
Polymastia arctica ( Merejkowsky, 1878 )
|
publication ID |
https://doi.org/ 10.5281/zenodo.5393690 |
|
persistent identifier |
https://treatment.plazi.org/id/0F5C87F6-7649-F96E-FF0E-BBE4FEEAFA0E |
|
treatment provided by |
Marcus |
|
scientific name |
Polymastia arctica ( Merejkowsky, 1878 ) |
| status |
|
Polymastia arctica ( Merejkowsky, 1878) View in CoL
Rinalda arctica Merejkowsky, 1878: 4 View in CoL ; 1880: 421.
Polymastia arctica View in CoL – Vosmaer 1885: 14.
Polymastia penicillus View in CoL – Swarczewsky 1906: 7.
Polymastia mamillaris View in CoL – Arnesen 1918: 8. — Ereskovsky 1993: 22; 1995: 724. — Plotkin & Ereskovsky 1997: 127. — Plotkin et al. 1999: 18.
Polymastia mamillaris mamillaris View in CoL – Koltun 1966 (partly): 69.
TYPE MATERIAL. — White Sea. Dvina and Onega bays, 1876-1877, Merejkowsky coll., 8 syntypes ( ZIRAS). Merejkowsky (1878: 15) had not designated a holotype. Lectotype (present designation): Archipelago of Solovki , Onega Bay , 64°57’0- 65°10’8N, 35°29’4- 35°51’6E, 9-22 m, summer 1877 ( ZIRAS catalogue n° 84a, spicule slide n° 5526a, section slide n° 1); paralectotypes: Archipelago of Solovki , Onega Bay, 64°57’0- 65°10’8N, 35°29’4- 35°51’6E, 9-22 m, summer 1877, 5 specimens ( ZIRAS catalogue n° 83, 84b; spicule slides n° 103/02a-d, 5526b); Dvina Bay , off Cape Kerets, 65°25’N, 39°38’E, 11 m, 22.VI.1876, 2 specimens ( ZIRAS catalogue n° 85, spicule slides N 9112a-b). GoogleMaps
OTHER MATERIAL EXAMINED. — Specimens from ZIRAS collections.
White Sea. Kandalaksha Bay, 66°31’5N, 33°18’5E, 109 m, 22.VIII.1964, 1 specimen (as P. mamillaris mamillaris ( Koltun 1966)) (catalogue n° 10, spicule slide n° 104/02). — Onega Bay, 64°40’2N, 35°24’4E, 35 m, 22.X.1964, 1 specimen (as P. mamillaris mamillaris ( Koltun 1966)) (catalogue n° 73a, spicule slide n° 2/02a, section slide n° 2a). — Onega Bay, Archipelago of Solovki, 18 m, summer 1905, 1 specimen (as P. penicillus ( Swarczewsky 1906)) (catalogue n° 144, spicule slide n° 9114, section slide n° 3).
Barents Sea. Murman Coast, Kildin Straight, 69°18’0N, 34°10’8E, 100 m; 25.VII.1884, 2 specimens (as P. mamillaris mamillaris ( Koltun 1966)) (catalogue n° 78-79, spicule slides n° 9107 and 5603). — Chesha Bay, off Cape Veliky Nos, 67°21’5N, 47°46’5 E, 7 m, 13.X.1964, 1 specimen (as P. mamillaris mamillaris ( Koltun 1966)) (catalogue n° 145, spicule slide n° 13732). — Murman Coast, Bay of Teriberka, 69°09’0N, 35°09’0E, depth unknown, summer 1880, 4 specimens (as P. robusta ( Koltun 1966)) (catalogue n° 148, spicule slides n° 5602a-d, section slide n° 41). Personal collection of the authors:
White Sea. Kandalaksha Bay , the Keret’ Straight , 66°17’4N, 33°43’8E, 15 m, 11.VII.1998, 6 specimens. — Sredny Island , Yushkovka Harbour , 66°17’4N, 33°45’0E, 10 m, 3.VII.2000, 1 specimen. — Sredny Island, Nagovitsa Harbour, 66°17’4N, 33°38’4E, 4 m, 19. VI.1998, 1 specimen GoogleMaps ; 8-10 m, 8.VIII.1998, 12 specimens GoogleMaps ; 10 m, 28. VI.2001, 1 specimen. — Chupa Inlet , Levaya Harbour, 66°20’4N, 33°39’6E, 25.X.2000, 6 m, 2 specimens GoogleMaps ; 25 m, 9.VII.2001, 1 specimen.
Barents Sea. Murman Coast, Bay of Dal’nie Zelentsy, Nemetsky Island, 69°07’2N, 36°06’0E, 15 m, 14.VIII.2001, 1 specimen.
DESCRIPTION
External characters
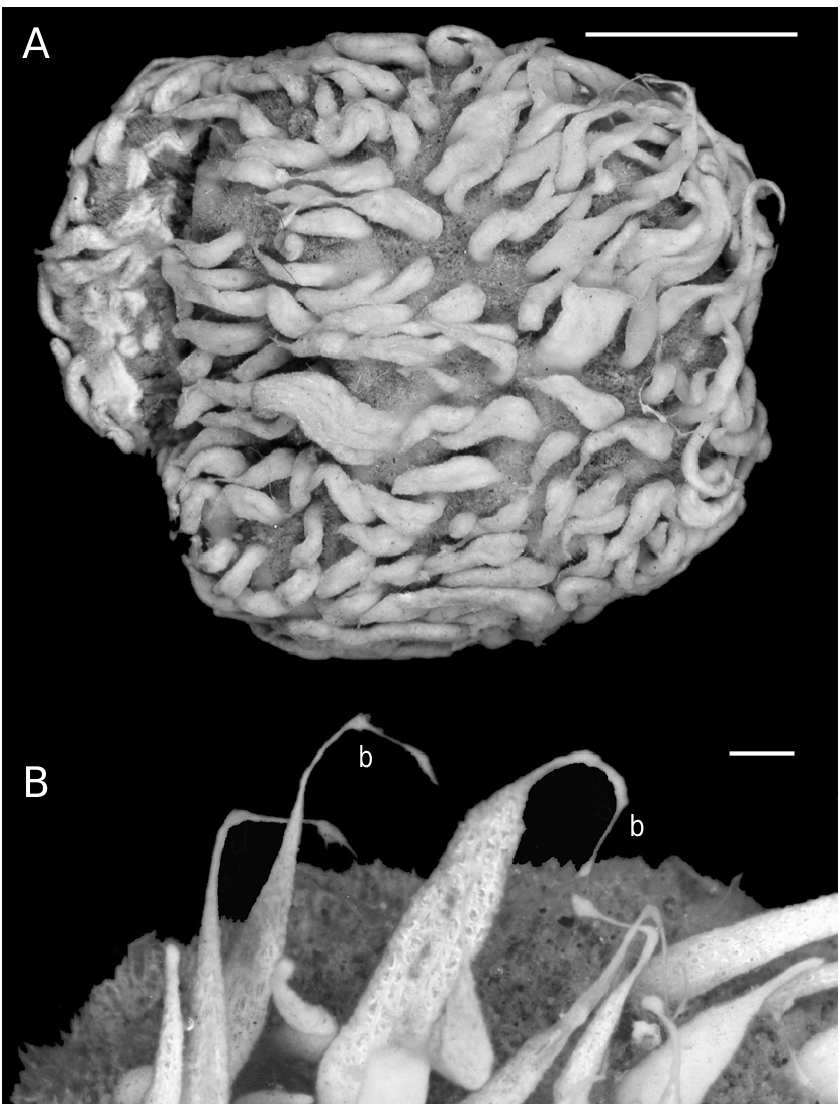
The lectotype is a cushion-shaped sponge measuring approximately 32 × 30 × 10 mm attached to a bivalve shell ( Fig. 1A View FIG ). The upper surface is hispid and partly covered with silt. The colour of the surface areas without silt and the papillae is cream in alcohol. Among other specimens of the collection, the largest ones may occupy more than 100 cm 2. The colour of the living specimens is yellow for the papillae and the surface free from silt and orange for the choanosome.
The lectotype has about 150 cylindrical inhalant and two conical exhalant papillae. The inhalant papillae are 4-9 mm in length and 1-3 mm in diameter. The exhalant papillae are 5 and 7 mm long. The diameter decreases approximately from 5 mm at the base to 3 mm at the top. The diameter of the osculum is about 0.5 mm. The number of papillae for the different studied specimens depends on body size ( Plotkin et al. 1999) and may exceed 400 inhalant and 19 exhalant papillae per specimen (mean 13 inhalant papillae per cm 2). They are respectively 2-18/ 1-6 mm and 3-12/ 3-7 mm in length. The osculum diameter is 0.2-1.0 mm. Young small sponges bear a single papilla with both exhalant and inhalant canals.
In the lectotype, about 60 inhalant papillae bear short threads with spherical buds at the top (one to three buds per papillae) ( Fig. 1B View FIG ). Of the other 42 examined specimens, 35 also possess buds at the top of the inhalant papillae. Buds are usually spherical (0.2-1.5 mm in diameter), slightly hispid and attached to a papilla by a thread formed by spicules and organic material. During the growing of buds, the thread becomes thinner. One thread may bear up to six buds arranged in line.
Skeleton
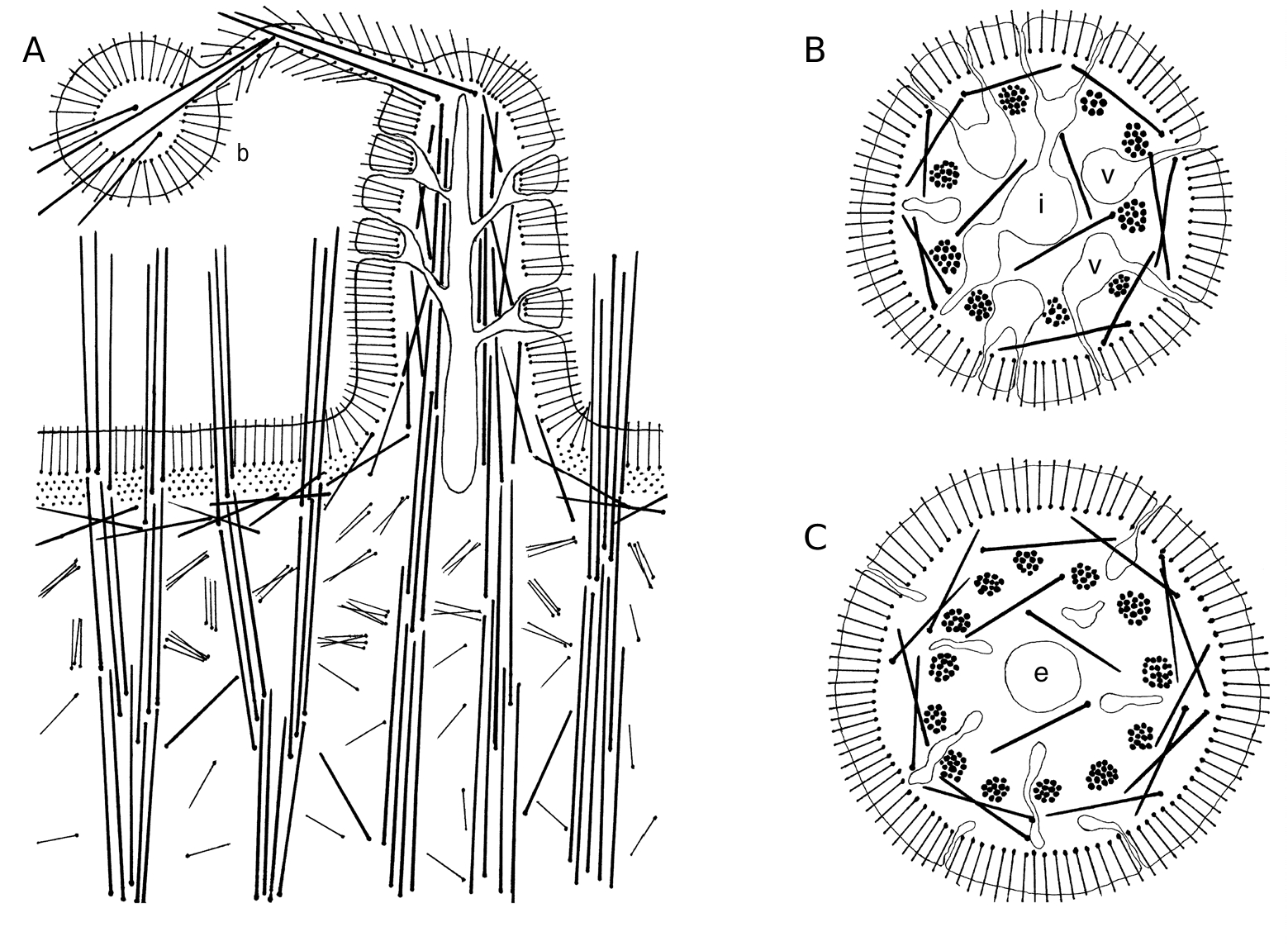
The ectosomal skeleton of the lectotype ( Fig. 2A View FIG ) is about 580 mm thick and consists of three layers: an external dense palisade layer of small tylostyles (270 µm), a middle layer of collagen (90 µm) and an internal layer of tangential intermediary spicules (220 µm). The ectosomal skeleton of the other specimens varies from 480 to 680 µm thick (mean 562 ± 47, n = 11) and the thickness of the three layers is respectively 180-310 µm (234 ± 32 µm), 90-180 µm (129 ± 19 µm) and 160- 250 µm (198 ± 18 µm).
In the lectotype the choanosomal skeleton ( Fig. 2A View FIG ) is composed of tracts of principal spicules (250-300 µm thick), which run perpendicular to the surface. They may divide below the cortex into two or three thinner ones (85-135 µm). Groups from two to five free spicules corresponding to the ectosomal tylostyles are scattered between the choanosomal tracts. They are especially abundant below the tangential layer of intermediary spicules forming a layer of about 720 µm thick. In other studied specimens the tracts of principal spicules are 170-460 µm (mean 281 ± 60 µm, n = 70 tracts) thick at the base, and after several divisions they are 70-140 µm (mean 110 ± 12 µm, n = 73 tracts) below the cortex. They cross the ectosome and echinate the surface by approximately 390- 1600 µm (mean 800 ± 254 µm, n = 11). Groups of from two to five free ectosomal spicules are scattered between the spicule tracts.They are especially abundant below the cortex forming a layer of 320- 780 µm thick (mean 558 ± 104 µm).
In the lectotype the skeleton of the papillae ( Fig. 2A View FIG ) consists of ascending tracts of principal spicules that run the length of the papillae. They are the continuation of the tracts of the choanosome. Each tract contains seven to 14 spicules. From the outside to the inside the papilla wall is
p
c t
s
composed of a palisade of ectosomal tylostyles (170-200 µm), and a tangential layer of intermediary spicules (40-60 µm). The collagenous layer presented in the main body cortex is absent in the papillae. Inhalant papillae usually possess nine tracts and a single central inhalant canal ( Fig. 2B View FIG ). The canal is connected with lateral vestibules, which open by ostioles in the wall. The internal laminae separating vestibules and the central canal contain intermediary spicules. Exhalant papillae possess 15 tracts and the central exhalant canal surrounded by several inhalant canals connected with vestibules and ostioles ( Fig. 2C View FIG ). Intermediary spicules are present in the laminae between exhalant and inhalant canals.
The number of spicules in each tract varies from six to 47 in the other studied specimens (mean 21 spicules/tract). Inhalant and exhalant papillae possess respectively seven to 11 and 11-15 tracts/ papillae. All cavities of the papillae are separated by internal laminae with intermediary spicules.
Several principal spicule tracts of an inhalant papilla may fuse at the top forming an extension, which is the central axis of a bud thread ( Fig. 2A View FIG ). The thread is echinated by ectosomal tylostyles. The bud skeleton consists of an external dense palisade of ectosomal tylostyles, a few free intermediary spicules inside and some principal spicules of the thread which cross the bud.
Spicules ( Fig. 3 View FIG )
All spicule measurements were made on 43 specimens and 10 spicules of each type were measured for each sponge.
Ectosomal spicules are small fusiform slightly bent tylostyles with well marked heads. In the lectotype they measure 150-215/5.0-6.3 µm (mean 171/5.1 µm, n = 10), the head is 5.0-7.5 µm (mean 5.6 µm, n = 10) in diameter. In the other specimens 120-215/3.8-6.3 µm (mean 161 ± 7/4.8 ± 0.2 µm). The diameter of the head is 3.8- 7.5 µm (mean 5.5 ± 0.3).
Intermediary spicules are straight or slightly bent subtylostyles 380-470/10.0-12.5 µm (mean 429/10.7 µm, n = 10) in the lectotype; 270- 550/5.0-17.5 µm (mean 414 ± 22/9.5 ± 0.8 µm) in the other specimens.
Principal spicules are fusiform straight subtylostyles: 620-960/10.0-16.3 µm (mean 803/ 13.8 µm, n = 10) in the lectotype; 620-1100/8.8- 20.0 µm (mean 868 ± 40/14.3 ± 0.8 µm) in the other specimens.

| VI |
Mykotektet, National Veterinary Institute |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Polymastia arctica ( Merejkowsky, 1878 )
| Plotkin, Alexander & Boury-Esnault, Nicole 2004 |
Polymastia mamillaris
| PLOTKIN A. S. & ERESKOVSKY A. V. & KHALAMAN V. V. 1999: 18 |
| PLOTKIN A. S. & ERESKOVSKY A. V. 1997: 127 |
| ERESKOVSKY A. V. 1995: 724 |
| ERESKOVSKY A. V. 1993: 22 |
| ARNESEN E. 1918: 8 |
Polymastia penicillus
| SWARCZEWSKY B. 1906: 7 |
Polymastia arctica
| VOSMAER G. C. J. 1885: 14 |
Rinalda arctica
| MEREJKOWSKY C. S. 1880: 421 |
| MEREJKOWSKY C. S. 1878: 4 |