
Dendronotus regius, Pola & Stout, 2008
|
publication ID |
https://doi.org/ 10.11646/zootaxa.1960.1.2 |
|
DOI |
https://doi.org/10.5281/zenodo.5242530 |
|
persistent identifier |
https://treatment.plazi.org/id/087D7947-FF83-FFE4-1A86-264D8850FE42 |
|
treatment provided by |
Felipe |
|
scientific name |
Dendronotus regius |
| status |
sp. nov. |
Dendronotus regius View in CoL sp. nov.
( Figures 1A–C View FIGURE 1 , 2 View FIGURE 2 , 3 View FIGURE 3 , 4 View FIGURE 4 , 5 View FIGURE 5 )
Material Examined: Holotype: CASIZ 087013 , Indonesia, Sulawesi, Celebes Sea, Manado, Bunaken Island , under rock. 12 mm alive (6 mm preserved), 46 m depth, 15 May 1990, collected by P. Fiene. Paratypes: CASIZ 175613 , one adult specimen completely dissected, 15 mm alive (same locality, date and collector as holotype) . CASIZ 078543 , Indonesia, Sulawesi, Celebes Sea, Manado, Bunaken Island , found out of rock, 1 adult specimen 9 mm alive (6 mm preserved), 40 m depth, 23 May 1991, collected by P. Fiene. CASIZ 097622 , Indonesia, Lembeh Strait , three adult specimens 5, 6 and 10 mm preserved, all dissected, collected by D.J. Ecenbarger.
Etymology: The specific epithet refers to the color pattern of the animal, which resembles the overcoat of a king. From Latin: "regius" = royal. Also, this species is dedicated to Joaquim Reis (From Portuguese: "rei" = king).
Distribution: Thus far, only known from Indonesia (present study; Warren 1998, 2000; Kodiat 2005; Tomeno 2006; Dotulong 2006; Huang 2007), Philippines ( Izumi 2007) and Malaysia ( Grall 2008).
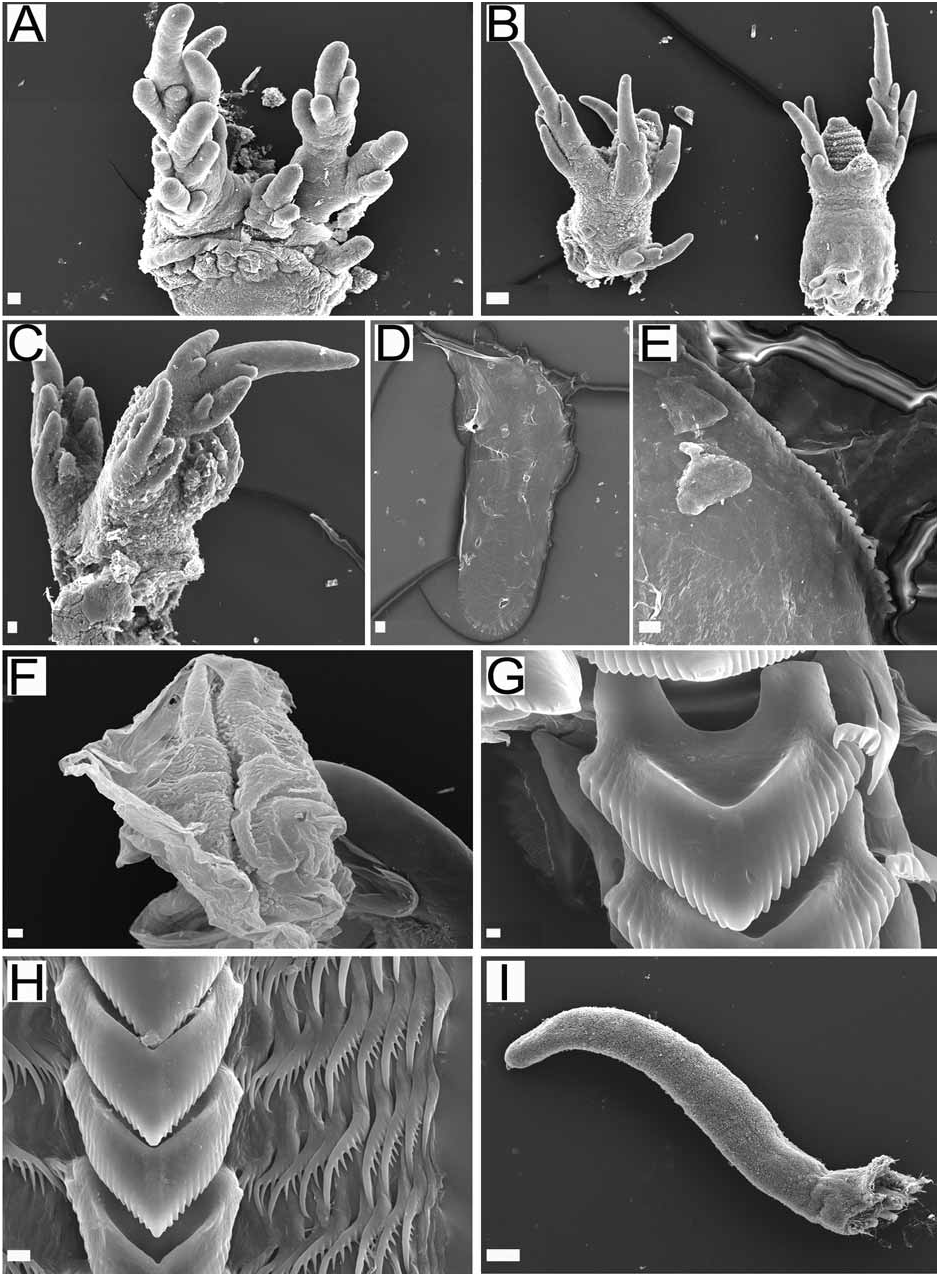
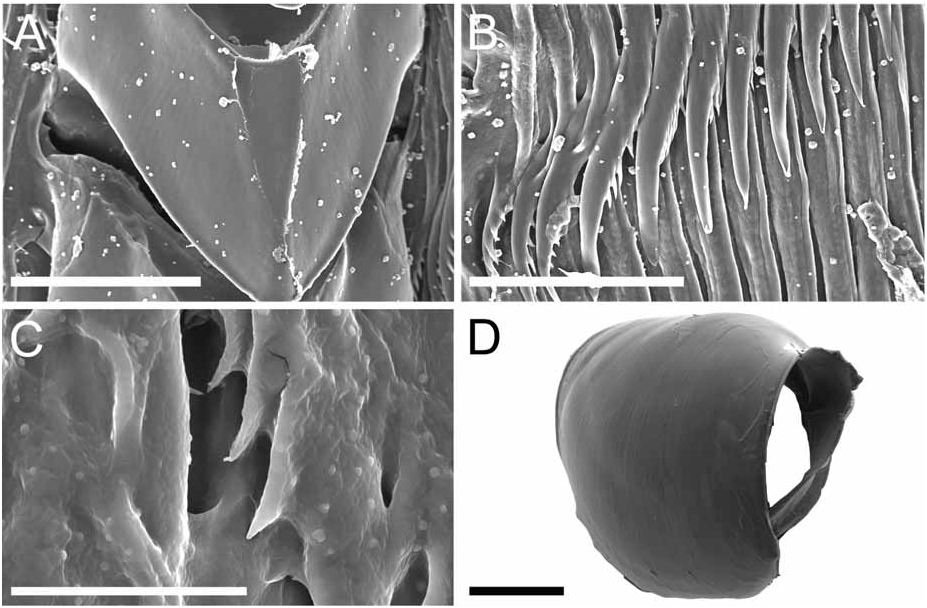
External morphology ( Fig. 1A–C View FIGURE 1 ): The general body shape is delicate and limaciform, laterally compressed, with a rounded dorsum and a tapering, pointed posterior end of the foot. The foot is narrow, rounded in front, and not sharply marked off from the body. The notum is not distinctly marked other than by the line of dorsolateral processes. The lateral sides of the body surface have a few low tubercles. The cardiac elevation is prominent. The body wall musculature is thin. The anterior end of the body is rounded with three different branched groups of veil papillae on either side of the mouth ( Fig. 2A View FIGURE 2 ). The series nearest the mouth are the shortest, with three small branches. Laterally there is a second tentacle made up of eight branches. The dorsalmost branch is the most branched, with a main long stalk and many shorter branches arising from it. The rhinophores consist of very tall stalks inclined forward. The sheath margin extends into four elongate and branched processes, most of them with a main longer papilla and three to four smaller ones ( Fig. 2B View FIGURE 2 ). The most posterior papilla is larger than the others and also is divided into at least four branches. Midway between the base of the rhinophoral sheath and the sheath margin there is another laterally extending branch with threefour short papillae. The rhinophores are perfoliate with about 10 to 11 lamellae. There are three to four pairs of strongly branched dorsolateral processes along the margins of the dorsum. The first pair of dorsolateral processes is the largest and most branched ( Fig. 2C View FIGURE 2 ) and the remaining pairs decrease in size and number of branches toward the posterior end of the body. Each dorsal process has a strong stalk, which ends in an elongate papilla several times branched on its base. Half way between the base of the dorsolateral process and the branches of the main papilla there are four smaller papillae. These papillae are shorter and branch three to four times with these branches extending from around each side of the main stalk. The anal opening is along the dorsolateral line about midway between the first and second dorsolateral processes on the right side. The small renal pore is just dorsal to the anal opening. The reproductive openings are on the right side below the dorsolateral line, about midway between the rhinophore and the first dorsolateral process.
The preserved specimen is white with brown patches all over the surface, including the stalk and branches of the rhinophoral sheaths and dorsolateral processes. The lamellae of the rhinophores are normally brown. A color picture of one animal alive (CASIZ 078543) shows a yellow background color pattern, with the patches described above as dark blue or brown ( Fig. 1A View FIGURE 1 ) and with orange rhinophoral lamellae. Slight variation in this color theme has been observed and photographed ( Rudman 2000). These photographs show animals with a creamy white body with pale orange on the notum and irregular dark brown patches down its length ( Fig. 1B–C View FIGURE 1 ).
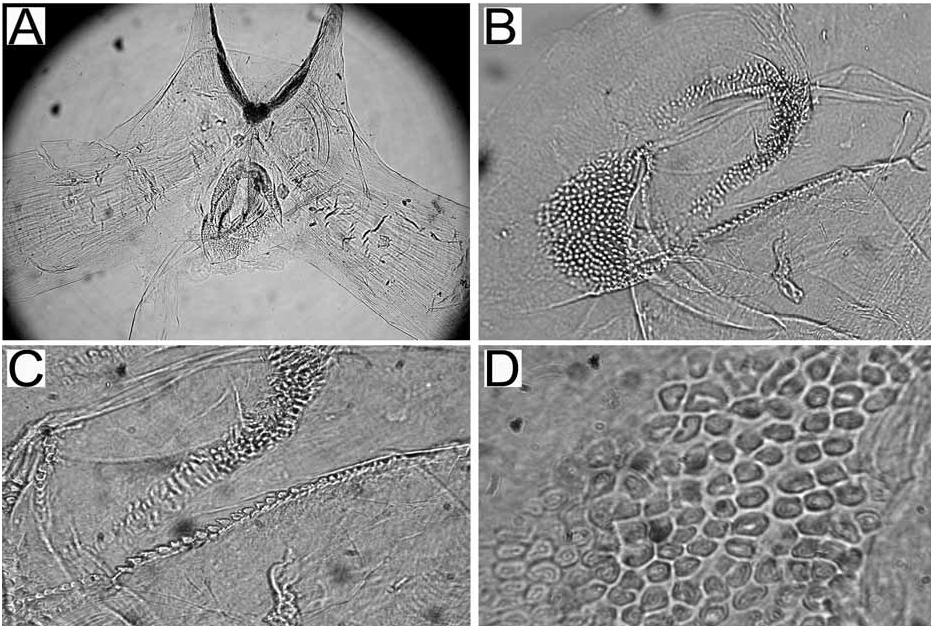
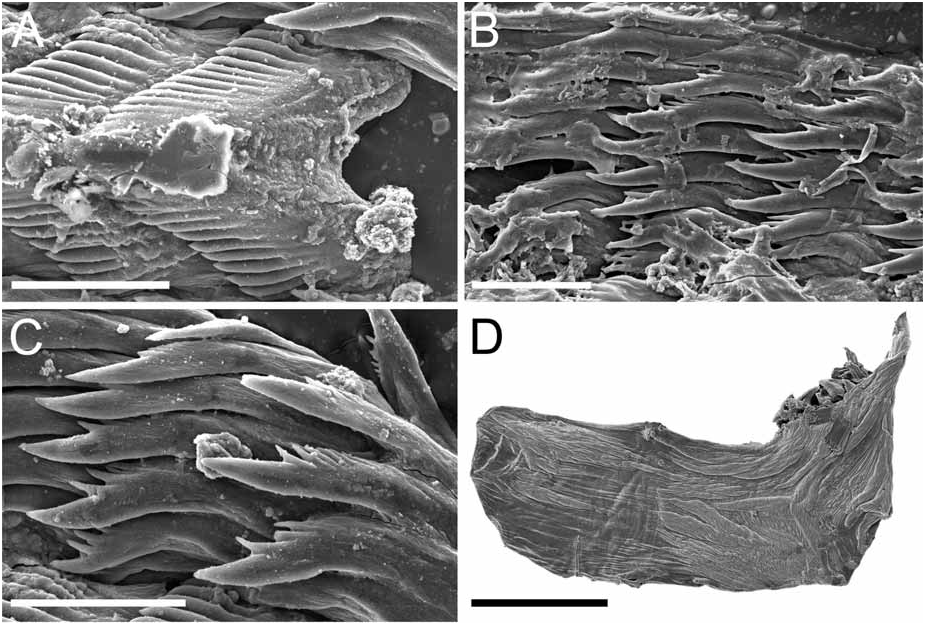
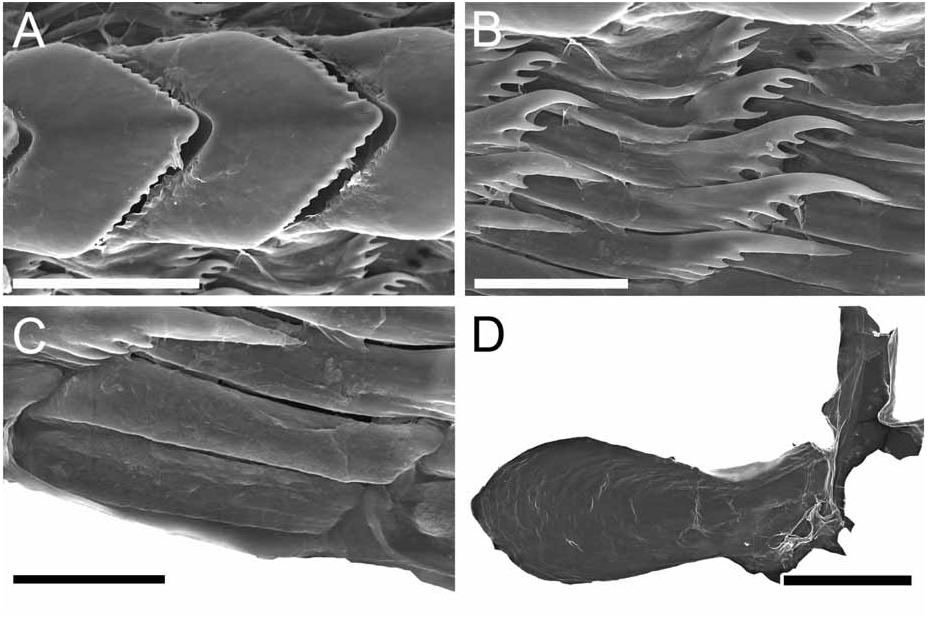
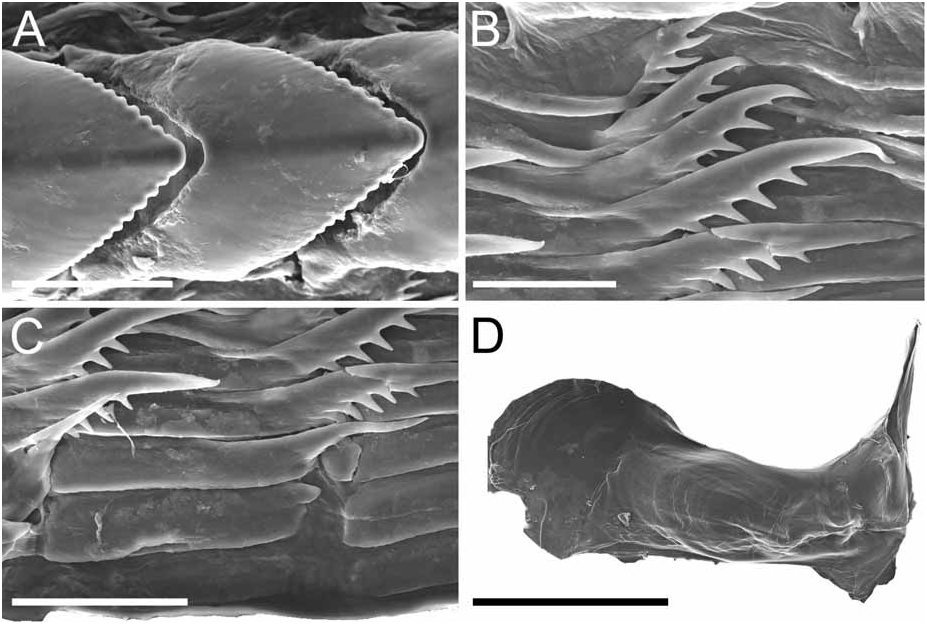
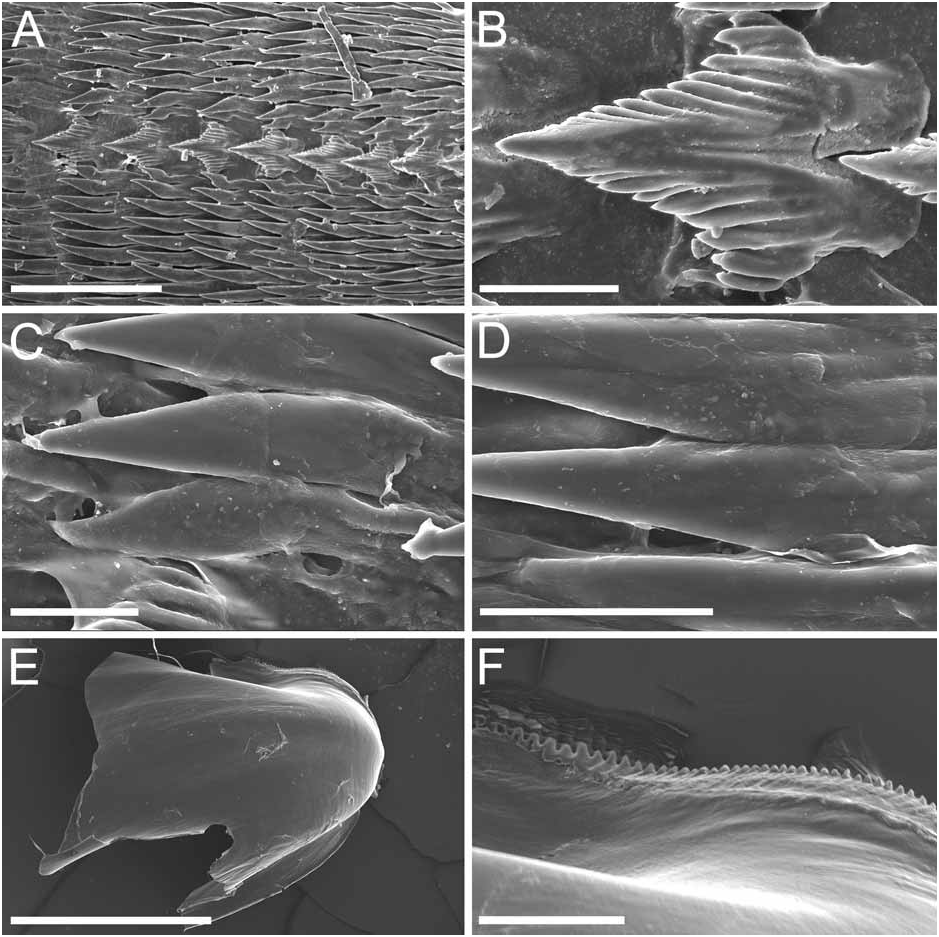
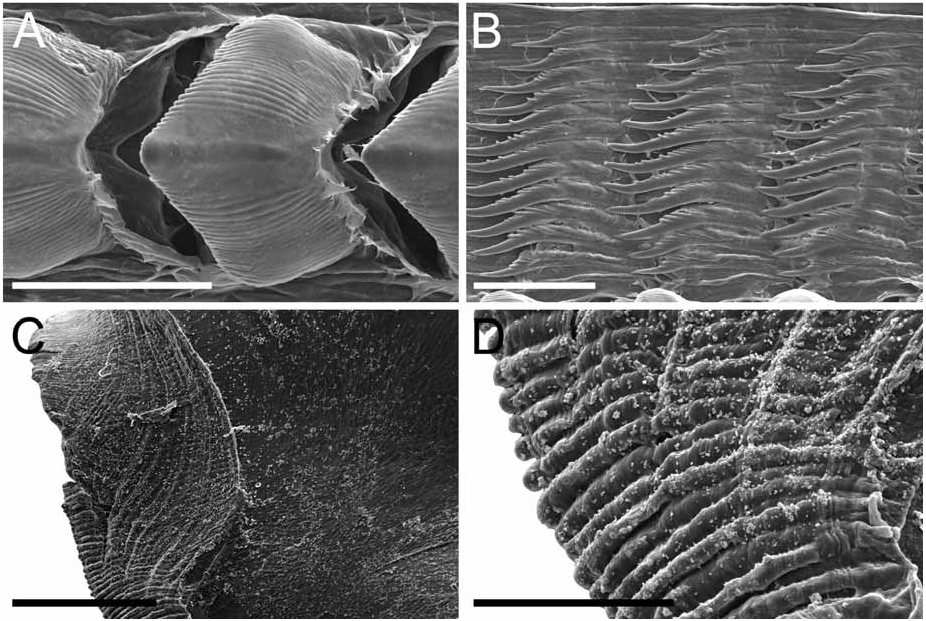
Alimentary Canal: The anterior digestive tract begins with an oval-shaped mouth opening surrounded by glandular, external lips. Internally, there are large tubular labial glands surrounding the lateral sides of the mouth and a high number of smaller, rounded frontal glands. The inner lips are strongly muscular and their inner concave face is covered with a strong cuticle that thickens progressively backward and forms a prehensile collar that surrounds the inner opening of the mouth just in front of the masticatory processes of the jaws ( Figs 2F View FIGURE 2 ; 3A–B View FIGURE 3 ). From this point the cuticle tends to fray out into an irregular fringe of blunt rodlets ( Fig. 3B View FIGURE 3 ). The jaws are elongate and concave in form ( Figs 2D View FIGURE 2 ; 3A View FIGURE 3 ) with a strongly convex posterior border. The masticatory process ( Figs 2E View FIGURE 2 ; 3A–C View FIGURE 3 ) is united to the ventral border of the mandible by a thick, arched expansion. The margin of the process bears a series of 20–30 denticles. These begin a short distance below the hinge as slight elevations terminating in short, low, parallel ridges on the outer face of the process. These gradually increase in size to form distinct ridge-like denticles ( Figs. 2E View FIGURE 2 ; 3A–C View FIGURE 3 ). The radular formulae are: 15 mm specimen, 36 x 9.1.9 (CASIZ 175613), 6 mm specimens, 35 x 7.1.7 (CASIZ 078543) and 31 x 7.1.7, 32 x 6.1.6 (CASIZ 097622). The median teeth are large and strong with a quadrilateral base and a strong triangular cusp directed upward and backward at an angle of about 45° to the base ( Fig. 2G View FIGURE 2 ). The lateral margins are highly denticulate, bearing 15 to 20 long and pointed denticles that decrease in length as they approach the cusp ( Fig. 2G–H View FIGURE 2 ). There are from six to nine lateral teeth, each consisting of a thin, elongate, flattened plate. Their posterior thickened margins are elevated and prolonged into strong points and curve towards the mid-line of the radula. The outer border of the cusps bears a series of six to seven sharp denticles. The outermost lateral tooth is narrow with a straight and smooth apex ( Fig. 2H View FIGURE 2 ). The buccal bulb is large, elongate, muscular and mostly rectangular in shape. The salivary glands consist of one pair. Their ducts pass through the nerve collar and enter the dorsal side of the buccal bulb close behind the exit of the oesophagus. The oesophagus emerges from the pharyngeal bulb and is short and wide. It opens into a rounded and large stomach that is thickened with folds. The stomach loops downward on the right, doubles sharply back upward, passes transversely over the dorsal face of the viscera and narrows into the intestine, which loops back down the right side to the anus. Two anterior and one posterior thin-walled lobes of the digestive gland open into the anterior region of the stomach. The anterior lobes send large branches up into the bases of the first pair of dorsolateral processes and to the rhinophoral sheaths. The posterior lobe lies beneath the lobules of the ovotestis throughout its entire length and branches into all of the posterior dorsolateral processes on each side.
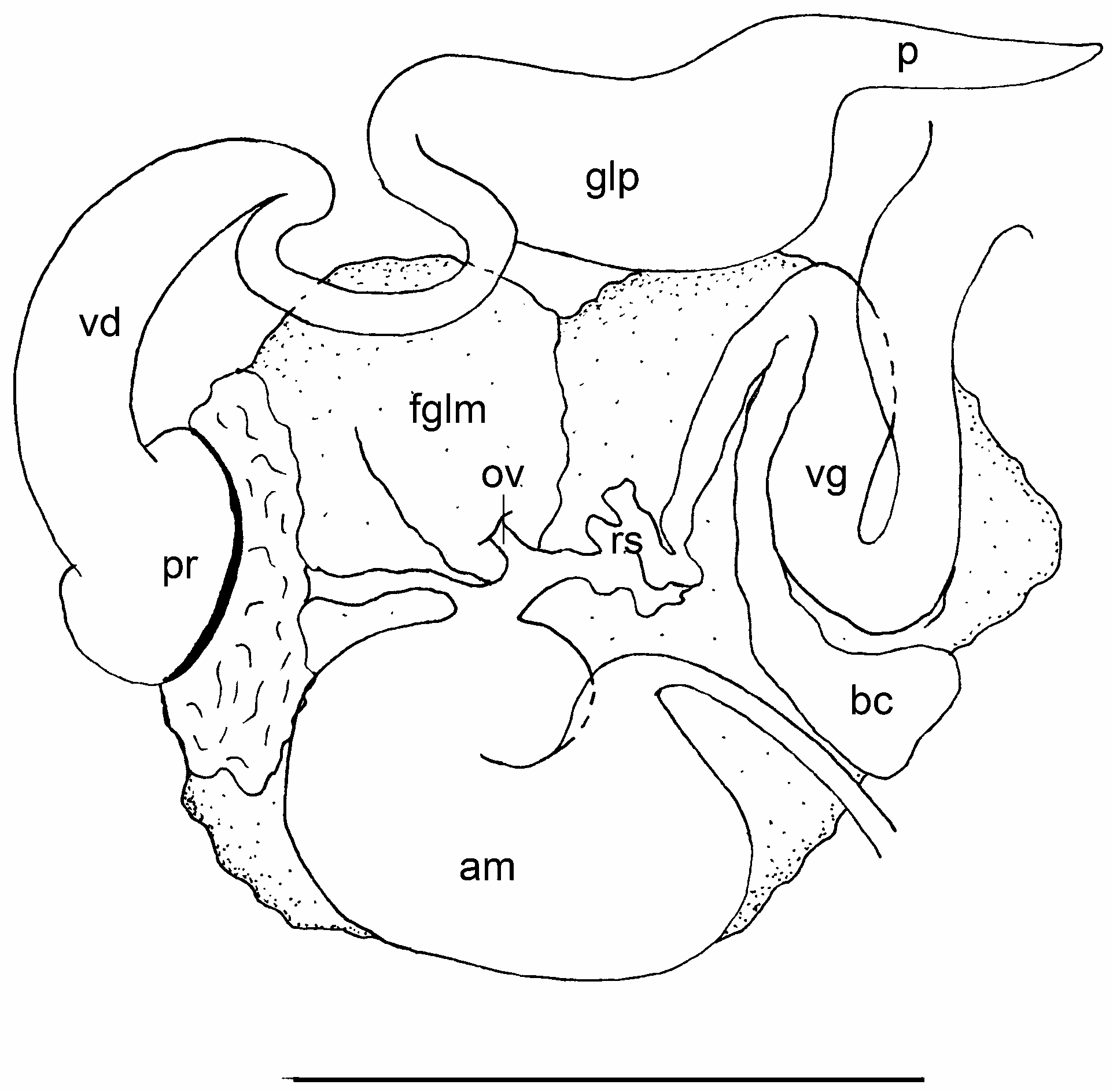
Reproductive system ( Fig. 4 View FIGURE 4 ): The reproductive system is triaulic. The ovotestis is made of up to twelve closely packed, rounded or somewhat pyriform lobules lying above the digestive gland in the posterior part of the body. The hermaphroditic duct passes forward from the ovotestis and expands into a sausage-shaped ampulla, which forms a closed loop upon the inner face of the anterior genital complex. Three ducts arise from the ampulla. The most anterior duct narrows and gives rise to the vas deferens. The proximal segment of the vas deferens widens at once into a large prostate with the proximal limit being marked by a closely set ring of alveolar glands. Beyond the group of alveoli, the vas deferens is coiled and gradually narrows towards the base of a large preputial sac. The penis is unarmed and tapering ( Fig. 2I View FIGURE 2 ). The next duct (oviduct) extends almost immediately into the thin-walled fertilization chamber of the female glandular mass. The third duct terminates in a long and wide vagina. Upon its second distal loop from the ampulla it bears a well-developed bursa copulatrix beyond which the vagina tapers into the vaginal duct. Just before the entrance into the gland complex, the proximal end of the vagina forms a very coiled and irregular section, which we think could work as seminal receptacle. The nidamental gland opens separately from the vagina.
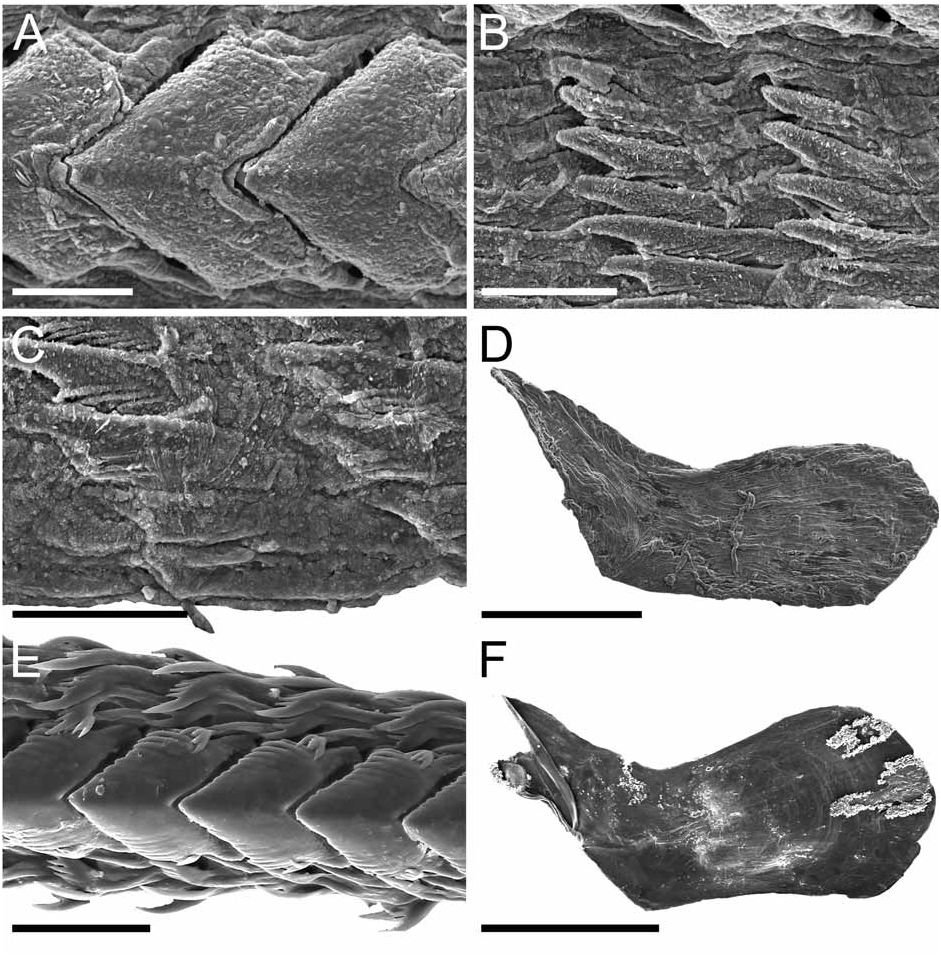
Remarks: Dendronotus regius is clearly distinctive in its external appearance from other species of the genus. Externally, the only already described species with even a remotely similar color pattern is D. lacteus (Thompson) , which is found in European waters (Table 1). Dendronotus lacteus has an opaque white body with brown spots, but these spots are much smaller and spread further apart. The rhinophores of D. lacteus are white instead of orange or brown (Table 1). The tips of dorsal processes and papillae in this new species are dark brown, while in D. lacteus are white ( Thollesson 1998). This specimen is also clearly different from D. lacteus in radula morphology. Dendronotus lacteus has no denticulation on the lateral teeth according to Thollesson (1998) or minimal denticulation ( Fig. 13C View FIGURE 13 ) and the median tooth of D. lacteus ( Fig. 13B View FIGURE 13 ) is not as elongate as the one found in D. regius . Regarding the reproductive system, D. lacteus has a much longer and convoluted distal vas deferens ( Thollesson 1998) than D. regius . Because this species has a large cardiac prominence and body tubercles, it must also be compared to D. frondosus (Ascanius) , D. albus MacFarland and D. subramosus MacFarland. Branching patterns of the dorsolateral processes are very different. It is unusual in Dendronotus to find such a robust, thick primary stalk compared to the small, delicate secondary and tertiary branches that extend from it. The most unique feature in the reproductive system of this new species is the vas deferens. It is thicker and shorter than any other species of Dendronotus . It is most similar to D. subramosus , but there are many other differences to distinguish these two, for example in the location, size, and shape of the bursa copulatrix. In D. subramosus the bursa copulatrix is very small, rounded, and located very close to the opening of the vagina ( Robilliard 1970). Details of the radula of D. albus MacFarland , D. frondosus (Ascanius) and D. subramosus MacFarland are shown in Figures 8 View FIGURE 8 , 11 View FIGURE 11 and 16 View FIGURE 16 , respectively. The alveolar glands of the prostate are also much larger and fewer in number than in D. regius . In the radula, the median tooth of D. regius (and D. noahi ) is also unique. In these new species, in the inner margin of the tooth, the width of the chevron seems to decrease towards the lateral sides of each tooth ( Fig. 5 View FIGURE 5 ). However, other Dendronotus species have median teeth in which the width of the chevron shape does not change from the left end of the tooth to the right end. Figures 7–16 View FIGURE 7 View FIGURE 8 View FIGURE 9 View FIGURE 10 View FIGURE 11 View FIGURE 12 View FIGURE 13 View FIGURE 14 View FIGURE 15 View FIGURE 16 show this character and other radula and jaw details for most already described species. We discuss further comparisons between both new species under the remarks following the description of D. noahi .





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.