
Syntropis Kraepelin, 1900
|
publication ID |
https://doi.org/ 10.1206/830.1 |
|
publication LSID |
lsid:zoobank.org:pub:5793E88C-9654-41BC-BAF0-36017C1AEE95 |
|
DOI |
https://doi.org/10.5281/zenodo.4627443 |
|
persistent identifier |
https://treatment.plazi.org/id/174CE445-FFC0-2E65-089A-9D359596FC17 |
|
treatment provided by |
Felipe |
|
scientific name |
Syntropis Kraepelin, 1900 |
| status |
|
Syntropis Kraepelin, 1900 View in CoL
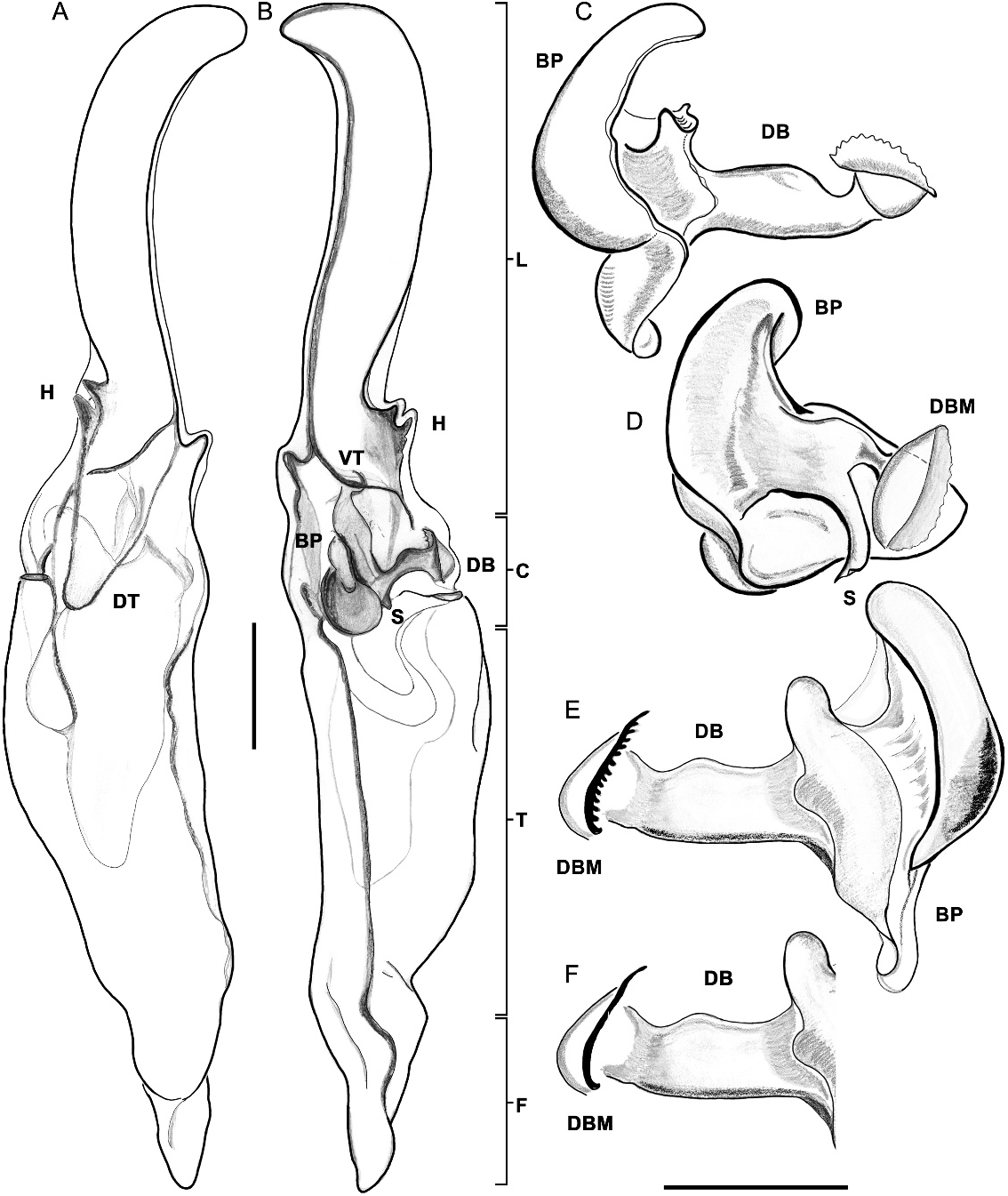
Figures 1D View Fig , 2F View Fig , 4 View Fig , 7 View Fig , 8 View Fig A–F, 12–14, 23B, 24C; table 1 View TABLE 1
Syntropis macrura Kraepelin, 1900 View in CoL , type species, by original designation.
Syntropis Kraepelin, 1900: 16 View in CoL , 17; Birula, 1917a: 163; Werner, 1934: 281; Kästner, 1941: 272; Mello-Leitão, 1945: 118; Stahnke, 1965: 257, 258; Williams, 1969: 285; 1974: 15 (part); Stahnke, 1974: 113–120; Vachon, 1974: 914, 916; Díaz Nájera, 1975: 3, 6; Williams, 1980: 47; Francke, 1985: 13, 18, 20; Sissom, 1990: 110, 114; 1991b: 26; Williams and Savary, 1991: 284; Nenilin and Fet, 1992: 9; Stockwell, 1992: 408; Kovařík, 1998: 146; Beutelspacher, 2000: 55, 70, 152, plate IId; Sissom, 2000: 526; PonceSaavedra and Beutelspacher, 2001: 20; Soleglad and Fet, 2003: 15, 36, 67, 144, 163, figs. 66, 79, 80, D-4, tables 3 View TABLE 3 , 4, 9; Fet and Soleglad, 2005: 4, 6, 7; Prendini and Wheeler, 2005: 482, table 10; Soleglad and Fet, 2005: 4, 7; Soleglad et al., 2007: 119–136, figs. 1–37, tables I, II; Soleglad and Fet, 2008: 1, 4, 13, 27, 30, 32, 38, 46, 51, 69, 71–74, 82, 84, 85, 89–92, 95, 104, figs. 10, 27, 53, 82, 106, 110, 139, 151, 180, 203, tables 1–3 View TABLE 1 View TABLE 2 View TABLE 3 , 9.
DIAGNOSIS: Syntropis differs from other Syntropinae in the carination of metasomal segments I–IV, where the vsm carinae are absent (indicated by pairs of macrosetae only, no difference in ornamentation evident from the adjacent intercarinal surfaces), and the vm carinae distinct (protruding above the adjacent intercarinal surfaces), smooth to finely granular, and unpigmented (fig. 24C). Additional characters that separate Syntropis from other genera in the subfamily are as follows. The pedipalps, legs, and metasoma are greatly elongated and slender in adult male Syntropis , although less so in females and immatures. Syntropis is the only genus in the subfamily that consistently exhibits eight prolateral denticles on the movable finger of the pedipalp chela (fig. 12B). Although Balsateres and some Thorellius species occasionally exhibit eight prolateral denticles, the basal prolateral denticle is situated adjacent to the preceding prolateral denticle, and up to three adjacent prolateral denticles are often observed (fig. 20A, B), compared with Syntropis , in which the prolateral denticles are separate. Adult male Syntropis display a whitish glandular area between the vsm carinae of mesosomal sternite VII, extending almost the entire length of the segment (fig. 23B), that is reduced but still evident in females. The dl and lm carinae of metasomal segments I–IV do not project posterolaterally in Syntropis , and the terminal granules of these carinae are not noticeably larger than the preceding granules, unlike most genera in the subfamily. Macrosetal counts on the leg basitarsi (23–28) and telotarsi (19–22) of Syntropis are greater than those on the basitarsi (14–27 in Chihuahuanus , gen. nov., and 16–21 in Kochius ) and telotarsi (14–17 in Chihuahuanus , gen. nov., and 15–18 in Kochius ) of other genera in the subfamily. Syntropis shares with Kuarapu , Vizcaino , gen. nov., and some species of Paravaejovis elongated pedipalp chela fingers and distally displaced trichobothria on the fixed finger, as a consequence of this elongation. However, the positions of trichobothria ib and it on the fixed finger, with respect to the prolateral denticles, differ among these genera as follows: ib and it are situated at PD 6 in Syntropis , whereas ib is situated at or closer to PD6 and it is situated between PD5 and PD 6 in Kuarapu , Vizcaino , gen. nov., and some species of Paravaejovis , such as P. confusus , comb. nov., and P. waeringi , comb. nov.
Syntropis shares with Konetontli , gen. nov., Kuarapu , Maaykuyak , gen. nov., Vizcaino , gen. nov., Chihuahuanus bilineatus , comb. nov., C. coahuilae , comb. nov., and Thorellius cristimanus the presence of a secondary hook on the hemispermatophore (fig. 8H), created by an extension of the axial carina of the distal lamina, that forms a pronounced bifurcation with the primary hook.
INCLUDED SPECIES: Syntropis aalbui Lowe et al. , in Soleglad et al., 2007; Syntropis macrura Kraepelin, 1900 ; Syntropis williamsi Soleglad, Lowe, and Fet, 2007 .
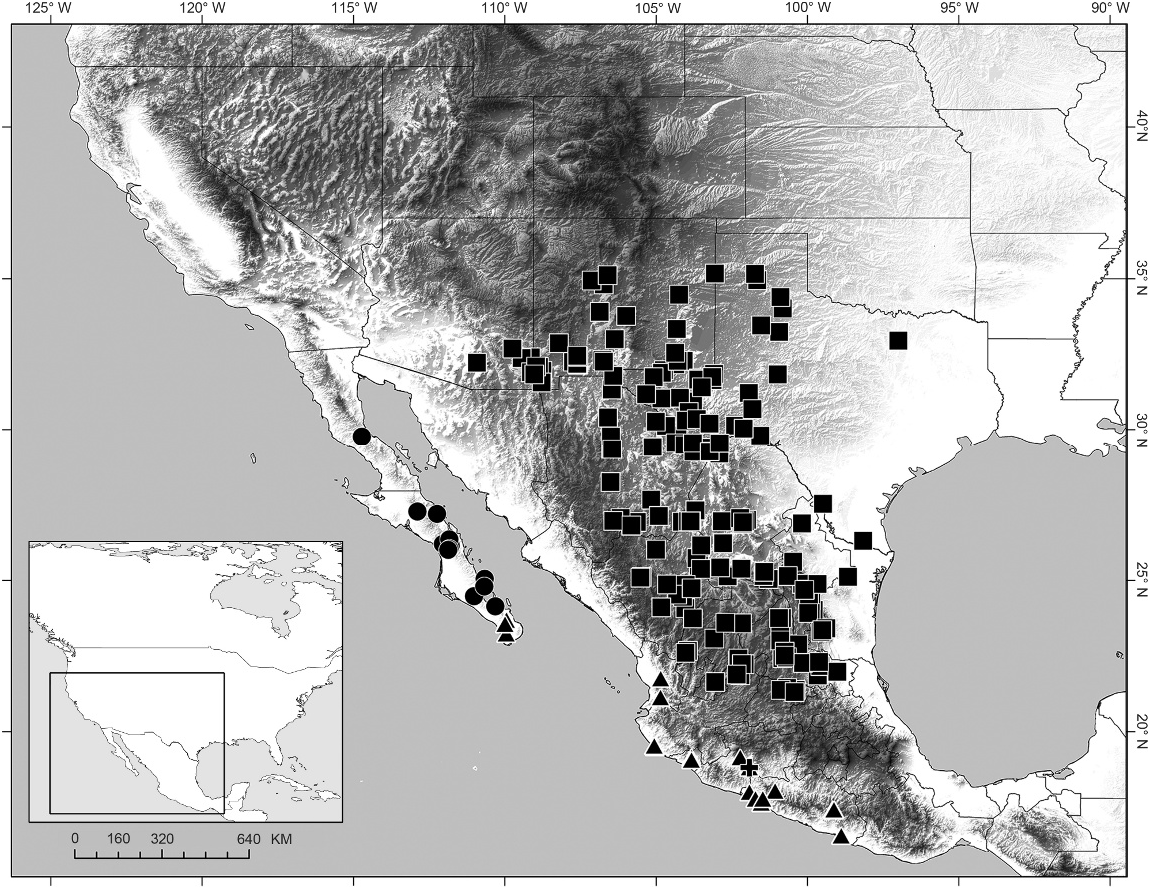
DISTRIBUTION: Syntropis is endemic to the Baja California Peninsula, Mexico, and recorded from the states of Baja California and Baja California Sur (fig. 4).
NATURAL HISTORY: Hjelle (1974) described the parturition and postparturition behavior of Syntropis . Syntropis species have been collected in rocky desert habitats, from sea level to 556 m altitude, with S. aalbui taken at the highest elevation. All three species of Syntropis are lithophilous, inhabiting the cracks and crevices of cliffs and rock walls, and hiding in loose rock piles or under exfoliating rock flakes. The habitat and habitus, particularly the slender, elongated pedipalps, legs, and metasoma, well-developed superciliary carinae of the medial ocular tubercle, and the tarsal morphology, are consistent with the lithophilous ecomorphotype ( Prendini, 2001a). Jiménez-Jiménez and Palacios-Cardiel (2010) reported capturing species in pitfall traps, indicating that these scorpions will disperse between rocky habitats. Syntropis macrura was reported to be synanthropic in the vicinity of the Comondús ( Williams, 1980; Jiménez-Jiménez and Palacios-Cardiel, 2010) and observations suggest that population densities are greater at such oases.
MATERIAL EXAMINED: Syntropis aalbui Lowe et al. , in Soleglad et al., 2007: MEXICO: Baja California: Municipio de Ensenada: Blue Palm Canyon (intermittent river La Bocana), 5 km N Cataviña, 13.vii.1979, R. Aalbu, holotype ♀ ( MHNG). Syntropis macrura Kraepelin, 1900 : MEXICO: Baja California Sur: Municipio de Comondú: San José Comondú, 27.viii.2006, C. Palacios, 18, 1♀ ( CAECIB). Syntropis williamsi Soleglad, Lowe, and Fet, 2007 : MEXICO: Baja California Sur: Municipio de La Paz: N of Los Aripes, 25.vi.1985, W. R. Lourenço and G.A. Polis, holotype ♀ ( MHNG). Municipio de La Paz: El Pilar , ca. 20 km NE Las Pocitas , 24 ° 28.7616 ′ N 111 ° 01.12 ′ W, 92 m, 25.vi.2008, H. Montaño and E. González, 18, 1♀ ( AMNH [ ARA 2825 ]). GoogleMaps



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Syntropis Kraepelin, 1900
| González-Santillán, Edmundo & Prendini, Lorenzo 2013 |
Syntropis
| Soleglad, M. E. & V. Fet 2008: 1 |
| Soleglad, M. E. & G. Lowe & V. Fet 2007: 119 |
| Soleglad, M. E. & V. Fet 2005: 4 |
| Prendini, L. & W. C. Wheeler 2005: 482 |
| Soleglad, M. E. & V. Fet 2005: 4 |
| Soleglad, M. E. & V. Fet 2003: 15 |
| Beutelspacher, C. R. 2000: 55 |
| Sissom, W. D. 2000: 526 |
| Kovarik, F. 1998: 146 |
| Nenilin, A. B. & V. Fet 1992: 9 |
| Stockwell, S. A. 1992: 408 |
| Williams, S. C. & W. E. Savary 1991: 284 |
| Sissom, W. D. 1990: 110 |
| Francke, O. F. 1985: 13 |
| Williams, S. C. 1980: 47 |
| Diaz Najera, A. 1975: 3 |
| Williams, S. C. 1974: 15 |
| Stahnke, H. L. 1974: 113 |
| Williams, S. C. 1969: 285 |
| Stahnke, H. L. 1965: 257 |
| Mello-Leitao, C. de 1945: 118 |
| Kastner, A. 1941: 272 |
| Werner, F. 1934: 281 |
| Birula, A. 1917: 163 |
| Kraepelin, K. 1900: 16 |