
Corydoras knaacki, Tencatt & Evers, 2016
|
publication ID |
https://doi.org/ 10.1590/1982-0224-20150019 |
|
publication LSID |
lsid:zoobank.org:pub:7825C540-0469-4B46-BF72-FAEB7EB11026 |
|
persistent identifier |
https://treatment.plazi.org/id/F1084581-5CC0-4CED-AD58-652BB7616F0E |
|
taxon LSID |
lsid:zoobank.org:act:F1084581-5CC0-4CED-AD58-652BB7616F0E |
|
treatment provided by |
Carolina |
|
scientific name |
Corydoras knaacki |
| status |
sp. nov. |
Corydoras knaacki , new species
urn:lsid:zoobank.org:act:F1084581-5CC0-4CED-AD58-652BB7616F0E
( Figs. 1-5 View Fig View Fig View Fig View Fig View Fig , Table 1)
Holotype. MUSM 52730 35.6 mm SL, Peru, Madre de Dios, Santa Rita, swamps in the vicinity of the town of Santa Rita, draining into the río Inambari , río Madre de Dios basin, 12°54’46.4”S 70°10’37.4”W, 19 Aug 2014, H.-G. Evers. GoogleMaps
Paratypes. All from Peru, Madre de Dios Department , río Madre de Dios basin . INPA 46982 View Materials , 4 View Materials , 24.5 View Materials -31.0 mm SL ; MNRJ 42888 View Materials , 1 View Materials , 24.6 mm SL ; MNRJ 42889 View Materials , 4 View Materials , 17.2-27.2 mm SL ; MZUSP 117378 View Materials , 4 View Materials , 20.7-28.4 mm SL; NUP 17307, 6, 22.1-36.9 mm SL; NUP 17308, 3 c&s, 34.0- 36.9 mm SL ; ZUFMS-PIS 4035 , 7 , 20.7-34.7 mm SL, collected with the holotype . ROM 82925, 1 About ROM , 37.0 mm SL, Martin Pescador stream, 11°49’60”S 71°20’60”W, 1 Oct 1991, H. Ortega & F. Zambrano GoogleMaps . ROM 82929, 12 About ROM , 29.8-39.6 mm SL, Martin Pescador stream, 11°49’60”S 71°20’60”W, 14 Oct 1991, H. Ortega & M.E. Guevara GoogleMaps . ROM 82932, 4 About ROM , 31.5-38.2 mm SL, stream tributary to the río Manu , Oct 1991, H. Ortega & M.E. Guevara .
Diagnosis. Corydoras knaacki can be distinguish from its congeners, with exception of C. acrensis , C. baderi , C. bifasciatus Nijssen, 1972 , C. boesemani , C. bondi , C. coppenamensis , C. gomezi , C. habrosus , C. haraldschultzi , C. incolicana Burgess, 1993 , C. isbrueckeri , C. julii , C. leopardus , C. nattereri , C. noelkempffi , C. ornatus , C. orphnopterus , C. pinheiroi Dinkelmeyer, 1995 , C. pulcher , C. robineae Burgess, 1983 , C. robustus , C. sipaliwini , C. sterbai and C. trilineatus , by the presence of a longitudinal black stripe along midline of flank (vs. midline of flank unspotted; with spots; not forming a conspicuous stripe); from C. bifasciatus , C. gomezi , C. haraldschultzi , C. incolicana , C. isbrueckeri , C. leopardus , C. noelkempffi , C. ornatus , C. orphnopterus , C. pinheiroi , C. pulcher , C. robineae , and C. robustus by the presence of a short mesethmoid, with anterior portion smaller than 50% of the bone length (vs. long, equal or larger than 50% of the bone length); and serrations on posterior margin of the pectoral spine directed towards the tip of the spine (vs. directed towards the origin of the spine); from C. acrensis , C. baderi , C. habrosus , C. julii , C. nattereri , C. sterbai and C. trilineatus by the presence of black pigmentation on the first branched ray, including membrane, the remaining areas with irregular black blotches (vs. anterodorsal portion of dorsal fin with a large black blotch, the remaining areas with irregular black spots in C. acrensis , C. julii and C. trilineatus ; dorsal fin entirely hyaline in C. baderi ; dorsal fin covered by black spots in C. habrosus and C. sterbai ; dorsal fin brownish, without blotches; membranes covered by black chromatophores, more concentrated on its anteriormost portion in C. nattereri ). Additionally, C. knaacki can be distinguished from C. boesemani by the absence of a vertically elongated black blotch across the eyes (vs. presence); from C. bondi and C. sipaliwini by the presence of conspicuously rounded moderately-developed black spots on the snout (vs. irregular small black spots; or scattered black chromatophores, in C. bondi ; larger irregular black spots in C. sipaliwini ); from C. coppenamensis by the presence of ventral expansion of infraorbital 1 moderately developed (vs. well developed).
Description. Morphometric data presented in Table 1. Head compressed with convex dorsal profile; triangular in dorsal view. Snout short and markedly rounded. Head profile convex from tip of snout to anterior nares; ascending, smoothly convex from this point to tip of posterior process of parieto-supraoccipital; region of posterior process of parieto-supraoccipital nearly straight; slightly convex in some specimens. Profile slightly convex along dorsal-fin base. Postdorsal-fin body profile nearly straight to adiposefin spine; markedly concave from this point to caudal-fin base. Ventral profile of body slightly convex from isthmus to pelvic girdle. Profile nearly straight from pelvic girdle to base of first anal-fin ray; abruptly concave to caudal-fin base. Body roughly elliptical in cross section at pectoral girdle, gradually becoming more compressed toward caudal fin.
Eye rounded, located dorso-laterally on head; orbit delimited dorsally by lateral ethmoid, frontal and sphenotic, ventrally by infraorbitals. Anterior and posterior nares close to each other, only separated by flap of skin. Anterior naris tubular. Posterior naris close to anterodorsal margin of orbit, separated from it by distance equal to diameter of naris. Mouth small, subterminal, width nearly equal to bony orbit diameter. Maxillary barbel moderate in size, not reaching anteroventral limit of gill opening. Outer mental barbel slightly larger than maxillary barbel. Inner mental barbel fleshy, with base close to its counterpart. Small rounded papillae covering entire surface of all barbels, upper and lower lips, and isthmus.
Mesethmoid short; anterior tip poorly developed, smaller than 50% of bone length (see Britto, 2003: 123, character 1, state 1; fig. 1B); posterior portion relatively narrow, partially exposed and bearing minute odontodes. Nasal slender, curved laterally, with inner margin laminar; mesial border contacting frontal and mesethmoid. Frontal elongated, narrow, with width slightly smaller than half of entire length; anterior projection short, size smaller than nasal length. Frontal fontanel large, oval; posterior tip extension slightly entering anterior margin of parietosupraoccipital. Parieto-supraoccipital wide, posterior process long and contacting nuchal plate; region of contact between posterior process and nuchal plate covered by thick layer skin.
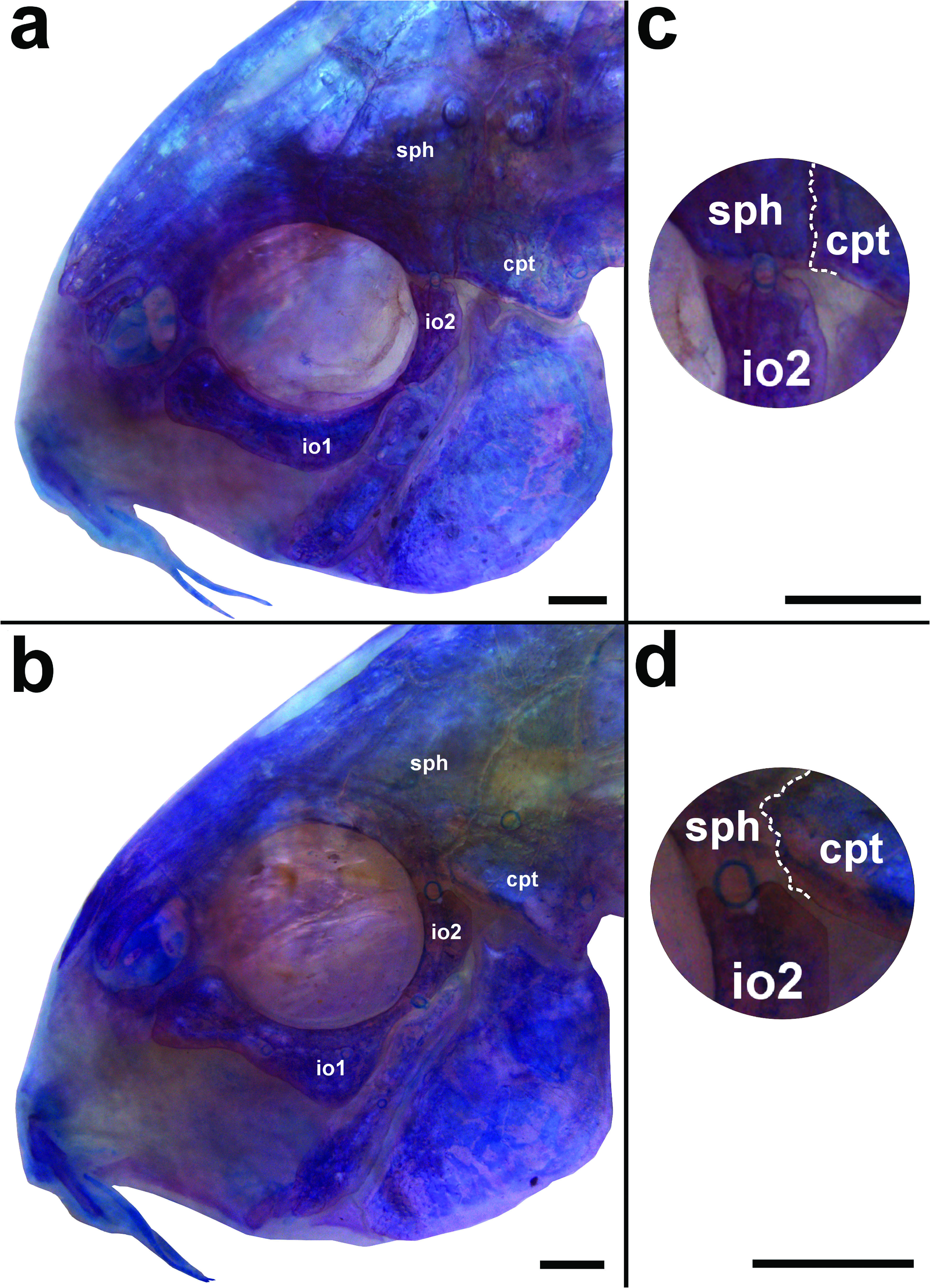
Two laminar infraorbitals with minute odontodes; infraorbital 1 large, ventral laminar expansion moderately developed; anterior portion with moderately-developed expansion ( Fig. 2a, b View Fig ); infraorbital 2 small, slender; with posterior laminar expansion moderately developed; posteroventral margin contacting posterodorsal ridge of hyomandibula, dorsal tip contacting only sphenotic ( Fig. 2c View Fig ); contacting sphenotic and compound pterotic in some specimens ( Fig. 2d View Fig ). Posterodorsal ridge of hyomandibula close to its articulation with opercle oblong; exposed, slightly thickened and bearing small odontodes; dorsal ridge of hyomandibula between compound pterotic and opercle entirely or almost entirely covered by a thick layer of skin. Interopercle entirely covered by thick layer of skin, somewhat triangular, anterior projection well-developed. Preopercle slender, elongated, minute odontodes sparse on external surface. Opercle dorso-ventrally elongated, width nearly equal to half of its length; free margin convex, without serrations and covered by small odontodes. Anteroventral portion of cleithrum and posterolateral portion of scapulocoracoid exposed; minute odontodes sparse on exposed areas. Vertebral count 21 (3); ribs 6 (3), first pair conspicuously large; complex vertebra compact in shape. Neural and haemal spines with expanded in distal tips.
Four branchiostegal rays decreasing in size posteriorly. Hypobranchial 2 somewhat triangular, tip ossified and directed towards anterior portion, posterior margin cartilaginous; ossified portion well developed, about twice size of cartilaginous portion. Five ceratobranchials with expansions increasing posteriorly; ceratobranchial 1 with small process on anterior margin of mesial portion; ceratobranchial 3 with continuous postero-lateral margin; ceratobranchial 5 toothed on postero-dorsal surface, 36 to 45 (3) teeth aligned in one row. Four epibranchials with similar size; epibranchial 2 slightly larger than others, with small pointed process on laminar expansion of posterior margin; epibranchial 3 with curved mesially uncinate process on laminar expansion of posterior margin. Two wide pharyngobranchials (3 and 4), pharyngobranchial 3 with irregular laminar exapansion on posterior margin. Upper tooth plate oval; 38 to 43 (3) teeth aligned in two rows on postero-ventral surface.
Lateral-line canal entering neurocranium through compound pterotic, splitting into two branches before entering sphenotic: pterotic branch with a single pore; preoperculomandibular branch conspicuously reduced, with a single pore opening close to postotic main canal. Sensory canal continuing through compound pterotic, entering sphenotic as temporal canal, which splits into two branches: one branch giving rise to infraorbital canal, other branch entering frontal through supraorbital canal, both with a single pore. Supraorbital canal not branched, running through nasal bone. Epiphyseal pore opening at supraorbital main canal. Nasal canal with three openings, first on posterior edge, second on posterolateral portion and third on anterior edge. Infraorbital canal running through entire second infraorbital, extending to infraorbital 1 and opening into two pores. Preoperculomandibular branch giving rise to preoperculo-mandibular canal, which runs through entire preopercle with three openings, leading to pores 3, 4, and 5, respectively.
Dorsal fin triangular, located just posterior to second dorsolateral body plate. Dorsal-fin rays II,7 (1), II,8* (19), posterior margin of dorsal-fin spine with 10 to 14 serrations directed towards tip of spine; serrations absent proximally. Nuchal plate moderately developed; exposed, with minute odontodes; spinelet short; spine moderately developed, adpressed distal tip reaching to or slightly surpassing origin of last dorsal-fin branched ray; anterior margin with small odontodes. Pectoral fin triangular, its origin just posterior to gill opening. Pectoral-fin rays I,7* (15), I,8 (5); posterior margin of pectoral spine with 21 to 27 small serrations along its entire length; serrations directed towards pectoral-spine tip ( Fig. 3 View Fig ). Pelvic fin oblong, located just below first ventrolateral body plate, and at vertical through first branched dorsalfin ray. Pelvic-fin rays i,5. Adipose fin roughly triangular, separated from base of last dorsal-fin ray by generally seven dorsolateral body plates. Anal fin triangular, located just posterior to 12 th ventrolateral body plates, and at vertical through anterior margin of adipose-fin spine. Anal-fin rays ii,5* (3), ii,5,i (15), ii,6 (1), ii,6,i (1). Caudal-fin rays i,11,i (1), i,12,i* (19), generally four dorsal and ventral procurrent rays; bilobed; dorsal lobe slightly larger than ventral lobe.
Two or three laterosensory canals on trunk; first ossicle tubular, second ossicle laminar, and third lateralline canal, if present, encased in third dorsolateral body plate. Body plates with minute odontodes scattered over exposed area, a conspicuous line of odontodes confined on posterior margins; dorsolateral body plates 22 (2), 23* (16), 24 (2); ventrolateral body plates 20 (12), 21 (8); dorsolateral body plates along dorsal-fin base 6; dorsolateral body plates between adipose- and caudal-fin 6(1),7* (15),8 (4);preadipose platelets 2* (11), 3(9); small platelets covering base of caudal-fin rays; small platelets disposed dorsally and ventrally between junctions of lateral plates on posterior portion of caudal peduncle. Anterior margin of orbit with platelets above lateral ethmoid. Ventral surface of trunk generally without platelets; few specimens with scarce small platelets.
Color in alcohol. Overall color pattern in Figure 1 View Fig . Ground color of body grayish yellow, with top of head and snout dark brown. Ventral margin of orbit, above infraorbital 1, blackened. Maxillary barbel covered by black chromatophores. Dorsal and lateral portion of head, dorsal two thirds of dorsolateral body plates anterior to adipose fin, cleithrum and dorsal portion of ventrolateral body plates anterior to dorsal fin with conspicuously rounded black spots. Dorsal portion of ventrolateral body plates between dorsal-fin origin and adipose-fin origin with vertically elongated black spots. Dorsal portion of caudal peduncle close to caudal-fin origin blackened. Midline of flank posterior to dorsal-fin origin with longitudinal black stripe; posterior margin of dorso- and ventrolateral body plates conspicuously blackened, forming longitudinal zigzag. Longitudinal black stripe along midline of flank fragmented in juvenile specimens. Dorsolateral body plates close to longitudinal stripe on midline of flank unspotted. Dorsal fin covered by irregular black spots; membrane between dorsal spine and first branched dorsal-fin ray with concentration of black chromatophores, generally more concentrated in its dorsal half. Pectoral and pelvic fins unspotted, with concentration of brownish chromatophores on their rays. Anal fin with brown spots; spots aligned forming one to two oblique brown bars in some specimens; first and second anal fin rays markedly blackened in holotype. Adipose fin hyaline with distal margin blackened. Caudal fin with generally three transversal black bars.
Color in life. Very similar to preserved specimens but with ground color of body rosy orange; body covered by greenish yellow iridescent coloration ( Fig. 4 View Fig ).
Sexual dimorphism. Additionally to the lanceolate genital papilla in males, which is present in all Corydoradinae (see Nijssen & Isbrücker, 1980; Britto, 2003), the males of Corydoras knaacki have an oblong pelvic fin, while in females the pelvic fin is rounded.
Distribution. Corydoras knaacki is known from the río Madre de Dios basin, Peru ( Fig. 6).
Ecological notes. Corydoras knaacki was captured in a swampy area with brownish “tea-colored” stagnant and very clear water. Some congeners, like Corydoras aff. aeneus , Corydoras cf. elegans and Corydoras cf. stenocephalus , were observed in the type-locality of C. knaacki ( Fig. 7 View Fig ). Additionally, some Characiformes were also observed in syntopy, like Aphyocharax sp. , Hoplias aff. malabaricus ( Bloch, 1794) , Hyphessobrycon sp. and Pyrrhulina vittata, Regan, 1912 .
Juveniles of Corydoras knaacki , between 8 up to 15 mm SL, are abundant in the shallow swampy area, forming bigger shoals. Specimens of Corydoras aff. aeneus with similar size to the specimens of C. knaacki were observed mixing in the same shoal with the new species, which is more abundant. Adult specimens of C. knaacki , up to 38 mm SL, can be found in small groups only in the deeper parts of the creeks, generally shaded by trees and palms. The adults do not form mixed shoals with Corydoras aff. aeneus .
Etymology. Corydoras knaacki is named after Dr. Joachim Knaack (2 January 1933 – 5 December 2012), German physician and biologist. He was an amateur ichthyologist and aquarist who devoted more than 60 years of his life for the study of South American catfishes, especially Corydoras . A genitive noun.
Conservation status. Despite the new species seems to be relatively well distributed in the río Madre de Dios basin, part of its currently known region of occurrence, the río Inambari basin, was recently affected by gold mining and road building. Lujan et al. (2013) demonstrated that the aforementioned threats have already affected the stream community structure of some tributaries to the río Inambari as a whole. However, since there is no available data of the possible direct effects of the gold mining and/or road building in the populations of C. knaacki , and also due to the quite plausible possibility that the new species may occurs in other non-affected water bodies of the region, the most reasonable category for Corydoras knaacki for the time being, according to the International Union for Conservation of Nature (IUCN) categories and criteria (IUCN Standards and Petitions Subcommittee, 2014), is Least Concern (LC).
Remarks. Corydoradinae catfish are well known in the aquarium hobby and have been collected for the ornamental fish trade for many decades. Many newly encountered species are clearly recognized for being scientifically undescribed. To avoid the creation of nomina nuda by using trade names, Evers (1993) suggested to implement a code-system for all undetermined species, giving each species a “C-number” (“C” for “ Corydoradinae ”) in the German aquarist magazine DATZ (Die Aquarien- und Terrarienzeitschrift). Posteriorly, this system was carried on by the “ Corydoras World ” website (www.corydorasworld.com), and the codes were changed to CW-numbers (“CW” for “ Corydoras World ”). Corydoras knaacki , new species, is well known in the aquarium hobby, and has been previously known under the code number CW 032. Corydoradinae species display specific coloration and pigmentation pattern in juveniles, changing pattern during development after hatching onto reaching their adult coloration. Corydoras knaacki has been reproduced under aquarium conditions by the second author, and the color pattern of juvenile specimens (with less than 20.0 mm SL) are provided herein ( Fig. 7 View Fig ), documenting the unique development of this species during growth.





| ROM |
Royal Ontario Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |