
Eideria Neita & Ocampo
|
publication ID |
https://doi.org/ 10.5281/zenodo.213516 |
|
DOI |
https://doi.org/10.5281/zenodo.6166654 |
|
persistent identifier |
https://treatment.plazi.org/id/0B3D878A-1E10-FFF9-FF73-F9C3C36BFA4F |
|
treatment provided by |
Plazi |
|
scientific name |
Eideria Neita & Ocampo |
| status |
|
Eideria Neita & Ocampo , new genus
( Figs. 1–29 View FIGURES 1, 2 View FIGURES 3, 4 View FIGURE 5 – 8 View FIGURES 9 a – b. E View FIGURES 10 – 15 View FIGURES 16 – 22 View FIGURE 23, 24 View FIGURES 25 – 28 View FIGURE 29 )
Type species. Eideria pentaphylla Neita & Ocampo , new species, here designated.
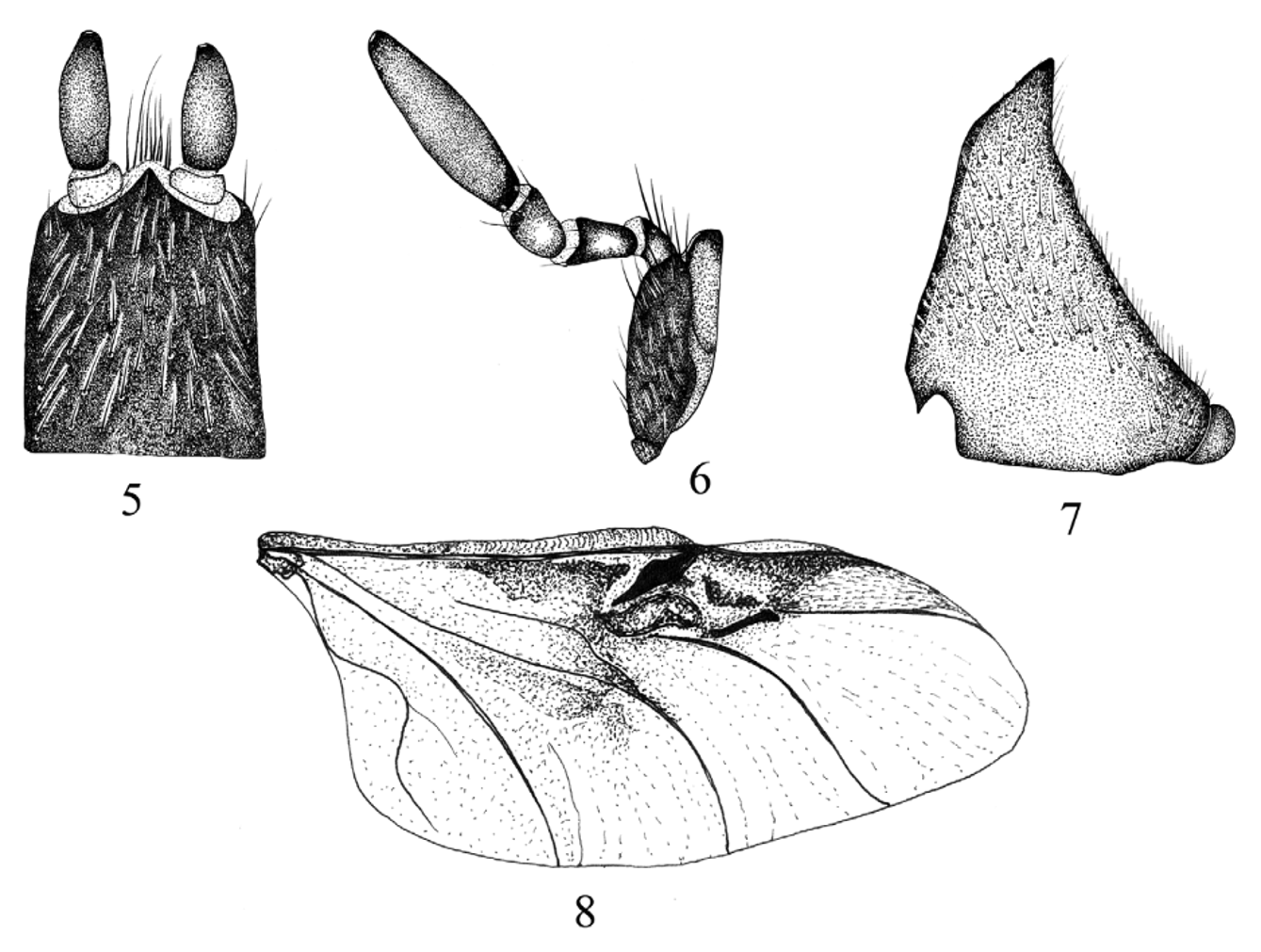
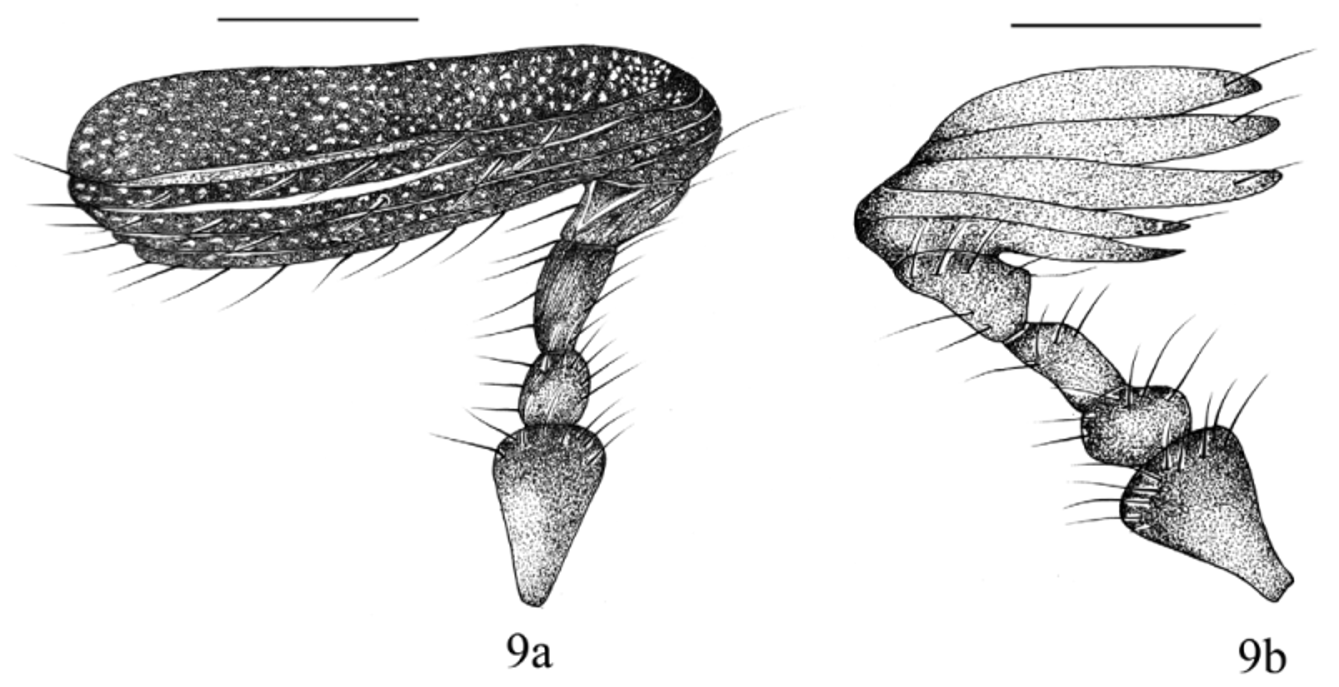
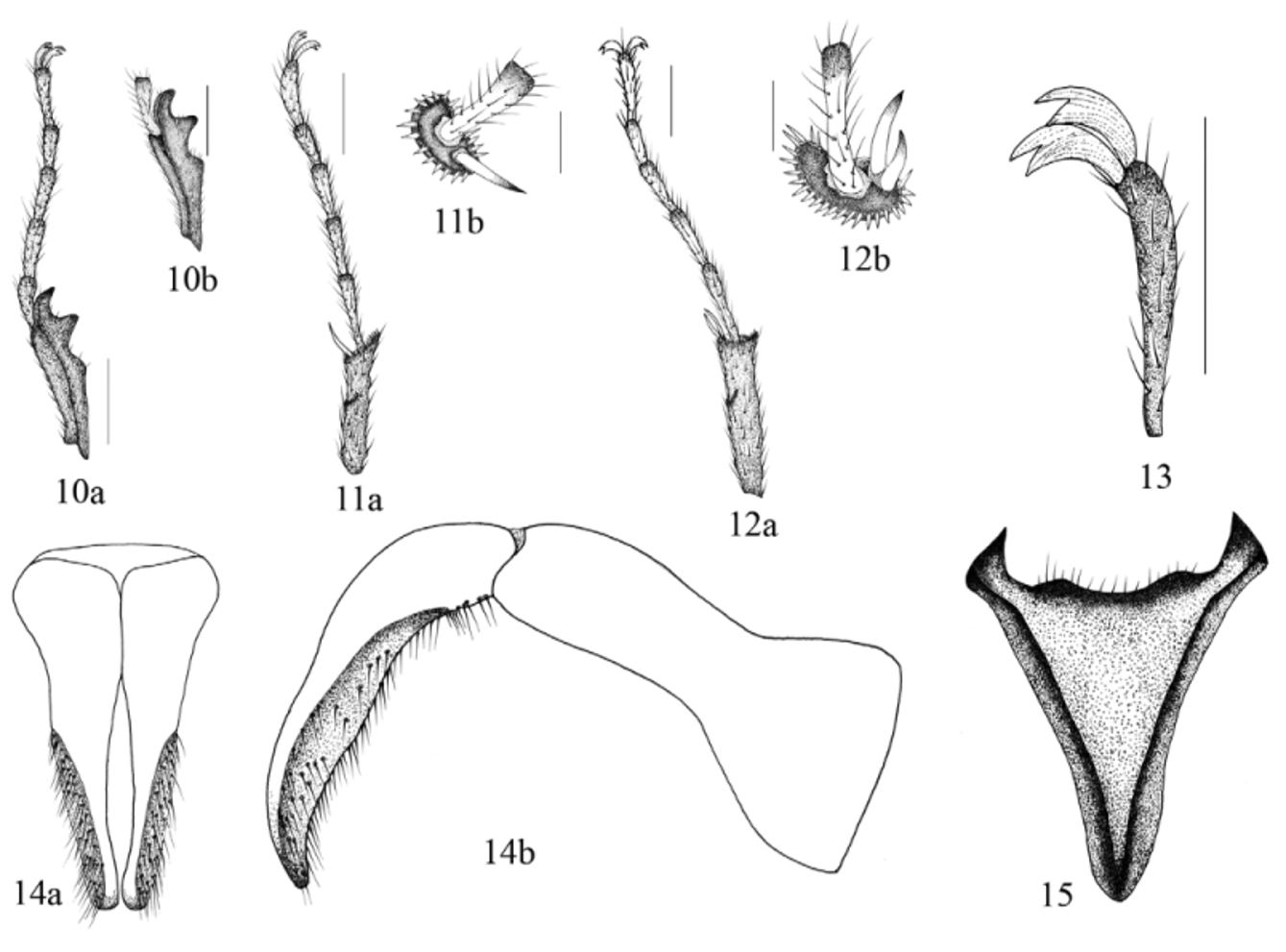
Description of male. Head ( Fig. 1 View FIGURES 1, 2 ): clypeus with apex round, margin reflexed. Labrum poorly developed, conical, not visible beyond clypeal margin (in dorsal view). Labium poorly developed, pentagonal; labial palpus with 2 palpomeres, palpomere 1 membranous, palpomere 2 sclerotized ( Fig. 5 View FIGURE 5 – 8 ). Maxilla with galea poorly developed, palpus long, with 4 palpomeres; palpomere 4 as long as 1–3 combined ( Fig. 6 View FIGURE 5 – 8 ). Mandible poorly developed ( Fig. 7 View FIGURE 5 – 8 ). Eye canthus strongly produced laterally, outer edge expanded in large, rounded projection. Antenna with 9 antennomeres, club large, subequal in length to antennomeres 2–4 combined ( Fig. 9a View FIGURES 9 a – b. E ). Pronotum ( Fig. 1 View FIGURES 1, 2 ): convex, wider than long. Margins slightly rounded, with lateral bead; bead extended to lateral half of posterior margin. Anterior angles acute; posterior angles broadly rounded. Elytra ( Fig. 1 View FIGURES 1, 2 ): convex, elongate, subparallel, completely covering dorsal surface of abdomen. Elytra with 4–8 striae. Hind wing ( Fig. 8 View FIGURE 5 – 8 ): subcostal and radial vein well sclerotized, not reaching radial sector vein at wing apex; radial sector vein forked at apex; medial vein well developed; medial loop short; cubital vein slightly sclerotized apically from medial loop connection; anal veins 1, 2 not connected, not forming a basal cell. Ve n te r: prosternal process absent, coxae transverse. Mesosternum with sulcus on middle, surface punctate, setose; punctures small, dense, ocellate. Metasternum with the apex divergent, punctures ocellate, densely setose. Abdomen with 6 ventrites exposed; ventrite V longer than ventrite I-IV combined. Pygidium wider than long, concave, recumbent toward metacoxae. Surface with punctures strongly impressed, ocellate, setose. Legs: protibia with 3 teeth ( Fig. 10 View FIGURES 10 – 15 a), protibial spur absent ( Fig. 10 View FIGURES 10 – 15 b). Mesotibia with spine-like setae on external margin, mesotibial apex with 1 spur on inner margin ( Fig. 11 View FIGURES 10 – 15 a–b). Metatibia with 2 spurs; spurs subcontiguous, on the inner edge ( Fig. 12 View FIGURES 10 – 15 a–b). Protarsomeres, mesotarsomeres, and metatarsomeres 1.85 times longer than tibiae ( Figs. 10 View FIGURES 10 – 15 a, 11a, and 12a). Tarsomeres 1 and 5 twice as long as 2–4 combined; tarsomeres 2–4 subequal; all tarsi with long, apical setae ( Figs. 10 View FIGURES 10 – 15 a–12b). Tarsal claws bifurcated, symmetrical ( Fig. 13 View FIGURES 10 – 15 ). Genitalia ( Figs.14 View FIGURES 10 – 15 a–b): parameres with a small carina on outer surface, outer surface setose; phallobase slender, with basal tegmen developed. spiculum gastrale developed ( Fig. 15 View FIGURES 10 – 15 ).
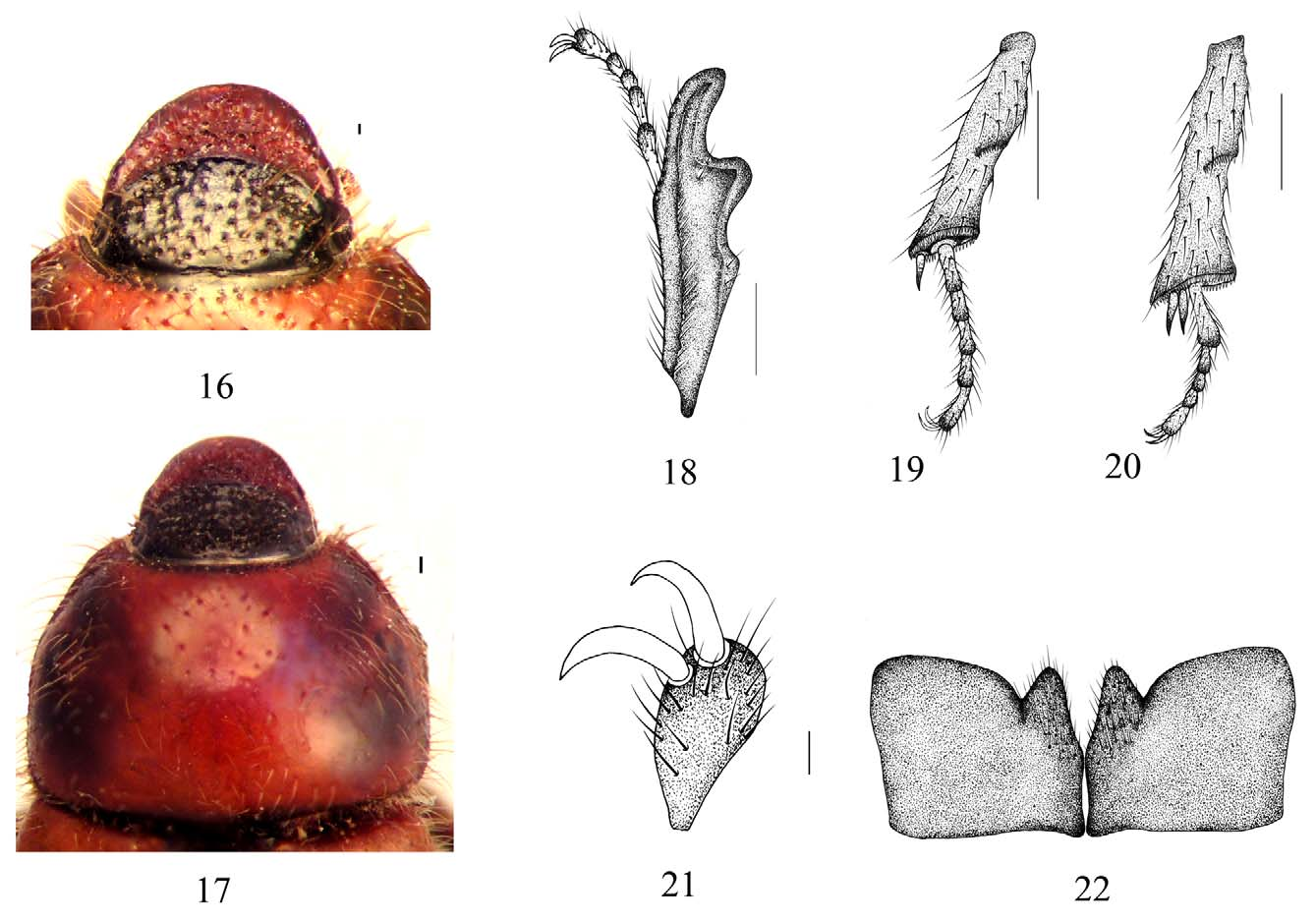
Description of female. Most New World Tanyproctini present strong sexual dimorphism. Differences can be expressed in body length and width, color, size of antennae (smaller in females), and eyes (reduced in females). Female specimens are known for the following New World Tanyproctini genera: Burmeisteriellus Berg, 1868 ; Castanochilus Ohaus, 1909 ; Lichniops Gutiérrez, 1946 ; Lichniopsoides Martínez, 1953 ; Myloxena Berg, 1881 ; Myloxenoides Martínez, 1975 , and Ptyophis Redtenbacher 1868 . Eideria females are larger than males; have lighter dorsal coloration of clypeus, pronotum, and elytra; and have reddish brown legs ( Figs. 2 View FIGURES 1, 2 , 4 View FIGURES 3, 4 ). Head: color black; punctures setose, larger than on male ( Fig. 16 View FIGURES 16 – 22 ). Clypeus slightly parabolic ( Fig. 16 View FIGURES 16 – 22 ). Eye canthus well developed, with few setae; setae long, slender. Eyes poorly developed. Frontoclypeal suture well defined, complete, convex at middle. Pronotum: convex, wider than long at middle. Surface moderately punctate, punctures ocellate, moderate in size. Anterior angles rounded; posterior angles broadly rounded ( Fig. 17 View FIGURES 16 – 22 ). Scutellum : surface opaque, punctures moderately dense. Elytra: convex, elongate. Surface shiny, setose, densely punctate. Hind wings: poorly developed, nonfunctional, reduced to a small lobe. Propygidium and pygidium not covered by elytra. Pygidium: not recumbent. Venter: prosternal process absent, slightly concave on sides. Abdomen, with 6 ventrites exposed medially, ventrite VI longer than ventrites III–V combined. Legs: protibia with 3 teeth; basal tooth small, dorsal surface with a line of setae from base to apex; protibial spur not developed ( Fig. 18 View FIGURES 16 – 22 ). Mesotibia with one spur ( Fig. 19 View FIGURES 16 – 22 ). Metatibia with two subcontiguous spurs ( Fig. 20 View FIGURES 16 – 22 ). Tarsi as long as tibiae ( Fig. 18–20 View FIGURES 16 – 22 ). Tarsal claws simple ( Fig. 21 View FIGURES 16 – 22 ).
Classification. Neotropical Tanyproctini were reviewed by Martínez (1975), who also provided a genus-level key to the tribe. Based on external morphology and male genitalia, we consider that Eideria belongs to the tribe Tanyproctini as it is currently defined.
Current classification of Melolonthinae and, in particular, the concepts currently used to diagnose Neotropical Tanyproctini and Macrodactilini, make difficult to characterize and even separate these two tribes. As a consequence of this, it is sometimes difficult to place new genera within these tribes. Katovich (2008) cited three synapomorphies for Macrodactylini : Fifth visible ventrite longer than fourth visible ventrite; propygidium lacking a complete suture between the ventrite and tergite (these characters are also found in other tribes in Melolonthinae ); and metatibiae with one or two closely placed apical spurs, inserted below the tarsal articulation, so that the metatarsus moves past them (occasionally spurs are secondarily absent). According to Katovich (2008), while none of these characters are unique, together they are unique to Macrodactylini .
According to our observations, Macrodactylini has the basal articulation condyle of the first metatarsomeres well developed, projecting above of the inner cuticle of metatibia ( Ancistrosoma Curtis, 1835 ; Barybas Blanchard, 1850 ; Ceraspis LePeletier & Serville, 1828 ; Chariodema Blanchard, 1850 ; Clavipalpus Laporte, 1832 ; Dicrania LePeletier & Serville, 1828 ; Gama Blanchard, 1850 ; Isonychus Mannerheim, 1829 ; Macrodactylus Dejean, 1821 ; and Plectris LePeletier & Serville, 1828 ). In Neotropical Tanyproctini , the condyle is poorly developed. Macrodactylini species also have the metatibae with two adjacent spurs (or no spurs), a characters commonly used to diagnose this tribe. Some Tanyproctini genera also have two adjacent spurs on the external edge of metatibia (some species in Acylochilus Ohaus, 1909 ; Leuretra Erichson, 1847 ; Myloxena Berg, 1881 ; Myloxenoides Martínez, 1975 ; and Puelchesia Ocampo & Smith, 2006 ), while other Tanyproctini genera such as; Anahi Martínez, 1958 ; Luispenaia Martínez, 1972 ; and Burmeisteriellus Berg, 1898 have the inner spur in the metatarsal apical notch and the outer spur on the external edge of metatibia; or Diaphylla Erichson, 1847 without spurs in the metatibia. The pygidium is strongly recumbent in Neotropical Tanyproctini and not strongly recumbent in Macrodactylini genera.
Sexual dimorphism in Macrodactylini is less evident than in most Neotropical Tanyproctini . Macrodactylini sexual dimorphism is generally expressed in the length and shape of antennal club, the form of the pygidium, and sometimes coloration, but these characters never reach the extreme morphological differences found in most Neotropical Tanyprocyini species. Sexual dimorphism in Tanyproctini is strongly expressed in characters such as body length and width, color, size of antennae (much smaller in females), eyes (reduced in females), and hind wings (reduced and nonfuctional in females).
Based on the above, we feel more confident placing Eideria within the tribe Tanyproctini .
Eideria shares with other New World Tanyproctini genera the following character states: body elongate, slen- der, elytral margins subparallel; clypeus broadly rounded, recumbent; frontoclypeal suture developed and evident; mouthparts (labrum, mandibles, maxillae, labium) strongly reduced; pronotum convex, wider than long; legs long slender; protibia with three teeth; mesotibiae and metatibiae with transverse carinae developed. Six ventrites exposed medially; V ventrite longer than ventrites I–IV combined; males with pygidium strongly recumbent; male genitalia symmetrical, parameres slender.
Diagnosis. Males of the genus Eideria are distinguished from all other Neotropical Melolonthinae by the following combination of characters: clypeus broadly rounded; labrum reduced, conical, not visible beyond clypeal margin (in dorsal view); antennae with 9 antennomeres, antennal club with 4–5 antennomeres ( Fig. 9a View FIGURES 9 a – b. E ); venter lacking prosternal process; metaesternum with apex divergent; pygidium recumbent toward metacoxae; protibial spur absent, mesotibia with 1 spur, metatibia with 2 spurs ( Figs.11 View FIGURES 10 – 15 a–12b); spurs subcontiguous, both set below tarsal articulation; protarsomeres, mesotarsomeres, and metatarsomeres 1 and 5 subequal in length (1.8 times longer than wide) ( Figs. 10 View FIGURES 10 – 15 a, 11a, 12a).
Eideria is similar to Luispenaia and Puelchesia . Luispenaia has a prosternal process, but this is absent in Puelchesia and Eideria . Luispenaia has tubercles on dorsal tibial surface, but this surface is smooth in Puelchesia and Eideria . Luispenaia and Puelchesia have tibiae with spurs, one on protibia and two on mesotibia and metatibiae, while Eideria has no spur on protibia, one spur on mesotibia, and two spurs on metatibia. Puelchesia and Eideria have two adjacent spurs on the on the apex of the metatibia; Luispenaia has two separated spurs, one spur on the lateral margin and one within the apical notch of the metatibia (the notch that allows mobility of the tarsus back and forth). Tarsal claws are simple in Puelchesia , but they are bifurcated in Luispenaia and Eideria . The apex of metasternum is bifurcated in Luispenaia , and simple in Puelchesia and Eideria . Puelchesia has the hind wing with anal veins V1 and V2 connected and forming a basal cell, in Luispenaia and Eideria anal veins V1 and V2 are not connected and the basal cell is not present ( Fig. 8 View FIGURE 5 – 8 ).
Etymology. Eideria is feminine in gender. We are pleased to name this genus after our friend and colleague Eider Ruiz-Manzanos, who unexpectedly passed away at a young age and who devoted her last years of research to the study of Neotropical Tanyproctini .




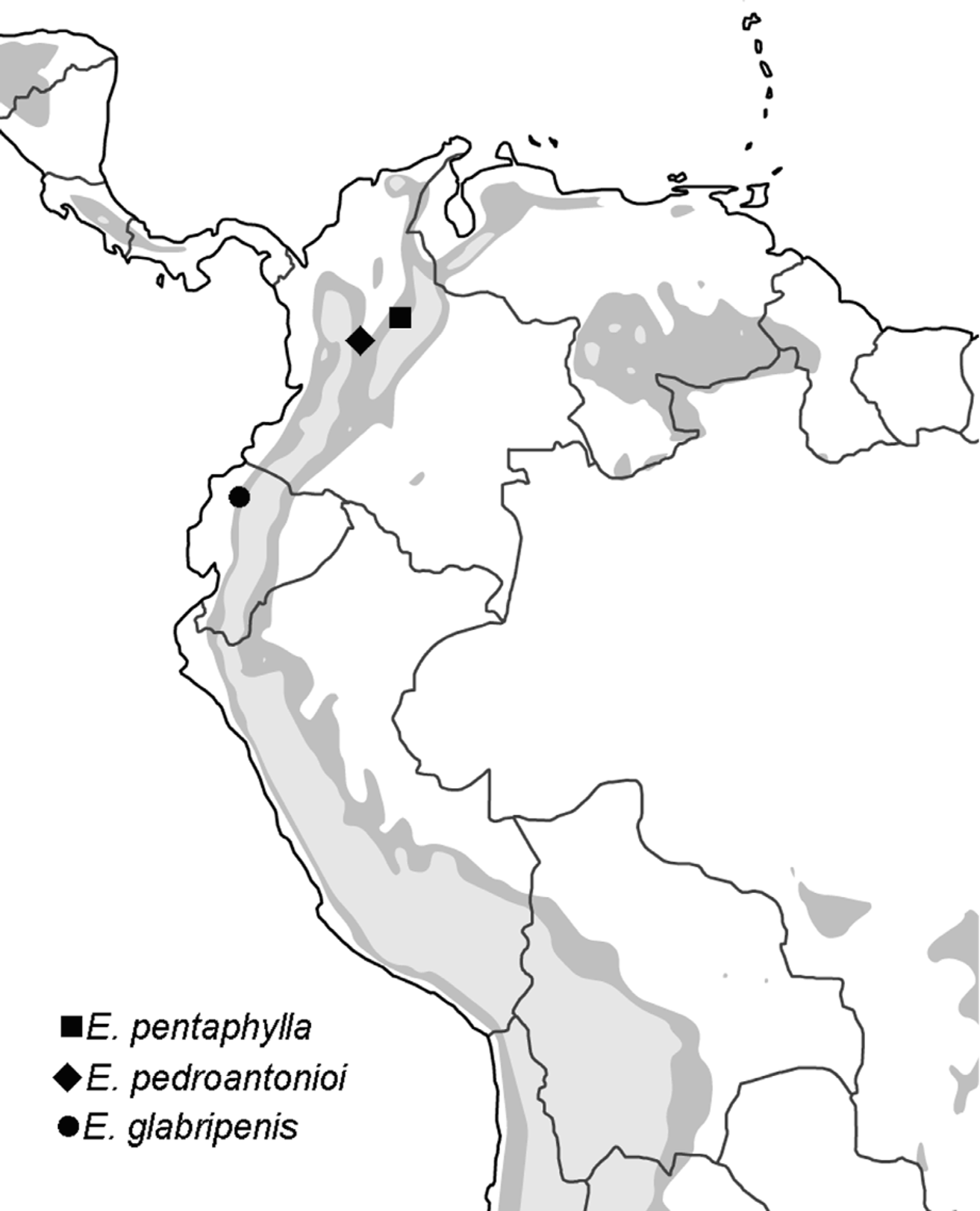
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Melolonthinae |