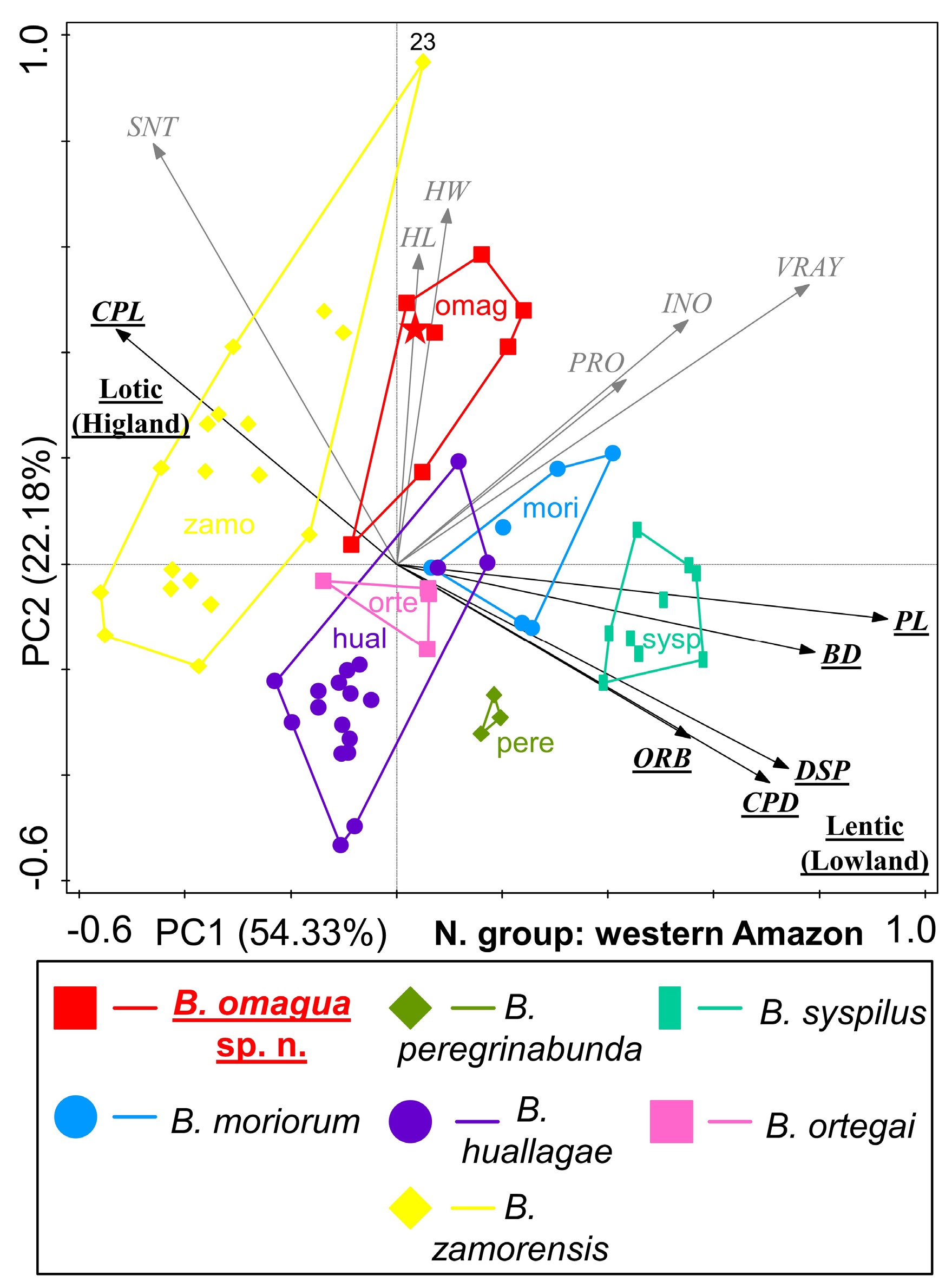
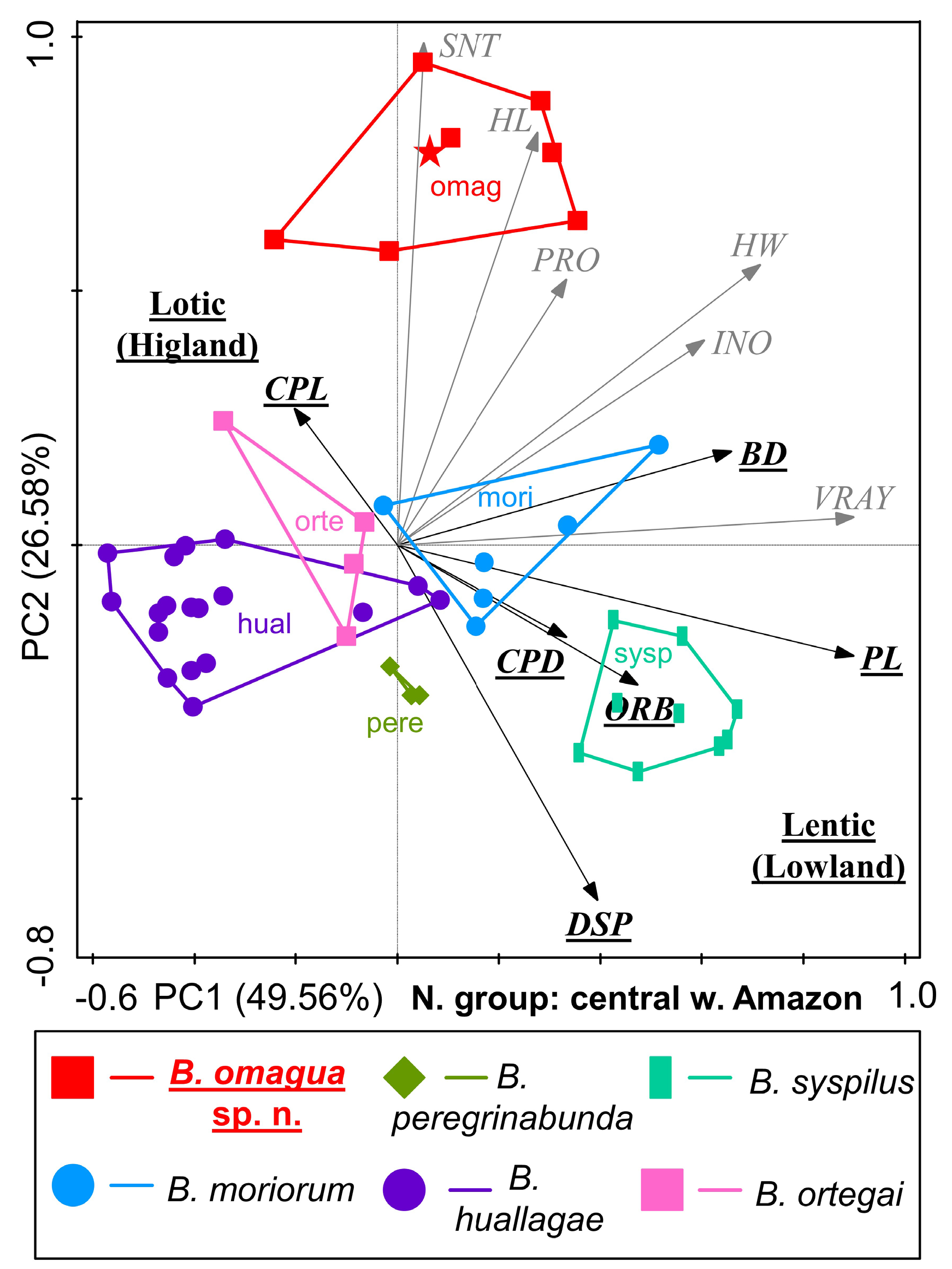
Bujurquina omagua sp. nov. has a highly apomorphic head and body shape that sets it apart from the sympatric/parapatric species. When analyzed through PCA these species are arranged along the main tentic-lotic axis as corresponding to their lowland-upland habitats ( Figs 14–15 View Fig View Fig ). The lotic/lentic ecomorphology dichotomy is in the PCA analyses best correlated with caudal peduncle length (long vs short), caudal peduncle depth (narrow vs deep), body depth (narrow vs deep), the size of the eye (orbital diameter; small vs large), pectoral fin length (short vs long), and last dorsal fin spine length (short vs long).
The remaining characters are not correlated with the main ecomorphological dichotomy (i.e., interocular distance, head width, head length, preorbital distance, snout length, and first ventral fin ray length) ( Figs 14–15 View Fig View Fig ).
The lotic/lentic ecomorphology dichotomy is best visible in the analysis including all central western Amazonian species ( Figs 1 View Fig and 14 View Fig ), while the diagnostic characters of Bujurquina omagua sp. nov. are best visible in the analysis excluding the westernmost and most highland species B. zamorensis ( Fig. 15 View Fig ). Bujurquina omagua is in the PCAs distinguished from the sympatric/parapatric species predominantly by characters not correlated with the lotic-lentic, namely large head, wide head, long snout, and long preorbital dostance. All these diagnostic characters are in line with the diagnosis of the species based on means and ranges of the morphometric characters (see above).
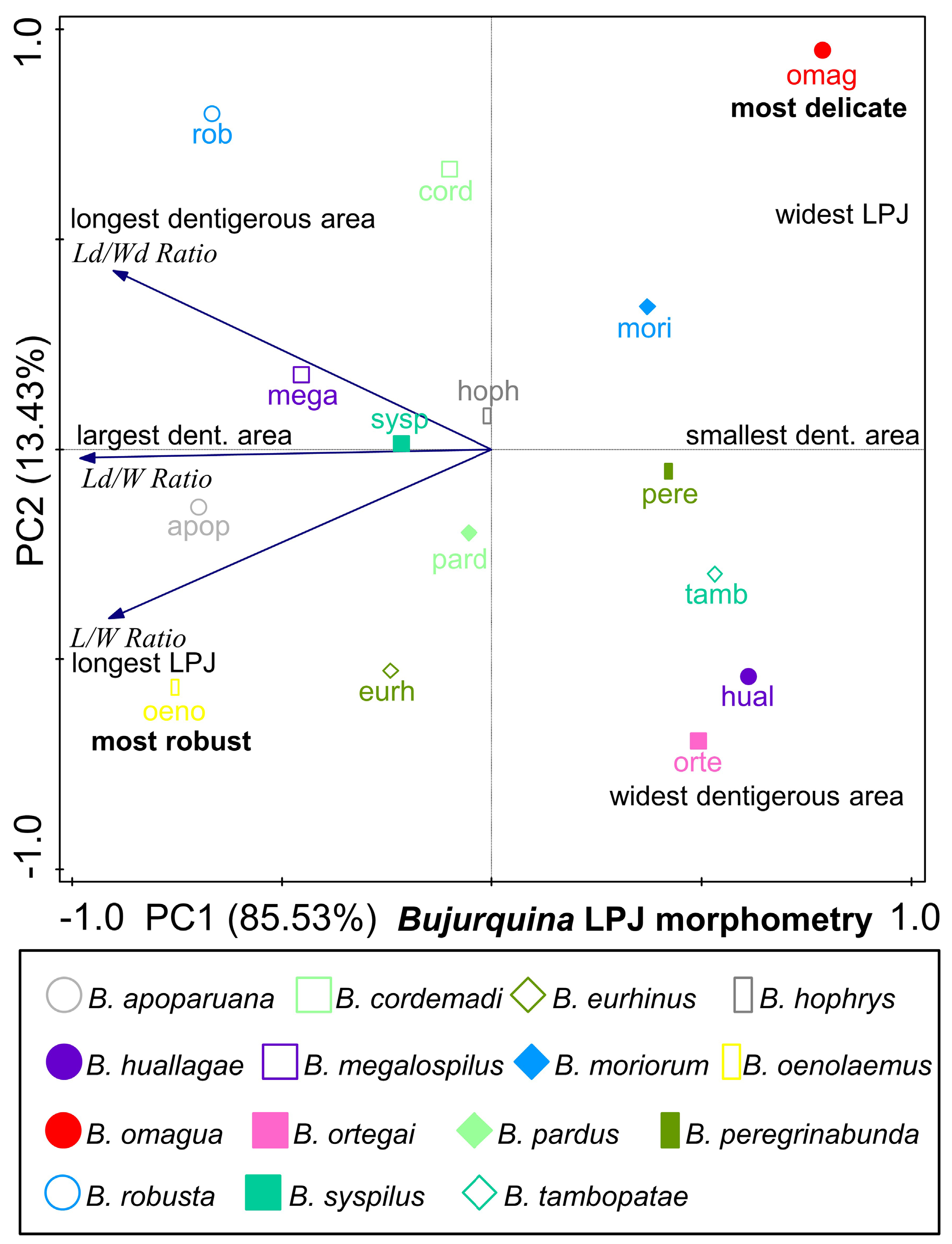
Bujurquina omagua sp. nov. has an outlying LPJ shape that can be described as the most delicate LPJ found among the valid species of Bujurquina ( Fig. 16 View Fig ). The LPJ of Bujurquina omagua (studied in two specimens, see above) as expressed by proportional measurements is the shortest and widest of all species (L/W ratio) and the LPJ tooth-plate is the shortest in the genus (Ld/W ratio). Additionaly to this the LPJ is very shallow vertically dorso-ventrally, but this character was scored as binary, in comparison to the length of teeth (see Results), and thus not included in the PCA analysis.
Paradoxically, the most delicate LPJ of B. omagua sp. nov. is associated with one of the largest heads among species of Bujurquina (together with B. labiosa ) and with the longest snout among species of Bujurquina . A similar head and snout shape in B. oenolaemus is however associated with the most robust LPJ ( Fig. 16 View Fig ) of this molluscivorous species.
A similar combination of a long snout and large head with a very delicate LPJ as in B. omagua sp. nov. is however also known from the cichlid genus Australoheros Říčan & Kullander, 2006 in the species A. forquilha Říčan & Kullander, 2008 and A. ykeregua Říčan, Piáleck, Almirón & Casciotta, 2011 ( Říčan & Kullander 2008; Říčan et al. 2011).
Phylogeny of Bujurquina and phylogenetic position of Bujurquina omagua sp. nov.
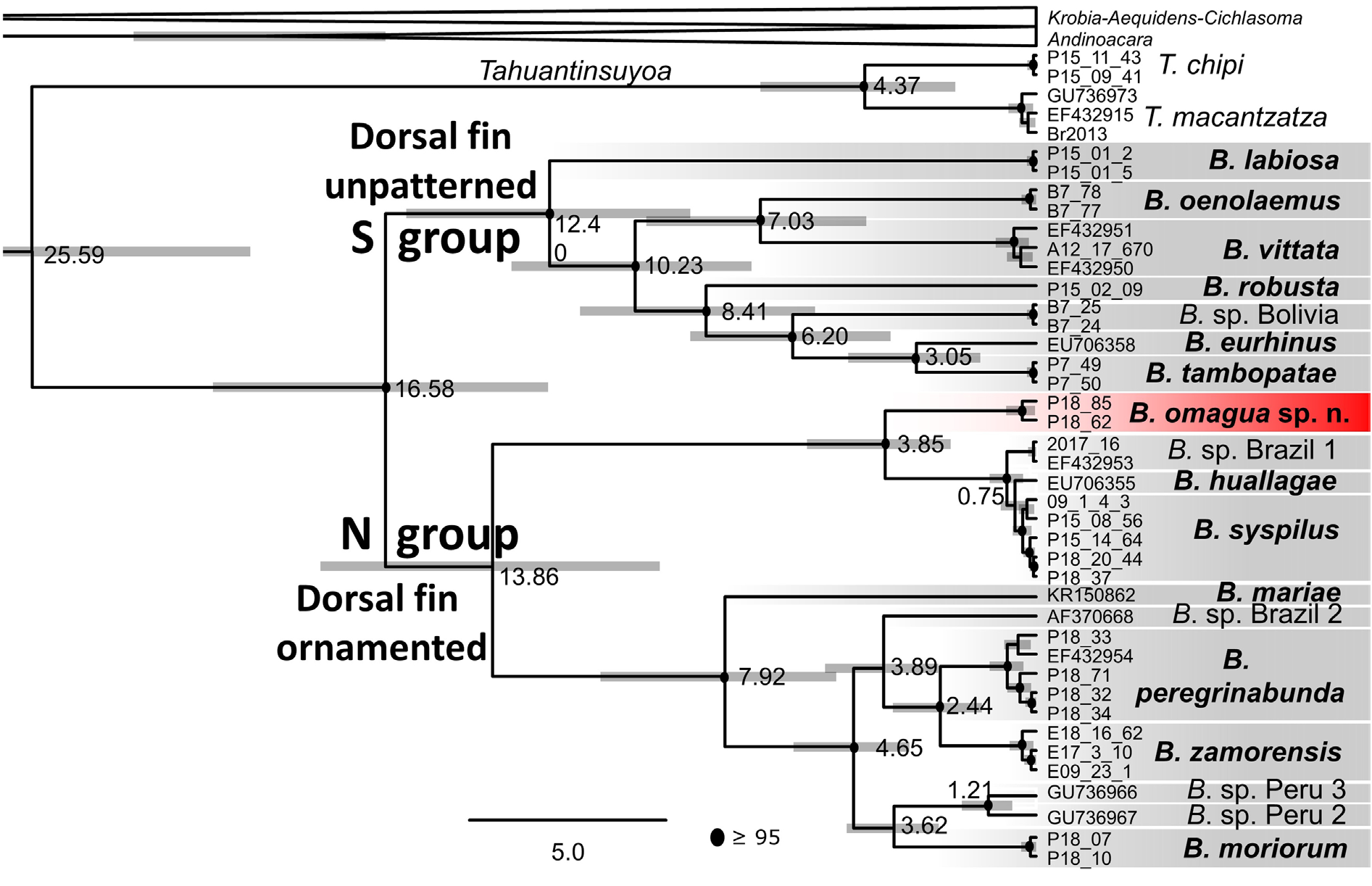
Phylogenetic analyses of the 1055 characters cytb data matrix performed with MP in PAUP* (including all ingroup sequences) and BI analyses in MrBayes and BEAST (performed with the corresponding haplotypes data matrix) provided robust and very similar results. BI runs in independent analyses with the best-fitting model of evolution (GTR+I+G) converged well (ESS>200 for all parameters; burn-in 10%) and led to identical topologies under the same settings. The NJ and MP analyses were done with the complete dataset, the BI and BEAST analyses with the haplotype dataset. No phylogenetic differences were found between the MP, BI and BEAST analyses, and hence only the BEAST analysis is shown in Fig. 17 View Fig .
The phylogenetic reconstructions divide Bujurquina into two main clades that can be diagnosed by patterned vs unpatterned dorsal fin, corresponding in geographical terms to Northern vs Southern group ( Figs 1 View Fig and 17 View Fig ). The main biogeographic dichotomy within the genus between the Southern and Northern groups is within the present upper Ucayali basin in Peru ( Fig. 1 View Fig ).
The two main clades of Bujurquina are dated as having diverged at 16.6 Ma. Within the Southern group the two southern-most species from Argentina, Paraguay, Brazil and Bolivia are sister species ( B. vittata and B. oenolaemus ), and they are a sister group to the clade composed of the Peruvian B. eurhinus and B. tambopatae as sister species, followed by the Bolivian B. sp. Bolivia and then the Peruvian B. robusta . The phylogenetically basal-most species of the Southern group is the Peruvian B. labiosa . The Peruvian species are thus in basal positions within the Southern group, and they also have the longest internodes and the earliest dates of divergence based on molecular clock dating, with the time frame of divergence among the Peruvian species dated between 12.4 Ma and 3.0 Ma ( Fig. 17 View Fig ).
The Northern group of Bujurquina is made up of two highly divergent clades ( Fig. 17 View Fig ) unlike the situation in the Southern group. The first clade within the Northern group shows a younger basal divergence at 3.9 Ma and includes, as a basal species, the here described B. omagua sp. nov., and then, among the Peruvian species, B. huallagae and B. syspilus . Samples of the morphologically and biogeographically Southern group species B. megalospilus are found within B. syspilus together with some samples of the Southern group species B. labiosa and B. robusta (Supp. file 2: Fig. S3 View Fig ). These three species clearly are Southern group species whose mtDNA in these particular samples was swept by mtDNA from B. syspilus following a single postulated hybridization event from B. syspilus into the three introgressed species, dated in our phylogeny at 0.2 Ma (cf. Fig. 17 View Fig and Supp. file 2: Fig. S3 View Fig ). Non-introgressed samples of B. labiosa and B. robusta are found as separate Southern group species in all phylogenetic analyses (see Fig. 17 View Fig ). Non-introgressed samples of B. megalospilus have not been found in this study. The close phylogenetic position in mtDNA of the morphologically dissimilar yet clearly Northern group species B. syspilus and B. huallagae may potentially also be a case of B. syspilus introgression into the sampled specimen of B. huallagae . This specimen was sampled in the lowermost portion of its distribution closest to the natural distribution of B. syspilus and not from the type locality of B. huallagae much higher up the namesake river.
In our present taxon sampling the first clade within the Northern group thus has a 10 my time gap from the basal node at 13.9 Ma to the presently sampled divergence at 3.9 Ma and this may suggest the existence of yet undiscovered older diverged species in this clade, or alternatively, it suggests extinction within the clade.
The second clade within the Northern group shows a younger basal divergence at 7.9 Ma, hence also with a time gap and with similar potential implications, which is strenghtened by the isolated position of the basal-most species of this clade, B. mariae ( Fig. 17 View Fig ). Bujurquina mariae (from Colombia) is the only species so far sampled from the Orinoco basin to the north of the Amazon basin. The two remaining Peruvian species ( B. peregrinabunda and B. moriorum ) diverged at 4.6 Ma, but they are not found as closely related in the mtDNA phylogeny despite being more similar and lowland parapatric species. Instead, the lowland B. peregrinabunda is found closely related to the allopatric Andean endemic B. zamorensis from Ecuador plus one putative lowland Ecuadorian species, most likely also found in Peru (B. sp. Ecuador 13). Bujurquina moriorum is found in a clade with two other putative Peruvian species from the Marañón basin.
Phylogeny and coloration patterns: the two main groups and phylogenetic lineages of Bujurquina
Our phylogenetic results have shown that Bujurquina is divided into two main clades. One clade, based on distribution patterns referred to us as the Northern group, is characterized by ornamented dorsal fins and a midlateral stripe running generally to the dorsal margin of the caudal peduncle ( Figs 1 View Fig , 4–5 View Fig View Fig and 17 View Fig ). In contrast to this the Southern group is characterized by unpatterned (hyaline, i.e., without any markings) dorsal fins and a midlateral stripe that always runs towards the posterior insertion of the dorsal fin ( Figs 1–3 View Fig View Fig View Fig and 17 View Fig ). Lower lip coloration is another character that almost fully separates the two groups, since in majority of the Northern group species the lower lip is distinctly azure blue while in the Southern group species it is always the same color as the upper lip (i.e., never blue).
Key to the species of Bujurquina View in CoL
For the key we have chosen to use only readily visible external characters. All characters except where specifically noted are for large adult specimens (usually above 60 mm SL), especially the color and coloration pattern characters, where most are best visible in live specimens. In bold are the main distinguishing character states for each step.
1. Spinous portion of the dorsal fin ornamented (i.e., with blotches, lines etc.); midlateral stripe on body oriented towards posterior insertion of dorsal fin except in one species ( B. syspilus View in CoL ), i.e., oriented nearly horizontally; flanks with dark, squarish or wedge-shaped spots, in overlapping region between scale base and overlying scale margin except in two species ( B. mariae View in CoL and B. ortegai View in CoL ) ................ 2
– Spinous portion of the dorsal fin unpatterned (i.e., hyaline without any markings); midlateral stripe on body oriented towards soft portion of dorsal fin, i.e., oriented more dorsally, flanks without spots .................................................................................................................................................. 9
2. Spinous portion of the dorsal fin with diagonal lines (i.e., dark lines on a hyaline background) ... 3
– Spinous portion of the dorsal fin with circular markings (i.e., hyaline blotches on a dark background) ...................................................................................................................................... 6
3. Diagonal lines on spinous portion of the dorsal fin thick, sometimes barely recognisable as diagonal line but approaching blotches in shape, arranged as one longitudinal series with one line per membrane, dorsal-fin lappets orange; midlateral stripe on body oriented towards soft portion of dorsal fin; suborbital stripe vertical, in adults barely visible ....................... B. syspilus (Cope, 1872) View in CoL
– Diagonal lines on spinous portion of the dorsal fin thin .................................................................. 4
4. Three diagonal lines vertically on each membrane of spinous portion of the dorsal fin, each line spanning three consecutive membranes, lines oriented distinctly diagonal, about 45%, dorsal-fin lappets orange; suborbital stripe vertical, in adults distinct................. B. mariae (Eigenmann, 1923) View in CoL
– One to two diagonal lines per membrane on spinous portion of the dorsal fin ............................... 5
5. Lines per membrane oriented only slightly diagonal, two lines only in largest adult specimens, one line in dorsal portion of membrane in all other sizes, dorsal-fin lappets dark, with orange tip; suborbital stripe bent from vertical to posteriad in juveniles, reduced to a preopercular spot in adults; large head, length equal or above 36.3% SL; longest snout of all species without overlap (mean 15.5%, 14.4–16.4% of SL) .................................................................................. B. omagua sp. nov.
– Lines per membrane oriented distinctly diagonal, about 45%, dorsal-fin lappets nearly hyaline, only very slightly tinged orange; suborbital stripe slightly bent from vertical to posteriad, in adults well developed ......................................................................................... B. ortegai Kullander, 1986 View in CoL
6. Suborbital stripe vertical .................................................................................................................. 7
– Suborbital stripe bent from vertical to posteriad .............................................................................. 8
– Suborbital stripe inclined posteriad, in adults reduced to its ventral portion in the form of a short stripe or a preopercular spot; dorsal-fin lappets dark to black, not orange ......................................... ............................................................................................................. B. moriorum Kullander, 1986 View in CoL
7. Suborbital stripe vertical, shifted posteriorly to a position behind the eye, in adults always as a stripe, sometimes reduced only to its ventral portion; dorsal-fin lappets orange-brown, same color as rest of markings on dorsal fin, not orange ................................. B. peregrinabunda Kullander, 1986 View in CoL
– Suborbital stripe vertical, in adults whole; dorsal-fin lappets orange-brown, same color as rest of markings on dorsal fin, not orange ..................................................... B. huallagae Kullander, 1986 View in CoL
8. Suborbital stripe bent from vertical to posteriad, in adults reduced to an ocellated preopercular spot; dorsal-fin lappets orange-brown, same color as rest of markings on dorsal fin, not orange ..... .............................................................................................................. B. zamorensis (Regan, 1905) View in CoL
– Suborbital stripe bent from vertical to posteriad, in adults whole; dorsal-fin lappets white with dark base ................................................... B. pardus Arbour, Barriga Salazar & López-Fernández, 2014 View in CoL
9. Head large, length equal or above 37% SL .................................................................................... 10
– Head small, mean length below 35.5% SL ......................................................................................11
10. Mean length of head above 38.6% of SL, average 39.1%, lower pharyngeal teeth molariform, suborbital stripe in adults complete, dorsal-fin lappets white with dark base .................................... .......................................................................................................... B. oenolaemus Kullander, 1987 View in CoL
– Mean length of head above 37%, lips hypertrophied, suborbital stripe in adults incomplete, limited to below eye or to middle of cheek (does not reach angle of preopercle), dorsal-fin lappets white with dark base .................................................................................................. B. labiosa Kullander, 1986 View in CoL
11. Suborbital stripe in adults complete ............................................................................................... 12
– Suborbital stripe in adults incomplete ........................................................................................... 13
– Suborbital stripe in adults reduced to preopercular spot ............................................................... 14
– Suborbital stripe in adults absent, dorsal-fin lappets white with dark base ....................................... ............................................................................................................... B. eurhinus Kullander, 1986 View in CoL
12. Suborbital stripe in adults complete, distinct, dorsal-fin lappets dark red to brown ......................... ...................................................................................................................... B. vittata (Heckel, 1840) View in CoL
– Suborbital stripe in adults complete, indistinct, dorsal-fin lappets distinctly orange ...................... .................................................................................................................. B. robusta Kullander, 1986 View in CoL
– Suborbital stripe complete, indistinct, dorsal-fin lappets white ...... B. cordemadi Kullander, 1986 View in CoL
13. Suborbital stripe in adults incomplete, limited to below eye, weakly developed, dorsal-fin lappets red in anterior portion, black in posterior portion ........................ B. megalospilus Kullander, 1986 View in CoL
– Suborbital stripe in adults incomplete, limited to below eye, weakly developed, dorsal-fin lappets white ................................................................................................. B. tambopatae Kullander, 1986 View in CoL
14. Suborbital stripe in adults reduced to preopercular spot, well developed, in most specimens clearly visible, dorsal-fin lappets with dark tips ................................................ B. hophrys Kullander, 1986 View in CoL
– Suborbital stripe in adults reduced to preopercular spot, very weakly developed, in most specimens almost invisible, dorsal-fin lappets with dark orange tips ............. B. apoparuana Kullander, 1986 View in CoL









No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |