
Tyrannosaurus rex, Osborn, 1905
|
publication ID |
https://doi.org/ 10.1080/02724634.1999.10011161 |
|
DOI |
https://doi.org/10.5281/zenodo.3483138 |
|
persistent identifier |
https://treatment.plazi.org/id/0F0287A7-FFA3-FFC9-7375-FEE7FE1DF91D |
|
treatment provided by |
Jeremy |
|
scientific name |
Tyrannosaurus rex |
| status |
|
CMNH 7541 , a damaged skull with lower jaws in occlusion ( Fig. 6A-D View FIGURE 6 ), was collected from the Hell Creek Formation of Montana in 1942 and described by C. W. Gilmore (1946) in a posthumous publication. CMNH 7541 was heavily restored in plaster, and Gilmore erred in his account of sutural fusion in the skull, a misinterpretation perpetuated by later workers ( Russell, 1970; Bakker et al., 1988; Paul, 1988). In fact, there is no evidence of sutural fusion in this specimen except for fusion of the intranasal and intraparietal sutures, which is typical of Stage 1 A. libratus . Gilmore noted the similarity of the skull to the smallest specimen of A. libratus then known (AMNH 5664); thus Gilmore (1946) made CMNH 7541 the holotype of the new taxon, Gorgosaurus lancensis .
In his review of tyrannosaurids from western Canada, Russell (1970) referred ”G”. lancensis to Albertosaurus and accepted Gilmore’s interpretation of sutural fusion and its indication of relative maturity. Later, Bakker et al. (1988:17) proposed a new genus of dwarf tyrannosaurid, Nanotyrannus , for “ G.” lancensis , offering sutural fusion between the frontal and prefrontal and between the parietal and frontal as the criteria for the adult nature of the specimen.
Relative Maturity of CMNH 7541
The presence of striated cortical bone was demonstrated by Bennett (1993) and Sampson (1993) to distinguish immature, fast-growing individuals from mature specimens for pterosaurs and centrosaurine ceratopsids, respectively. Immature bone grain is lost with increase in size and development of ontogenetic characters, thus providing a crude measure of relative maturity among reptiles.
On CMNH 7541 , immature bone grain is present on the antorbital fossa of the maxilla ( Fig. 7A View FIGURE 7 ) and lacrimal, lateral surface of the vomer, dentary, surangular ( Fig. 7C View FIGURE 7 ), angular, palatine, jugal ( Fig. 7B View FIGURE 7 ), ventral process of the maxilla, quadratojugal process of the squamosal, squamosal ramus of the postorbital, rostral surface of the supraoccipital crest of the parietal, medial surface of the prearticular and splenial, caudal margin of the quadratojugal, caudal surface of the quadrate, and dorsal surface of the frontal and nasals ( Fig. 7D View FIGURE 7 ).
In addition, none of the “fusions” (frontal-frontal, frontalprefrontal, nasal-maxilla, nasal-lacrimal, nasal-prefrontal, premaxilla-maxilla, maxilla-lacrimal, maxilla-nasal) claimed by Gilmore (1946) or by Bakker et al. (1988) actually occurs in CMNH 7541 . In fact, none of the fusions enumerated above occurs in any tyrannosaurid skulls externally (pers. obs.), except suture closure around the prefrontal varies in some T. rex (Brochu, pers. comm.). Therefore, I regard CMNH 7541 as a Stage 1 specimen on the basis of cortical bone texture, a conclusion consistent with its size (skull length 572 mm). Also, CMNH 7541 exhibits the characters typical of Stage 1 specimens of A. libratus (see Appendix 1 View APPENDIX 1 View APPENDIX 1 ).
Premaxilla—In rostral view, the premaxillae are narrow and their lateral margins are concave at the base of the maxillary processes, and the alveolar region is shallow ( Fig. 8J View FIGURE 8 ).
Maxilla—The maxillae of CMNH 7541 are laterally flattened, the alveolar process is shallow, the first tooth is incisiform, and the remaining teeth are labiolingually narrow ( Fig. 6A, D View FIGURE 6 ). Also, the ventrolateral rim of the ventral jugal process is not breached by the caudalmost neurovascular sulcus of the ventral row of foramina ( Fig. 6A View FIGURE 6 ). The lateral surface does not extend caudally over the rostral margin of the antorbital fossa ( Fig. 6A View FIGURE 6 ), the promaxillary fenestra is dorsoventrally elongate and not recessed ( Fig. 6A View FIGURE 6 ), the antorbital fenestra is longer than high, and the small maxillary fenestra is midway between the rostral margins of the antorbital fossa and restored fenestra ( Fig. 6A View FIGURE 6 ).
Nasal—In CMNH 7541 , the nasals are smooth with low transverse and fine rostrocaudal ridges ( Fig. 6A, B View FIGURE 6 ). In the smallest Stage 1 specimen of A. libratus examined (TMP 86.144.1), the rugose texture typical of mature specimens is present. The condition in CMNH 7541 might represent individual, ontogenetic, or taxonomic variation.
Lacrimal—In CMNH 7541 the rostral margin of the rostroventral lamina is concave to straight and the contact of the lamina with the jugal exceeds that of the ventral ramus as in Stage 1 A. libratus ( Fig. 6A View FIGURE 6 ). Also, there is no evidence of fusion between the ventral margins of the medial and lateral processes of the rostral ramus.
Jugal—As in Stage 1 A. libratus , the maxillary ramus of CMNH 7541 is dorsoventrally shallow and tapered ( Fig. 6A View FIGURE 6 ). The jugal pneumatic recess is a rostrally-restricted slit ( Fig. 6A View FIGURE 6 ), the postorbital articular surface approaches the orbit floor ( Fig. 6A View FIGURE 6 ), the region ventral to the postorbital ramus is convex ( Fig. 6A View FIGURE 6 ), the caudal margin of the postorbital ramus is convex at midheight, and the caudal rim of the lacrimal articular surface is subvertical ( Fig. 6A View FIGURE 6 ).
Postorbital—As in Stage 1 A. libratus , the laterodorsal margin is not everted medially into the dorsotemporal fenestra ( Fig. 6A, B View FIGURE 6 ). There is no postorbital cornual process or suborbital prong ( Fig. 6A View FIGURE 6 ); the former is represented by a textured surface. Unlike A. libratus , the rostral and caudal margins of the jugal process taper rostroventrally ( Fig. 6A View FIGURE 6 ).
Prefrontal—As in Stage 1 A. libratus specimens, the prefrontal is situated at the rostrolateral margin of the frontal, bounded caudally by a triangular, tab-like process from the frontal to separate it from the lacrimal caudolaterally ( Fig. 6B View FIGURE 6 ).
Frontal—As in Stage 1 A. libratus , the lacrimal notch in CMNH 7541 is elongate and narrow in dorsal view, the prefrontal notch is rostrocaudally stout in dorsal view, and the paired frontals are longer than wide ( Fig. 6B View FIGURE 6 ). Further evidence of the immature nature of the specimen is provided by the relatively elongate orbital margin (11 mm) and the shallow dorsotemporal fossa, which has a barely discernible rostral margin ( Fig. 6B View FIGURE 6 ). Unlike Stage 1 A. libratus , the frontals appear to rise to meet each other along the midline rostral to the dorsotemporal fossa ( Fig. 6B View FIGURE 6 ); however, cracks suggest this may be an artifact of dorsoventral crushing.
Parietal—In CMNH 7541 the delicate nuchal crest is low in caudal view ( Fig. 6C View FIGURE 6 ). Its dorsal margin is rostrally everted and the laterodorsal margin is convex in frontal section ( Fig. 6A, B View FIGURE 6 ). Unlike Stage 1 A. libratus , the sagittal crest in lateral view is tall and blade-like.
Ectopterygoid—In CMNH 7541 , unlike Stage 1 A. libratus , the ectopterygoid is inflated, except for the jugal ramus ( Fig. 6D View FIGURE 6 ). As in Stage 1 A. libratus , the muscle scar on the jugal ramus is caudolaterally positioned.
Supraoccipital—As in Stage 1 A. libratus , the dorsal process of the supraoccipital of CMNH 7541 is narrow and has a horizontal dorsal border with flange-like rostrolateral edges ( Fig. 6C View FIGURE 6 ).
Basioccipital—As in Stage 1 A. libratus , the ventrolateral margins of the occipital condyle taper toward each other ventrally and the caudoventral condylar surface is flattened ( Fig. 6C View FIGURE 6 ). The ventral surface of the basituberal web is flat and arched in caudal view ( Fig. 6C View FIGURE 6 ). The median portion of the basioccipital is concave between the laminae and the bone is strongly excavated caudolaterally by the subcondylar recess ( Fig. 6C View FIGURE 6 ).
Parabasisphenoid—As in Stage 1 A. libratus , the pneumatic foramina in the basisphenoid are small and situated ventrally and the oval scar is smooth and lateroventrally oriented ( Fig. 6C, D View FIGURE 6 ).
Dentary—As in Stage 1 A. libratus the dentary of CMNH 7541 is relatively shallow in lateral view and narrow in ventral view ( Fig. 6A, D View FIGURE 6 ).
Surangular—In CMNH 7541 the surangular is shallow and the surangular shelf is horizontal ( Fig. 6C View FIGURE 6 ).
Prearticular—As in Stage 1 A. libratus the dorsal margin of the caudal ramus of the prearticular is restricted caudally ( Fig. 6D View FIGURE 6 ), the caudal ramus is shallow with parallel dorsal and ventral margins ( Fig. 6D View FIGURE 6 ), the rostral lamina is slightly expanded and its parallel margins converge to a point distally ( Fig. 6D View FIGURE 6 ), and the caudodorsal surface of the rostral lamina is smooth ( Fig. 6D View FIGURE 6 ).
CONCLUSIONS
Immaturity of CMNH 7541
The morphological structure of CMNH 7541 agrees with that of Stage 1 specimens of A. libratus and displays no mature features. The weight of evidence indicates that CMNH 7541 is juvenile. The presence of immature bone grain ( Fig. 7 View FIGURE 7 ) precludes the specimen from representing an adult pygmy tyrannosaurid.
Status of Nanotyrannus— Rozhdestvensky (1965) first suggested that CMNH 7541 might be a juvenile Tyrannosaurus rex , based on his observations of T. bataar , in which juveniles vary in “slightly different proportions than in the large specimens, as we should expect if they are of different (ontogenetic) ages” ( Rozhdestvensky, 1965:106).
Carpenter (1992) also suggested that CMNH 7541 might be an immature specimen of T. rex on the basis of its long and low snout, separate nasals and frontal, circular orbit, and rostrocaudally oval margin of the antorbital fenestra. Carpenter was uncertain of the significance of the last feature, owing to its acute rostral margin relative to that in immature specimens of T. bataar . In fact, on both sides, this region is absent and restored in plaster in CMNH 7541 . Carpenter’s suggestion concerning ontogenetically variable features, except for the nasalfrontal contact, is consistent with the observations reported here. He noted two features that united the specimen with Tyrannosaurus : considerable lateral constriction of the snout, and a dorsally broad temporal region ( Fig. 8C, D View FIGURE 8 ). However, Carpenter (1992:528) tentatively accepted Russell’s identification of CMNH 7541 as? A. lancensis .
CMNH 7541 , and specimens referred to T. rex , share the following characters: (1) nasal processes of the premaxillae tightly appressed throughout their entire length ( Fig. 8I, J View FIGURE 8 ); (2) restricted exposure of the jugal within the antorbital fenestra ( Fig. 8E, F View FIGURE 8 ); (3) antorbital fossa reaches the nasal suture caudodorsally ( Fig. 8E, F View FIGURE 8 ); (4) transversely broad jugal pneumatic recess ( Fig. 8C, D View FIGURE 8 ); (5) elongate frontal sagittal crest ( Fig. 8C, D View FIGURE 8 ); (6) strongly divergent and short basal tubers ( Fig. 8G, H View FIGURE 8 ); (7) rostroventrally-oriented caudal occipital plate ( Fig. 8A, B View FIGURE 8 ); (8) shallow subcondylar recess ( Fig. 8G, H View FIGURE 8 ); (9) rostroventrally deep basisphenoid plate and rostrocaudally-restricted basisphenoid recess ( Fig. 8A, B, G, H View FIGURE 8 ); (10) inflated ectopterygoid ( Fig. 8A, B View FIGURE 8 ); (11) strongly convex rostral plate of the surangular ( Fig. 8E, F View FIGURE 8 ); (12) transversely narrow snout and broad temporal region relative to other tyrannosaurids ( Fig. 8C, D View FIGURE 8 ) ( Carpenter, 1992); and (13) deep mandible relative to other tyrannosaurids ( Fig. 8E, F View FIGURE 8 ). Given the low number of specimens of T. bataar examined first-hand for this study, it is conceivable that some of these characters may be generic synapomorphies. Based on the close morphological similarities between the skull of CMNH 7541 and large T. rex skulls,
I thus consider Nanotyrannus lancensis Bakker et al., 1988 a subjective junior synonym of Tyrannosaurus rex Osborn, 1905 . View Cited Treatment
Ontogeny in Tyrannosaurus rex
Ontogeny in the craniofacial skeleton of Tyrannosaurus rex is characterized by a global shift from a gracile early ontogeny to a robust late ontogeny morphotype. In T. rex the rostral maxillary and dentary teeth become conical, expanding and deepening the alveolar processes of the maxilla and dentary ( Fig. 8A, C, E, F View FIGURE 8 ). The tooth row becomes rostrodorsally reoriented, and the teeth become procumbent ( Bakker et al., 1988). Also, the maxilla loses three to four teeth from the rostral end of the tooth row, because the rostral teeth undergo the greatest change. Although tooth count has been used to distinguish tyrannosaurid taxa (e.g., Bakker et al., 1988), caution is advised because counts appear to be sensitive to ontogenetic and individual variation.
A similar pattern of loss of tooth positions is present in the maxilla of A. libratus , dropping from 16 to 13 alveoli ( Table 2 View TABLE 2 ). Although one large Stage 1 specimen (USNM 12814) has a low alveolus count, this specimen is the most mature of the growth stage (Carr, in prep.). While loss of alveoli may be individually variable, it is evident that the phenomenon occurs in mature specimens ( Table 2 View TABLE 2 ). Among other archosaurs, ontogenetic tooth loss has been reported by Mook (1921), Wermuth (1953), and Iordansky (1973) in Crocodylus cataphractus, C. porosus, C. siamensis, and Tomistoma schlegelii. This indicates that ontogenetic tooth loss among Tyrannosauridae is not withXout precedent among Archosauria.
In addition, the craniofacial air sac system (sensu Witmer, 1987, 1990, 1997) had a primary role in modifying bone structure. The antorbital air sac rested within the antorbital fenestra and antorbital fossa, sending diverticula into the ectopterygoid, palatine, lacrimal, jugal, and maxilla. As maturity was attained, the diverticula invaded the bones more fully, expanding the sinuses and bones. Combined with changes induced by an enlarged dentition, late ontogeny specimens became ‘swollenfaced’ relative to their smaller, sleeker progeny. It is probable that the swollen bones had greater cross-sectional strength than the strap-like bones of smaller animals, a morphological shift that would be important for taking live prey.
Finally, pneumatic features indicate further ontogenetic change in T. rex . The Stage 4 specimens CM 9380 View Materials , LACM 23844 View Materials , and UCMP 118742 View Materials share maxillary fenestrae that are rostrodorsally deep and extend medial to the rostral margin of the antorbital fossa to a greater degree than in other specimens (e.g., AMNH 5027 View Materials , BHI 2033 , TMP 81.6.1 , MOR 555 ). In addition, the interfenestral strut is thin, and additional pneumatic foramina are present at the apex ( CMN 9380 View Materials ) or at the base ( LACM 23844 View Materials ) of the interfenestral strut.
It is evident that the distinct structural patterns of early and late ontogeny specimens of T. rex constrained their respective foraging behaviors, which in late ontogeny individuals appears specialized to grasp and hold live prey or to dismember a large carcass. Alternatively, the changes represent a reaction norm to the size of the skull, increased bite forces, and increased prey size. Nonetheless, it is not improbable that small and large animals differed in foraging strategy, consumption technique, and/ or prey type. Such size dependent differences are found in extant crocodilians ( Grenard, 1991) and monitor lizards ( Auffenberg, 1988, 1994).
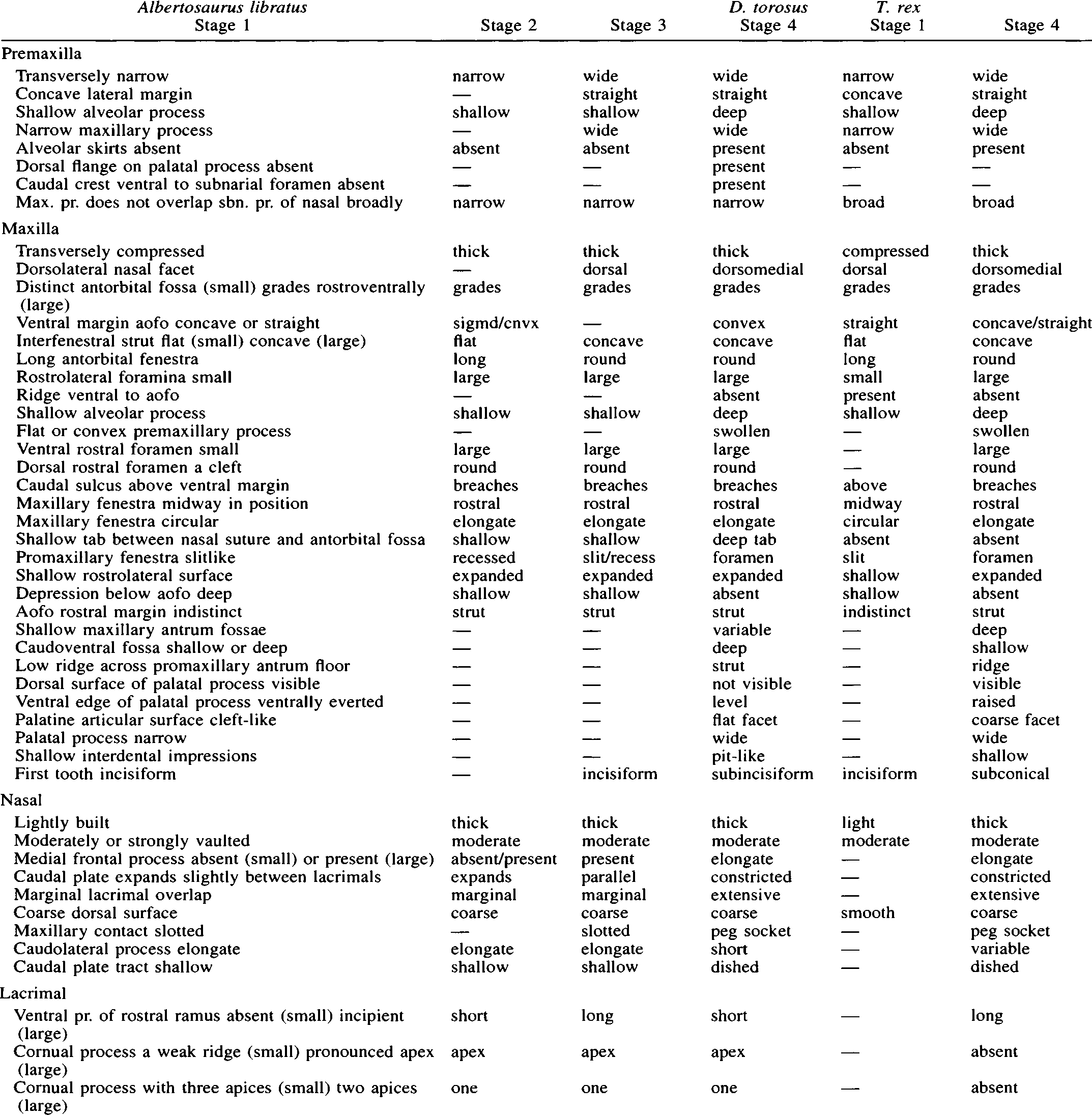



TABLE 2. Maxillary tooth counts of maxillary alveoli, growth stage, and skull lengths of Albertosaurus libratus specimens showing a tendency for more mature specimens to have fewer tooth positions. () = estimated.
| Specimen | Stage | # Maxillary alveoli | Skull length (mm) |
|---|---|---|---|
| AMNH 5664 | 1 | 15 ( Bakker et al., 1988) | 678 ( Russell, 1970) |
| ROM 1247 | 1 | 14 (pers. obs.) | (750) (pers. obs.) |
| USNM 12814 | 1 | 13 ( Bakker et al., 1988) | (795) ( Russell, 1970) |
| TMP 83.36.100 | 1 | 15 (pers. obs.) | — |
| CMN 2270 | 1 | 15 (pers. obs.) | — |
| CMN 12063 | 1 | 14 (pers. obs.) | — |
| AMNH 5336 | 2 | 13 ( Bakker et al., 1988) | (962) ( Bakker et al., 1988) |
| UA 10 | 3 | 13 (Currie, pers. comm.) | (900) (Currie, pers. comm.) |
| AMNH 5458 | 3 | 14 (pers. obs.) | 990 ( Russell, 1970) |
| CMN 2120 | 3 | 13 (pers. obs.) | 1,000 (pers. obs.) |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Tyrannosaurus rex
| Carr, T. D. 1999 |