
Ilyodrilus
|
publication ID |
https://doi.org/10.11646/zootaxa.3716.2.2 |
|
publication LSID |
lsid:zoobank.org:pub:D212CA46-26FE-4F2D-96FD-C35B58C8153D |
|
DOI |
https://doi.org/10.5281/zenodo.5676730 |
|
persistent identifier |
https://treatment.plazi.org/id/0E1287EE-FF81-FFC5-FF6C-F9FEFD0EFB98 |
|
treatment provided by |
Plazi |
|
scientific name |
Ilyodrilus |
| status |
|
Ilyodrilus (?) sp.
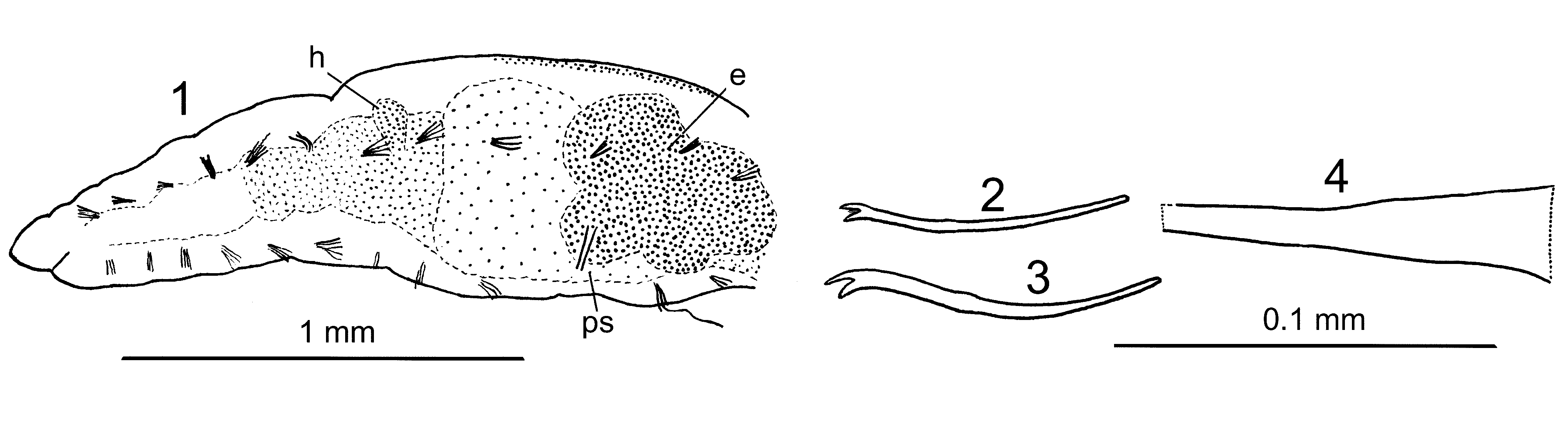
( Fig. 3 View FIGURE 3 )
Three individuals of this undetermined taxon were collected in 2011 at depths of 70 m (station 3, sexually mature individual), 100 m (station 9, maturing individual) and 135 m (station 11, maturing individual) and studied as unstained whole mounts. Length 8–16 mm, segment number about 60–80. Body wall smooth, intersegmental furrows weak, prostomium obtusely conical. Most chaetae were broken during the sampling process. In anterior dorsal bundles 1–2 thin hair chaetae (one of them 220 Μm long) and 4–5 pectinates (?), about 120 Μm long, with equal thin teeth, intermediate denticles not clearly seen. In anterior ventral bundles (3)4–6 bifid chaetae, 120–125 Μm long, with slightly longer and thinner upper tooth. No ventral chaetae in XI. In the midbody and tail regions bundles of 2–3 bifid chaetae, with upper tooth slightly longer or equal to lower. Chloragogen tissue on esophagus beginning in VI, midgut beginning in VIII; small lateral "hearts" present at the dissepiment 8/9. Clitellum in the mature individual thin, traceable in X–XII. Penis sheaths in XI as thin-walled, prolonged symmetrical truncate cones, in the mature individual measuring 160 x 40 Μm, in the maturing ones 80 x 20 or 90 x 30 Μm; the distal pore terminal. Yolk-rich eggs in XI–XII. Male ducts and spermathecae not studied.
The general shape of the penis sheaths fits the genus Ilyodrilus but the sheaths are more slender than those in the widely distributed species I. templetoni (Southern, 1909) . Tubifex montanus parvus Giani, Martínez-Ansemil & Brinkhurst, 1984 has penis sheaths very similar to those of our form; however, its anterior dorsal chaetae are almost palmate, with rough intermediate denticles (Giani et al. 1984). In the nominotypical subspecies of Tubifex montanus Kowalewski, 1919 , chaetae are more similar to those of our form while the penis sheath is depicted by Kowalewski (1919) as more funnel-shaped, with dilated proximal and tubular distal portion. Assessing the correct systematic position of our form would need a study of internal reproductive organs, as well as of DNA data. A COI barcode (not more closely accounted for here) of the sexually mature specimen from site 3 suggests that Ilyodrilus (?) sp. is a member of the subfamily Tubificinae , but it shows no close affinity to any of the barcodes of other tubificine taxa available to us (including GenBank data).
Both I. templetoni and T. montanus (the latter without subspecies name) have been repeatedly recorded from fresh waters in Turkey, although without any description: the former by Balık et al. (2004), Yıldız & Balık (2005) and Yıldız et al. (2005, 2007b, 2008, 2012); and the latter by Yıldız & Balık (2005, 2006) and Yıldız et al. (2005, 2007b, 2012). Records of another species of Ilyodrilus , I. frantzi Brinkhurst, 1965 , from the Turkish inland waters (Yıldız & Balık 2005; Yıldız et al. 2005, 2007), are dubious since it is a North American species unknown from elsewhere in the eastern hemisphere. The penis sheath of I. frantzi is depicted as much shorter than broad by Brinkhurst & Jamieson (1971).
Genetic analyses of Potamothrix . For the COI gene, the maximum pairwise distance (p-distance as well as TN93 distance) within Potamothix alatus paravanicus is 0.9 %, within P. a. hazaricus 0.8 %. Between these taxa, the p-distances are 5.3–6.2 %, the TN93 distances 5.6–6.6%. The distribution of all these distances is shown in Fig. 4 View FIGURE 4 , exposing a distinct barcoding gap. The p-distances between all Potamothrix species (with P. a. paravanicus and P. a. hazaricus together considered as one of them) vary between 13.9 and 20.5 %, and the TN93 distances between 15.6 and 24.7 %. The p and TN93 distances between the two subspecies of P. alatus and their suggested sister taxon P. bavaricus (see below and Fig. 5 View FIGURE 5 ) are 16.1–17.3 % and 18.6–20.2 %, respectively.
The ITS sequences are identical in all specimens of P. a. paravanicus and P. a. hazaricus ; otherwise this marker shows considerable variation between the taxa included in the study.
Discussion
As mentioned in the introduction, only three species of Oligochaeta have previously been recorded from the littoral of Lake Hazar: Monopylephorus irroratus , Rhynchelmis sp. and Haplotaxis sp. The first of them is probably misidentified as it is known mostly in brackish water. None of them was met in our study, which however did not consider the littoral. Instead, three tubificid taxa, all new for Lake Hazar and one of them new for science, were found in profundal. Potamothrix alatus hazaricus n. ssp. appeared to be absolutely dominating at all stations deeper than 2 m while the two other occurred very seldom.
P. a. hazaricus is rather similar to the nominotypical subspecies P. alatus alatus Finogenova, 1972 known from brackish waters of the Dnepr-Bug Estuary (Black Sea) and the Caspian Sea (Finogenova 1972, 1975). It differs from that form, apart from the freshwater habitat, in the longer upper tooth in the anterior ventral chaetae (the teeth of each chaeta being described as equally long in P. a. alatus ), the more lateral position of the spermathecal pores (in line of ventral chaetae in P. a. alatus ), and the thin, curved distal end of the spermathecal chaetae (tip of spermathecal chaetae not curved in P. a. alatus ). Finogenova (1972) and Finogenova & Poddubnaja (1990) also mention the shorter (125–170 Μm) vas deferens in P. a. alatus , in comparison with the third subspecies, P. a. paravanicus Poddubnaja & Pataridze, 1989 (150–310 Μm; about the same, 200–300 Μm, was found in the new form).
Potamothrix a. paravanicus was described from the profundal of the Transcaucasian lakes Paravani, Sagamo and Sevan, as a separate species, but which soon was attested to be a subspecies of P. alatus (Poddubnaja & Pataridze 1989; Finogenova & Poddubnaja 1990). It differs morphologically from the nominotypical subspecies and the Lake Hazar form in the complete lack of the lateral "wings" in the genital region of the body. For the individuals from Lakes Paravani and Sagamo in Georgia, Poddubnaja & Pataridze (1989) noted a large number of preclitellar ventral chaetae (6–10, seldom up to 12, per bundle) while we found much smaller numbers of them, only 2–6 per bundle, in Lake Sevan. Finogenova & Podubnaja (1990) noted the smaller number of dorsal chaetae in the Lake Sevan worms, in comparison with those from the Georgian lakes, but did not discuss it further. The upper tooth of the anterior ventral chaetae is longer in all lacustrine forms, i.e. those from Lakes Sevan, Hazar, and the Georgian lakes. Finogenova & Podubnaja (1990) described a single, thickened ventral chaeta (not observed by us) at the male pores in XI, in the Sevan material.
Spermathecal chaetae have been described as distally straight in P. a. alatus , with slightly curved distal end in P. a. paravanicus both from Lake Paravani and Lake Sevan (our new material) but with thin and much curved tip in P. a. hazaricus n. ssp.; this difference, however, may depend on a different angle of observation. The lateral position of the spermathecal pores and the structure of the male ducts are similar in P. a. paravanicus from Lake Sevan and P. a. hazaricus . In the original description of P. a. paravanicus , the third, vertical portion the of atrium was depicted as shorter and interpreted as a muscular "penial bulb" (Poddubnaja & Pataridze 1989); the latter term, however, was not repeated by Finogenova & Poddubnaja (1990). Thus, P. a. paravanicus from Lake Sevan differs from P. a. hazaricus n. ssp. morphologically in the lack of "wings" (even in the case of similar fixation with strong alcohol) and in the slightly different shape of the spermathecal chaetae, while the structure of the reproductive system is virtually identical.
The two freshwater subspecies, P. a. paravanicus (with populations in the lakes of Georgia and Armenia also being slightly different from each other) and P. a. hazaricus appear to be ecologically similar. They are abundant in the profundal of oligotrophic mountain lakes with alkaline water: 1100–3500 ind. m - 2 in Lake Sevan (Jenderedjian 1994) and 178–9067 ind. m - 2 in Lake Hazar, without the co-occurence of any other form of Potamothrix . Almost no other tubificids occur in this biotope in Lake Hazar, while the oligochaete fauna was found to be slightly more diverse in the profundal of Lake Sevan, and yet P. a. paravanicus comprised as much as 95.7% of all individuals (Jenderedjan & Poddubnaja 1987). In greater depths of Lake Sevan, the individuals of P. a. paravanicus can grow very large, up to 55 mm long (Finogenova & Poddubnaja 1990), and reach an individual age of 20 years (Jenderedjian 1994). No enlarged individuals were observed in the deepest zone of Lake Hazar, while the lakes of Paravani and Sagamo are shallower.
Species concepts and species delimitation have been issues of endless disputes in the biological literature. De Queiroz (2007) tried to alleviate the discussion by introducing his “unified species concept”, which defines species as “separately evolving metapopulation lineages”. He also pointed out that speciation is usually a gradual process, and that species delimitation therefore is a matter of finding as much evidence as possible (e.g., geographical separation, mitochondrial and nuclear gene differences, and morphological differences) that any two putative sister species indeed are evolving separately. In a more traditional approach, Mayr (1969, pp. 41–42) advocated the use of subspecies to taxonomically handle what may be regarded as incipient speciation, by recognizing subspecies as aggregates of “phenotypically similar populations of a species, inhabiting a geographic subdivision of the range of the species, and differing taxonomically from other populations of the species”; taxonomically here meaning “sufficient diagnostic morphological characters”. Mayr also noted that there may be both variation and overlap in these characters between subspecies. The subspecies category is still in common use for some vertebrate groups, but less so for most invertebrates, megadrile Oligochaeta (earthworms, Crassiclitellata) being a notable exception.
The Potamothrix alatus case has a history of subspecies designations (Poddubnaja & Pataridze 1989; Finogenova & Poddubnaja 1990), and we faced the question whether to regard the new form from Lake Hazar as yet another subspecies, and, in particular, whether we were able to assess the latter’s taxonomic affinity to P. a. paravanicus . Our genetic data clearly shows that the worms from Lake Sevan (morphologically identified as paravanicus) and Lake Hazar ( hazaricus) are closely related, and it is unfortunate that individuals of neither the nominotypical subspecies nor of P. a. paravanicus from its type locality were available to us. The COI barcoding gap ( Fig. 4 View FIGURE 4 ), associated with an approx. 6% genetic distance suggests a considerable time of separation of the populations of P. a. paravanicus and P. a. hazaricus , although these genetic differences are much smaller than those observed between other species of Potamothrix , with COI distances of 16–25% (TN93). For the nuclear ITS region, however, the time of isolation apparently has not been long enough to develop variation. Our ITS data show no conclusive evidence that paravanicus and hazaricus do not belong to the same panmictic metapopulation. Nevertheless, the geographical distribution, and the slight differences in mitochondrial and morphological characters suggest that the two forms are in a phase of at least incipient speciation, and we find it justified to regard them as separate taxonomic entities. With De Queiroz’ (2007) reasoning, we find it likely that they represent two separately evolving lineages, i.e., we could propose them to be different species. However, as we have no genetic information on P. alatus alatus , and no direct or indirect evidence of reproductive isolation between any of the forms, we have chosen the conservative alternative of assigning the Lake Hazar form to a third (new) subspecies of P. alatus . Future research will hopefully resolve the taxonomic uncertainties about this complex.
Without genetic information of P. alatus also from the brackish-water populations in the Ponto-Caspian Basin, it is difficult to judge whether the common ancestor of this complex was of euryhaline or freshwater origin. The isolation, dispersal and differentiation of the separate forms may have had a complex history, considering the geological past of the modern large Ponto-Caspian Seas. Many other Potamothrix spp. are diverse and abundant in this area, both in fresh and brackish water (Finogenova 1972, 1975), but their lack in these Transcaucasian and Turkish mountain lakes where P. alatus is present, remains an enigma. On the other hand, P. alatus has not been found in any of the many smaller Anatolian mountain and lowland lakes. Some of the latter are inhabited by a rich oligochaete fauna including several other species of Potamothrix (Yıldız & Balık 2005; Yıldız et al. 2005, 2007a, 2012; Şahin & Yıldız 2011).
Acknowledgements
The Research Foundation of Eskişehir Osmangazi University financed this study under the project No. 201119008. The first author has been supported by the target financing of the Estonian Ministry of Education and Research (No. 0170006s08). The molecular work was supported by the Swedish Research Council and the Swedish Taxonomy initiative (ArtDatabanken, SLU). We cordially thank Drs Özgür Emiroğlu and Cansev Akkan-Kökçü who kindly provided a collection of samples in Lake Hazar; Dr. Susanna Hakobyan (Institute of Hydroecology and Ichthyology, Yerevan, Armenia) for the comparative material from Lake Sevan; Annette Bergter and Dennis Kallert, for collecting other material for DNA studies; and Anna Ansebo, Per Hjelmstedt, Emelie Lindquist, Maria Lindström and Marcus Svensson, for assistance in the molecular lab. We thank Drs Enrique Martιnez-Ansemil and Patrick Martin for their thorough and useful reviews.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SubClass |
Oligochaeta |
|
Order |
|
|
Family |
|
|
Genus |