
Pungitius modestus, Matsumoto & Matsuura & Hanzawa, 2021
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5005.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:3FEFF213-7140-4134-9A2A-11D83097767E |
|
persistent identifier |
https://treatment.plazi.org/id/0F500776-FFCE-791D-FF05-ACAB3D381CA0 |
|
treatment provided by |
Plazi |
|
scientific name |
Pungitius modestus |
| status |
sp. nov. |
Pungitius modestus , sp. nov.
New Japanese name: Kakure-tomiyo ( Figure 2a View FIGURE 2 ).
Pungitius sp. 3 (in part) Hosoya, 2013: 607.
Holotype. NSMT-P 133674 , 45.4 mm SL, tributary of Mogami-gawa River , Tendo, Yamagata Prefecture, Japan, 30 October 2014, collected by R. Sato and N. Hanzawa.
Paratypes. 17 specimens in total. 8 specimens, 39.7–48.4mm SL , tributary of Mogami-gawa River , Tendo, Yamagata Prefecture, Japan, NSMT-P 133670 , NSMT-P 133671 , NSMT-P 133672 , NSMT-P 133673 , 30 October 2014 ; NSMT-P 133676 , 2 November 2018; NSMT-P 136595 , NSMT-P 136596 , NSMT-P 136597 , 26 March 2020. 7 specimens, 43.6–51.7mm SL , small pond connected to Mogami-gawa River , Tendo, Yamagata Prefecture, Japan, NSMT-P 133667 , NSMT-P 133668 , NSMT-P 133669 , 22 October 2014 ; NSMT-P 133675 , NSMT-P 133677 , 7 July 2018; NSMT-P 133678 , 22 October 2014; NSMT-P 133679 , 22 October 2014. 2 specimens, 38.7–40.5mm SL , small pond connected to Mogami-gawa River , Higashine, Yamagata Prefecture, Japan, NSMT-P 140553 , NSMT-P 140554 , 29 September 2020 .
Non-type material. YAMA 3 Pi 000564, 3 specimens, 43.9–49.9 mm SL, small river connected to Mogamigawa River , Higashine, Yamagata Prefecture, Japan, 21 February 1973 ; YAMA 3 Pi 000566, 3 of 4 specimens, 38.4– 49.9 mm SL, locality same as the holotype, 18 September 1985 ; YAMA 3 Pi 000567, 1 of 2 specimens, 42.9 mm SL, tributary connected to Mogami-gawa River , Higashine, Yamagata Prefecture, Japan, 20 June 1973 ; YUMB I-1-9 , 7 specimens, 39.8–57.0 mm SL, locality same as the preceding, collection date unknown .
Comparative material. Gasterosteus platygaster Kessler, 1859 , syntypes, BMNH 1897.7 . 5.2, 2 specimens, 39.3–44.4 mm SL ; Gasterosteus sinensis Guichenot, 1869 , syntypes, MNHN-IC-0000-5228, 7 specimens, 21–26 mm SL ; Gasterosteus stenurus Kessler, 1876 , syntypes, ZIN 2471 View Materials , 5 specimens, 46–55 mm SL ; Gasterosteus bussei Warpachowski, 1888 , syntypes, ZIN 7100 View Materials , 4 specimens 42–53 mm SL ; Gasterosteus tymensis Nikolskii, 1889 , syntype, BMNH 1892.4 .28.1, 58.5 mm SL; Pygosteus kaibarae Tanaka, 1915 , ZUMT 8197 View Materials , holotype, 45.0 mm SL , Kyoto, Japan; 7 paratypes, 29.4–45.0 mm SL, ZUMT 59848–59854 View Materials , collection data same as the holotype; NSMT- P 29878, 10 non-type specimens, 35.0– 40.7 mm SL , Kyoto, Japan; YAMA 3 Pi 000592, 3 non-type specimens, 38.1–41.7 mm SL , Kyoto, Japan; NSMT-P 140555 , non-type specimens, 30.8 mm SL , Gangneung , Gangwon-do, Korea; NSMT-P 140556 , 31.8 mm SL, collection data same as the preceding; NSMT-P 140557 , 31.4 mm SL, collection data same as the preceding.
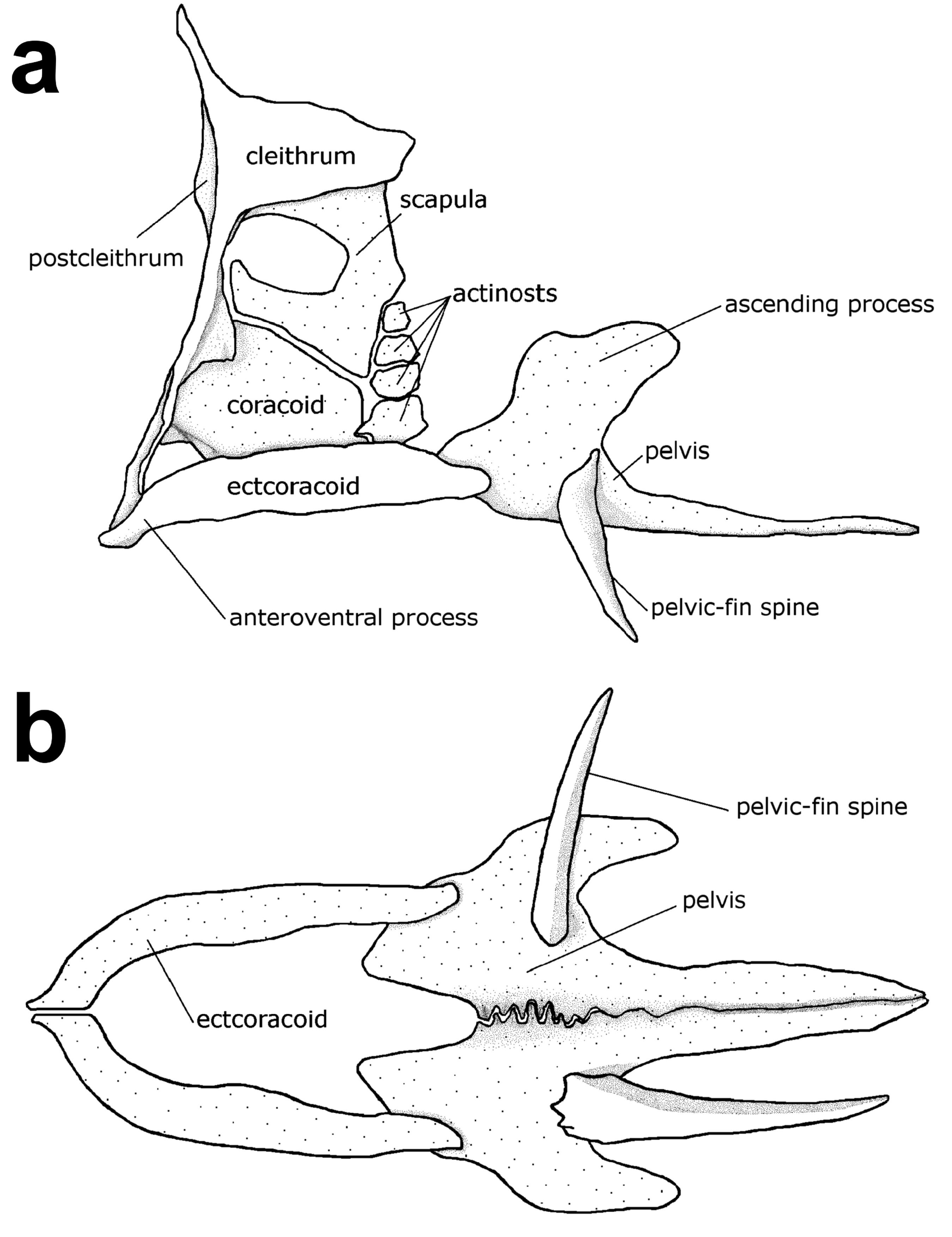
Diagnosis. Pungitius modestus is distinguished from other species of Pungitius by the following combination of characters (see also Remarks): dorsal-fin rays IX (VIII–IX)+9 (9–11); pectoral-fin rays 10 (10); 30 (30–32) small unconnected lateral plates running from just behind the dorsal end of the gill opening to the caudal peduncle ( Fig. 2b View FIGURE 2 ); keel on the caudal peduncle present; dorsal-fin spine inclining alternately to left and right of the mid-line; dorsal, pelvic and anal-fin spines short, 3.6% (2.7%–4.9%) SL, 7.4% (6.3%–9.1%) SL and 5.0% (4.1%–6.0%) SL, respectively; base of the spiny dorsal fin short, 28.7% (26.5%–29.8%) SL; first dorsal-fin spine located above or slightly behind the pectoral-fin base; anal-fin spine located below the 1st (1st–3rd) dorsal-fin ray; ascending process of the pelvis relatively developed and extending dorsally to the level of the dorsal-most actinost ( Fig. 3a View FIGURE 3 ); anteroventral process of the ectocoracoid present, right and left ectocoracoids articulating with each other at the anterior end ( Fig. 3b View FIGURE 3 ); and membranes of the dorsal-fin spines dark brown with many black pigments.
Description. Counts and morphometric measurements are shown in Tables 1–4. Body elongate and laterally compressed, tapering posteriorly to the caudal peduncle, body depth greatest just in front of the pelvic-fin origin; lateral plates on the side of the body small, oval in shape, unconnected; dorsal profile of the head almost straight, gently ascending from the mouth to dorsal-fin origin, ventral profile slightly convex; mouth small, slightly supraterminal, posterior end of the upper jaw not reaching the level of the anterior edge of the eye; eye large and rounded, diameter 6.2% SL (5.7%–7.4%), larger than the caudal peduncle depth; snout conical in lateral view, slightly shorter than the eye diameter; dorsal-fin spines strong and sharply pointed, all fin spines almost equal in length; soft dorsalfin opposite to the anal-fin, all fin rays of soft dorsal and anal-fins branched; pectoral fin fan-shaped, all rays simple; pelvic-fin composed of 1 short spine and 1 (1–2) soft ray; anal-fin composed of 1 short spine and 8 (7–9) rays; caudal-fin slightly rounded with 12 (11–13) rays composed of 10 (9–11) branched rays, and simple dorsal- and ventralmost rays; ectocoracoid located on the ventral side of the pectoral girdle, curving antero-mesially to articulate with its opposite element; anterior side of the pelvic girdle articulating with the posterior end of the ectocoracoid.
Color of fresh specimens. Dorsal and lateral sides of the head and body dark brown with a yellowish tinge, ventral side silvery white ( Figs. 2a View FIGURE 2 , 4a View FIGURE 4 ); membranes of dorsal-fin spines dark brown with many black pigments ( Fig. 2a View FIGURE 2 ); pectoral-fin hyaline; soft dorsal, anal, and caudal fins with yellowish brown rays and commonly hyaline membranes; pelvic-fin spine and membrane white; in the breeding season, male body and dorsal, anal and pelvic fins black ( Fig. 4b View FIGURE 4 ), female body light brown with numerous dark brown blotches and spots ( Fig. 4c View FIGURE 4 ).
Color in preservation. Dorsal and lateral surfaces of the head and body dark brown, ventral side light brown ( Fig. 2b View FIGURE 2 ).
Distribution. Pungitius modestus is distributed in the inland area of Yamagata Prefecture in northern Honshu, Japan. This species was recorded from the northeastern area of Yamagata Prefecture in the 1930s by Hashimoto (1938). However, the northeastern population is now thought to have been extirpated because it has not been observed in the area for several decades.
Habitat and biology. Pungitius modestus inhabits small streams and ponds refreshed by abundant cold spring water from the bottom, with temperatures of around 16°C throughout the year. Adults feed mainly on amphipods such as Jesogammarus jesoensis (identified by K. Tomikawa), which are extremely abundant in the type locality of P. modestus . In contrast, larvae of P. modestus feed on tubificids, and 30-day-old juveniles feed mainly on chironomids and copepods (Kumada et al. in preparation). Pungitius modestus tends to hide in water grasses, and also frequently remains still, suspended just above the bottom. The peak of the breeding season is May, with courtship behavior tending to occur at night, although also observed in the daytime. Females usually spawn at night, between 9 pm and dawn (T. Takeda, personal communication).
Etymology. The species name, modestus , refers to its moderate behavior: adults do not fight each other except during the reproductive season. As stated above, Pungitius modestus tends to hide in water grasses, and also frequently remains still just above the bottom. However, other species of Pungitius frequently swim in the water column ( Hart 2003, personal observation). The new Japanese name “Kakure-tomiyo” is proposed for the new species. “Kakure” implies its hiding behavior, and “tomiyo” refers to nine-spined sticklebacks.
Remarks. As stated in Introduction, Pungitius is currently represented by 11 species. Below, we describe the differences between these known species and P. modestus .
Denys et al. (2018) studied the morphological and genetic characteristics of Pungitius in France, recognizing three valid species: P. pungitius , P. laevis , and P. vulgaris . They synonymized P. breviceps ( Blanchard, 1866) and P. lotharingus ( Blanchard, 1866) with P. laevis , and P. burgundianus ( Blanchard, 1866) with P. pungitius . Based on the descriptions and data provided by Keivany & Nelson (2000), Shedko et al. (2005) and Denys et al. (2018), P. modestus is distinguished from these three valid species as follows: it is distinguished from P. pungitius by the greater number of lateral plates (30–32, vs. 0–12), smaller number of dorsal-fin spines (VIII–X [usually IX], vs. VIII–XI [usually X]), and smaller number of anal-fin rays (7–10 [usually 8], vs. 7–12 [usually 9]), the smaller diameter of the eye (21.1%–25.6%, vs. 27.3%–33.1% HL), the lack of markings on the body (vs. brownish gray blotches arranged irregularly), dark brown membranes of spiny dorsal fin (vs. hyaline), and in the breeding season, blackening of the soft dorsal and anal fins of the male (vs. hyaline).
Pungitius modestus differs from P. laevis by the larger number of lateral plates (30–32, vs. 0–4, and only on the caudal peduncle), the lack of markings on the body (vs. blackish blotches), dark brown membranes of spiny dorsal fin (vs. hyaline); and the ascending process of the pelvis, which is relatively well developed and extends dorsally to the level of the dorsal-most actinost (vs. less developed and smaller). The new species differs from P. vulgaris by the larger number of lateral plates (30–32, vs. no lateral plates), a keel on the caudal peduncle (vs. no keel), the smaller number of pectoral-fin rays (9–10 [usually 10], vs. 10–13 [usually 11]), and the smaller diameter of the eye (21.1%–25.6%, vs. 25.3%–33.9% HL).
Upon our request, James Maclaine of the BMNH kindly provided us with photographs and morphological data of syntypes of Gasterosteus platygaster ( Fig. 5 View FIGURE 5 ). In addition, we included the counts and morphometric data documented in Keivany & Nelson (2000) and Shedko et al. (2005) for comparisons of Pungitius modestus and P. platygaster . Available characteristics differentiate P. modestus from P. platygaster by its lateral plates (small and unconnected, and running from just behind the dorsal end of the gill opening to the posterior side of the caudal peduncle, vs. large connected lateral plates), a keel on the caudal peduncle (vs. no keel), the larger number of dorsal-fin rays (9–11, vs. 6–10), the first dorsal-fin spine above or behind the pectoral-fin base (vs. in front of the pectoral-fin base), and dark brown membranes of spiny dorsal fin (vs. hyaline).
Guichenot (1869) did not provide counts of fin rays in his original description of Gasterosteus sinensis ; however, Jonathan Pfliger of MNHN kindly provided us with photographs and counts of G. sinensis syntypes ( Fig. 6 View FIGURE 6 ). The morphological data of the syntypes and those given by Shedko et al. (2005) clearly differentiate Pungitius modestus from P. sinensis by its lateral plates (small and unconnected, and running from just behind the dorsal end of the gill opening to the posterior side of the caudal peduncle, vs. large connected lateral plates), dark brown membranes of spiny dorsal fin (vs. hyaline), and in the breeding season, blackening of the male soft dorsal and anal fins (vs. hyaline).
We obtained photographs and counts of syntypes of Gasterosteus stenurus through help of Anastasia Yurtseva of ZIN ( Fig. 7 View FIGURE 7 ). The morphological data clearly differentiate Pungitius modestus from P. stenurus by its lateral plates (small and unconnected, and running from just behind the dorsal end of the gill opening to the posterior side of the caudal peduncle, vs. large connected lateral plates), the smaller number of anal-fin rays (7–10 [usually 8], vs. 9–10), and the first dorsal-fin spine above or behind the pectoral-fin base (vs. in front of the pectoral-fin base).
Staff members of ZIN attempted to locate four syntypes (ZIN 7100) of Gasterosteus bussei , but unfortunately, they were missing. However, Hiroshi Takahashi of the National Fisheries University in Japan was able to provide a photograph and X-ray of the syntypes ( Fig. 8 View FIGURE 8 ) sent to him from Valentina G. Sideleva of ZIN in 2017. The available morphological data differentiate Pungitius modestus from P. bussei by the first dorsal-fin spine located above or behind the pectoral-fin base (vs. in front of the pectoral-fin base), the lateral plates (small and unconnected, and running from just behind the dorsal end of the gill opening to the posterior side of the caudal peduncle, vs. large connected lateral plates), the longer pre-dorsal (33.0%–35.8%, vs. 27.0%–29.4% SL), the shorter base of the spiny dorsal fin (26.5%–29.8%, vs. 32.1%–33.5% SL), and the shorter pelvic-fin spine (6.3%–9.1%, vs. 10.0%–11.6% SL) and the longer caudal-fin (12.1%–15.5%, vs. 10.7%–10.9% SL).
James Maclaine of BMNH kindly provided us with photographs and morphological data of syntypes of Gasterosteus tymensis ( Fig. 9 View FIGURE 9 ). This plus the characteristics of Pungitius tymensis provided in Keivany & Nelson (2000) and Shedko et al. (2005) differentiate P. modestus from P. tymensis by its lateral plates (small and unconnected, and running from just behind the dorsal end of gill opening to the posterior side of the caudal peduncle, vs. several lateral plates in the area just behind the end of the gill opening and the caudal peduncle only), the smaller number of dorsal-fin spines (VIII–X, vs. VIII–XIII), the smaller number of anal-fin rays (7–10, vs. 8–12), the ascending process of the pelvis (relatively developed and extending dorsally to the level of the dorsal-most actinost, vs. less developed and smaller).
Within the genus Pungitius , P. modestus is most similar to P. kaibarae , which was described by Tanaka (1915) based on the type specimens collected from Kyoto Prefecture ( Fig. 10a View FIGURE 10 ). Since this original description, this species has also been collected from Hyogo, Kyoto, and Osaka prefectures in Japan as well as from Korea ( Tanaka et al. 1982, Chae and Yang 1993, Hosoya 2013, Bae and Suk 2015). Although all Japanese populations were extirpated in the period from the 1930s to 1950s ( Hosoya 2018), the species is still found along the eastern side of the Korean Peninsula ( Bae & Suk 2015) ( Fig. 10b View FIGURE 10 ). However, Hosoya (2013) suggested that the Kyoto and Korean populations of P. kaibarae differ at the species level. Our examinations differentiate P. modestus from the type specimens of P. kaibarae ( Table 5) and specimens from Korea by its lateral plates (small and unconnected, and running from just behind the dorsal end of gill opening to the posterior side of the caudal peduncle, vs. large connected lateral plates) ( Fig. 11a–c View FIGURE 11 ), the location of the first dorsal-fin spine (behind, vs. in front of the pectoral-fin base) ( Fig. 12a–c View FIGURE 12 ), the larger number of dorsal-fin spines (VIII–X [usually IX], vs. VIII–IX [usually VIII]) and the larger number of anal-fin rays (7–10 [usually 8], vs. 8–9 [usually 9]), the shorter base of the spiny dorsal fin (26.5%–29.8%, vs. 28.0%–36.3% SL), the longer pre-anal fin (59.9%–67.4%, vs. 54.7%–61.9% SL) and the anal-fin spine below the 1st–3rd dorsal-fin rays (vs. below or in front of the 1st dorsal-fin ray), the shorter pelvic-fin spine (6.3%–9.1%, vs. 8.2%–13.3% SL), and dark brown membranes of spiny dorsal fin (vs. black).
Keivany et al. (1997) re-described in detail Pungitius hellenicus , which is restricted to central Greece. In addition, its color was detailed in Keivany & Nelson (2000). Pungitius modestus differs from P. hellenicus by VIII–X dorsal-fin spines inclining alternately left and right along the mid-line (vs. II–VI dorsal-fin spines vertically arranged), a keel on the caudal peduncle (vs. no keel), a lack of markings on the body (vs. dark bars or blotches on the body), dark brown membranes of spiny dorsal fin (vs. hyaline), the presence of the anteroventral process of the ectocoracoid (vs. absence of the anteroventral process of the ectocoracoid), and the presence of a pelvic girdle (vs. absence of a pelvic girdle).
Shedko et al. (2005) described Pungitius polyakovi based on the holotype and 25 paratypes collected from southeastern Sakhalin Island, Russia. Their detailed description clearly differentiates P. modestus from P. polyakovi by the larger number of lateral plates (30–32, vs. 4–6), the smaller number of dorsal-fin spines (VIII–X [usually IX], vs. VIII–XI [usually X]), smaller dorsal-fin rays (9–11, vs. 10–12), the smaller number of anal-fin rays (7–10, vs. 9–11); the longer caudal peduncle (14.3%–19.7%, vs. 7.8%–11.6% SL); the lack of markings on the body (vs. 4–8 transverse brown bars on the body); in the breeding season, blackening of the entire male body (vs. blackening of only the ventral side of the head and body); the presence of the anteroventral process of the ectocoracoid (vs. absence of the anteroventral process of the ectocoracoid); articulation of the right and left ectocoracoids at the anterior end (vs. no articulation of the right and left ectocoracoids); and the ascending process of the pelvis (extending dorsally to the level of the dorsal-most actinost, vs. less developed and smaller).










| SL |
University of Sierra Leone, Njala University College |
| R |
Departamento de Geologia, Universidad de Chile |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Pungitius modestus
| Matsumoto, Tatsuya, Matsuura, Keiichi & Hanzawa, Naoto 2021 |
Pungitius sp. 3
| Hosoya, K. 2013: 607 |