
Haliclona (Halichoclona) arequipaensis, Bispo & Willenz & Hajdu, 2022
|
publication ID |
https://doi.org/10.11646/zootaxa.5087.2.1 |
|
publication LSID |
lsid:zoobank.org:pub:4B472D23-386F-497F-A6DA-8867C081D6D8 |
|
DOI |
https://doi.org/10.5281/zenodo.5824041 |
|
persistent identifier |
https://treatment.plazi.org/id/0A10034B-2948-0D53-7DC7-FF5F6D8CFD42 |
|
treatment provided by |
Plazi (2022-01-06 07:10:12, last updated 2024-11-28 21:50:17) |
|
scientific name |
Haliclona (Halichoclona) arequipaensis |
| status |
sp. nov. |
Haliclona (Halichoclona) arequipaensis View in CoL sp. nov.
( Figure 6 View FIGURE 6 , Table 3 View TABLE 3 )
Holotype. MNRJ 12140 View Materials (Vouchers: RBINS-IG 32240 -POR 12140, MHNG 85947 View Materials )— Playa Catarindo, Mollendo , Arequipa Region (17°01’08.93” S, 72°02’03.25” W), depth 4–5 m, coll. Y. Hooker, U. Zanabria & Ph. Willenz (26/ XI/2008) GoogleMaps . Paratype. MNRJ 12147 View Materials (Vouchers: RBINS-IG 32240 -POR 12147, MHNG 85589 View Materials ) — Punta Hornillos, Matarani , Arequipa Region (16°52’49.80” S, 72°17’18.24” W), depth 15 m, coll. Y. Hooker, U. Zanabria & Ph. Willenz (28/XI/2008) GoogleMaps .
Diagnosis. The only Haliclona in the Eastern Pacific with the combination of thickly encrusting habit, reaching up to 30 cm in diameter, eventually with erect, lamellate or crest-like projections, dull pink colour alive, dense isotropic reticulation with few paucispicular tracts in the choanosome, and oxeas reaching up to 108–198 µm in length.
Description ( Fig. 6A, B View FIGURE 6 ). Thickly crustose, up to 5 mm thick, over 30 cm in largest diameter (MNRJ 12140), forming erect, lamellate or crest-like projections (MNRJ 12147, expanding distally, terminally serrated) up to 60 mm high x 20 mm wide x 5 mm thick. Surface regularly smooth to the naked eye, minutely reticulated upon closer inspection. Oscula, 1–2 mm in diameter, spread all over the sponge surface, apical on abundant, 1 mm high volcaniform bumps. Consistency resilient, flexible. Colour in life dull pink.
Skeleton ( Fig. 6C, D View FIGURE 6 ). Ectosome a dense, slightly confused, tangential isotropic reticulation. Choanosome a dense, uni- to multispicular isotropic reticulation, with 2–10 oxeas by knot, pierced here and there by subectosomal and choanosomal spaces, up to 900 µm in diameter. Tracts nearly totally absent, the few seen were loose, paucispicular, parallel to the surface, albeit deep in the choanosome. Spongin scarce, only observed at the nodes of the reticulation.
Spicules ( Fig. 6E–G View FIGURE 6 ). Oxeas, fusiform, straight, or more frequently subtly bent at centre, sharp acerate points, holotype: 123– 161.3 –198 x 2.4– 7.1 –12.6 µm (n = 40 x 40); paratype: 108– 147.2 –178 x 2.2– 7.3 –11.9 µm (n = 40 x 40).
Ecology. Specimens collected from erect rocky substrate; co-occurring with limpets, shrimps, anemones, additional sponges (including Niphates ruthecitae sp. nov., see below), and large seastars. Several crabs (hermit crabs and others) were observed on the surface of H. (Halich.) arequipaensis sp. nov. The paratype carried a thin brown (turf?) mat in parts of its surface. Recorded depth was 4 to 15 meters. Water temperature during collections was 15–16°C.
Distribution ( Fig. 3C View FIGURE 3 ). Only known from areas close to Matarani and Mollendo (Arequipa Region), in Peru.
Etymology. The epithet “ arequipaensis ” refers to the Arequipa Region where both specimens were collected.
Remarks. There are five other Haliclona spp. along the Eastern Pacific that approach the most H. (Halich.) arequipaensis sp. nov. regarding their shape and/or skeletal architecture: H. agglutinata Desqueyroux-Faúndez, 1990 , from Easter Island; H. (Halich.) algicola , from Chile; H. (Halicl.) dianae Sim-Smith, Hickman Jr. & Kelly, 2021 from Galápagos; H. (Halich.) gellindra ( de Laubenfels, 1932) , from California; H. (Halich.) paracas sp. nov., from Peru; H. rapanui ( Desqueyroux-Faúndez, 1990) , from Easter Island; and Haliclona (Halich.) thielei van Soest & Hooper, 2020, from Chile ( Table 3 View TABLE 3 ).
The encrusting shape, punctate surface, confused subisotropic skeleton, oxeas 105–122 µm long, and pale lavender colour in life ( de Laubenfels 1932; de Weerdt 2002) make H. (Halich.) gellindra the most similar to the new species. Though its few and irregular oscula bearing raised collars about 1 mm high make conspecificty with H. (Halich.) arequipaensis sp. nov. unlikely. Additionally, H. (Halich.) gellindra is only known from the California coast ( de Laubenfels 1932) and the Gulf of California ( Dickinson 1945), separated from the Peruvian coast by the warmer waters of the Tropical Eastern Pacific.
H. (Halich.) arequipaensis sp. nov. is readily distinguished from H. agglutinata due to the latter’s massive shape, reaching 10–28 cm in thickness ( Desqueyroux-Faúndez 1990), in contrast to a thickly encrusting shape, 5 mm thick, in H. (Halich.) arequipaensis sp. nov. Divergence in colour and dimensions of the oxeas only marginally overlapping ensure H. (Halich.) arequipaensis sp. nov. and H. (Halich.) algicola as distinct species. The latter has a light grey colour alive and oxeas of 193–230 x 4–12 µm ( Thiele 1905; Hajdu et al. 2013), contrasting with the dull pink colour in life and oxeas ranging between 108–198 x 2–13 µm in H. (Halich.) arequipaensis sp. nov. Haliclona (Halicl.) dianae is reddish pink in colour with a regular anisotropic reticulation of uni- to paucispicular primary lines ( Sim-Smith et al. 2021), such an arrangement is distinct from the dense, slightly confused and isotropic reticulation present in the new species. In its turn, H. rapanui has a thickly encrusting habit, 19 mm thick, hispid surface, and lacks an ectosome ( Desqueyroux-Faúndez 1990), whilst H. (Halich.) arequipaensis sp. nov. has 5 mm in thickness, a smooth surface, and a tangential ectosome. Comparison with H. (Halich.) paracas sp. nov. is provided below in the remarks section of the latter species.
Haliclona (Halich.) thielei is an intertidal species from the Caleta Tumbes area in Chile ( Hajdu et al. 2013). The original description of this species was based on several fragments in a same jar that Thiele (1905) doubtfully considered as part of an interspecific variability in two forms: one has a dense and irregular skeleton (from now on called “ Spicule reinforced form”) and the other with a regular skeleton of oxeas and abundant spongin.(from now on called “ Spongin reinforced form). Nevertheless, these differences regarding the skeletal architecture and oxeas’ dimensions in both forms indicate the presence of, at least, two species in the type material of H. (Halich.) thielei. Thus, a taxonomic revision of this species is necessary. Notwithstanding, the “ Spicule reinforced form” of H. (Halich.) thielei is similar to H. (Halich.) arequipaensis sp. nov., though both are distinct based on the following features: H. (Halich.) thielei “ Spicule reinforced form” has blue-green or violet-grey colour alive, more organized skeleton close to the surface, with discernible paucispicular tracts (similar to Rhizoniera subgenus architecture), and shorter oxeas of 140–150 µm. In turn, H. (Halich.) arequipaensis sp. nov. has dull pink colour alive, an isotropic skeleton, with loose paucispicular tracts parallel to the surface in the deeper choanosome, and oxeas occurring in a larger size range, 108–198 µm.
de Laubenfels, M. W. (1932) The marine and fresh-water sponges of California. Proceedings of the United States National Museum, 81 (2927), 1 - 140. https: // doi. org / 10.5479 / si. 00963801.81 - 2927.1
de Weerdt, W. H. (2002) Family Chalinidae Gray, 1867. In: Hooper, J. N. A. & van Soest, R. W. M. (Eds.), Systema Porifera: A Guide to the Classification of Sponges. Springer US, Boston, Massachusetts, pp. 852 - 873. https: // doi. org / 10.1007 / 978 - 1 - 4615 - 0747 - 5 _ 91
Desqueyroux-Faundez, R. (1990) Sponges (Demospongiae) from Easter Island (Isla de Pascua) (South Pacific Ocean). Revue suisse de Zoologie, 97 (2), 373 - 410.
Dickinson, M. G. (1945) Sponges of the Gulf of California. In: Reports on the collections obtained by Alan Hancock Pacific Expeditions of Velero III off the coast of Mexico, Central America, South America, and Galapagos Islands in 1932, in 1933, in 1934, in 1937, in 1939, and 1940. The University of Southern California Press, Los Angeles, California, pp. 1 - 55.
Hajdu, E., Desqueyroux-Faundez, R., Carvalho, M. de S., Lobo-Hajdu, G. & Willenz, Ph. (2013) Twelve new Demospongiae (Porifera) from Chilean fjords, with remarks upon sponge-derived biogeographic compartments in the SE Pacific. Zootaxa, 3744 (1), 1 - 64. https: // doi. org / 10.11646 / zootaxa. 3744.1.1
Sim-Smith, C., Hickman Jr, C. & Kelly, M. (2021) New shallow-water sponges (Porifera) from the Galapagos Islands. Zootaxa, 5012 (1), 1 - 71. https: // doi. org / 10.11646 / zootaxa. 5012.1.1
Thiele, J. (1905) Die Kiesel- und Hornschwamme der Sammlung Plate. Zoologische Jahrbucher, Supplement 6 (Fauna Chilensis III), 407 - 496.

FIGURE 3. Map showing the geographic distribution of Peruvian haplosclerids. A, distribution of Chalinula chelysa sp. nov.; B, distribution of Chalinula ramiculosa sp. nov. and Haliclona (G.) concreta sp nov.; C, distribution of H. (Halich.) arequipaensis sp. nov., H. (Halich.) multiosculata sp. nov. and H. (Halich.) marcoriosi sp. nov.; D, distribution of H. (Halich.) paracas sp. nov., H. (Halich.) pellucida sp. nov. and (Reniera) parvuloxea sp. nov.; E, distribution of H. (Rhizoniera) spp. nov.; F, distribution of Niphates ruthecitae sp. nov. and Pachychalina lupusapia sp. nov.
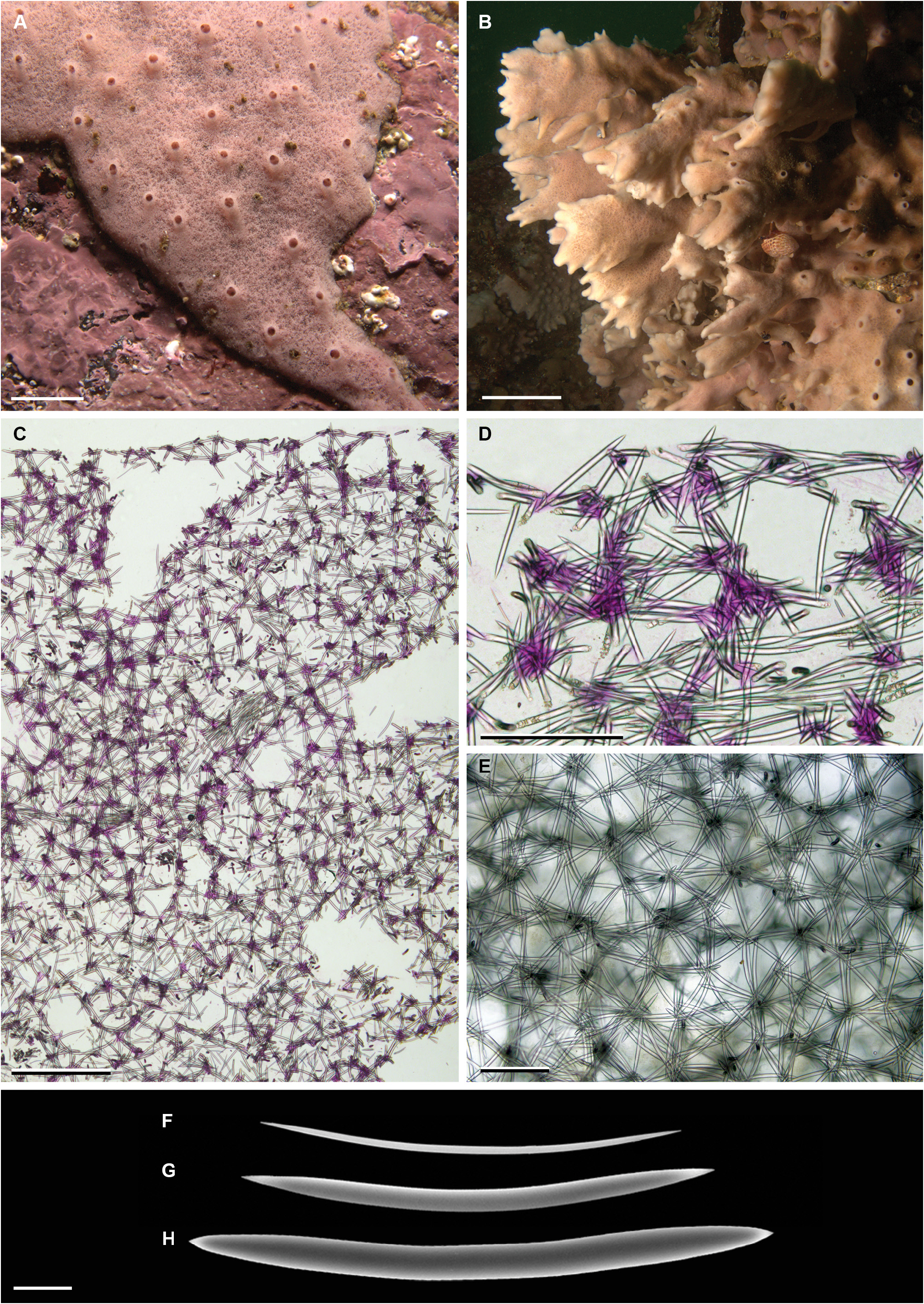
FIGURE 6. Haliclona (Halichoclona) arequipaensis sp. nov. A, holotype alive (MNRJ 12140); B, paratype alive (MNRJ 12147); C–D, skeletal architecture in transverse ground section (MNRJ 12140, holotype); E, ectosomal skeleton architecture in tangential ground section (MNRJ 12147, paratype); F–H, oxeas (MNRJ 12147, paratype). Scale bars: A, 1 cm; B, 2 cm; C, 500 µm; D–E, 200 µm; F–H, 20 µm.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
|
SubGenus |
Haliclona |