
Rineloricaria cacerensis
|
publication ID |
https://doi.org/ 10.1590/S1679-62252012000200006 |
|
persistent identifier |
https://treatment.plazi.org/id/0748878F-FF92-0355-FCF2-FD22FF58FBC2 |
|
treatment provided by |
Felipe |
|
scientific name |
Rineloricaria cacerensis |
| status |
|
Rineloricaria cacerensis View in CoL ( Miranda Ribeiro, 1912) Fig. 7 View Fig
Loricaria cacerensis View in CoL Miranda Ribeiro, 1912: 11 [original description; syntypes: MNRJ 645 and MNRJ 30030; type locality: Paraguay River, Cáceres, Estado de Mato Grosso, Brazil]. - Miranda Ribeiro, 1953: 400 [designation of lectotype: MNRJ 645].
Rineloricaria cacerensis View in CoL . - Fowler, 1954: 113 [listed; Paraguay River]. - Isbrücker & Nijssen, 1976: 111 [listed]. -Isbrücker, 1978: 87 [listed]. - Britski et al., 1999:130 [short transcription of the original description; Brazilian Pantanal]. - Ferraris, 2003: 341 [listed; Paraguay River basin in Brazil]. - Britski et al., 2007: 160 [short transcription of the original description; Brazilian Pantanal]. - Ghazzi & Oyakawa, 2007: 90 [listed; upper Paraguay River basin in Brazil].
Hemiloricaria cacerensis View in CoL . - Isbrücker et al., 2001: 23 [listed]. -Isbrücker, 2001: 27 [listed]. - Ferraris, 2007: 245 [listed; Estado de Mato Grosso, Brazil].
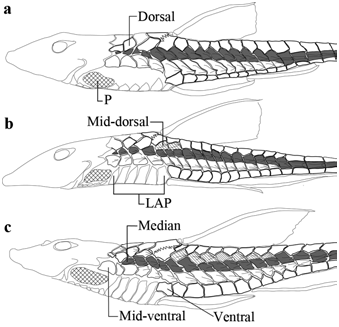
Diagnosis. Rineloricaria cacerensis is distinguished from most of its congeners by having five lateral plate series in longitudinal rows below the dorsal fin, the mid-dorsal series present ( Fig. 4c View Fig ) [vs. four lateral plate series in longitudinal rows below the dorsal fin, the mid-dorsal series absent ( Fig. 4a View Fig ) in R. aurata , R. beni , R. cadeae , R. castroi , R. catamarcensis , R. felipponei , R. lanceolata , R. langei , R. longicauda , R. magdalenae , R. misionera , R. nigricauda , R. pareiacantha , R. parva , R. quadrensis , R. sanga , R. setepovos , R. sneiderni , R. stellata , R. strigilata , R. thrissoceps , R. uracantha , and R. wolfei ]; and by presenting the lateral region of the body posterior to the pectoral-fin insertion with a slender area of naked skin, narrower than the caudal peduncle width ( Fig. 4c View Fig ) [vs. with a broad area of naked skin, similar in thickness to the caudal peduncle width ( Fig. 4a View Fig ) in R. aequalicuspis , R. anhanguapitan , R. anitae , R. baliola , R. capitonia , R. isaaci , R. jaraguensis , R. kronei , R. latirostris , R. maacki , R. malabarbai , R. maquinensis , R. microlepidogaster , R. pentamaculata , R. reisi , R. rupestris , R. stewarti , and R. tropeira ]. It is distinguished from remaining members of the genus, except R. daraha , R. fallax , R. formosa , R. hasemani , R. jubata , R. melini , R. morrowi , R. osvaldoi , R. teffeana , and R. zaina , by having the lateral plate of the mid-dorsal series consisting in five to seven keeled plates extended posteriorly around end of the dorsal-fin base ( Fig. 4c View Fig ) [vs. lateral plate of the mid-dorsal series consisting in two keeled plates situated around insertion of first ray of the dorsal fin ( Fig. 4b View Fig ) in R. altipinnis , R. caracasensis , R. eigenmanni , R. heteroptera , R. konopickyi , R. phoxocephala , R. platyura , and R. steindachneri ]. It is distinguished from R. daraha , R. osvaldoi , and R. zaina by having three longitudinal series of abdominal plates on the median complex (vs. several unorganized abdominal plates on the median complex in R. daraha ; five longitudinal series of abdominal plates on the median complex in R. osvaldoi and R. zaina ). It is distinguished from R. fallax , R. formosa , R. hasemani , R. jubata , R. melini , R. morrowi , and R. teffeana by the caudal fin coloring, consisting in a plain dark brown caudal fin on branched rays and membranes, and a barred superior and inferior unbranched rays (vs. caudal fin excepting outer unbranched rays with a small black stain at its base followed by a broad light-colored area with dark bars on rays and a terminal black margin in R. hasemani , R. jubata , R. melini , R. morrowi , and R. teffeana ; caudal fin dark brown with vertical darker bars on rays in R. fallax and R. formosa ). It is further distinguished from R. fallax , R. formosa , R. hasemani , R. jubata , R. morrowi , and R. teffeana by having the dorsum of body with large irregular spots (vs. dorsum with a single predorsal spot in R. fallax ; dorsum with small spots in R. formosa and R. morrowi ; and dorsum without spots in R. hasemani , R. jubata , and R. teffeana ); and from R. melini by having the dorsum of head without dark cross bars (vs. head with four dark cross bars).
Description. Morphometric data presented in Table 3. Snout tip raised in lateral view. Dorsal profile from snout tip to posterior nasal plates straight, convex from frontals to nuchal plate, declined at dorsal-fin base, straight from end of dorsal-fin base to penultimate dorsal plate, caudal fin damaged on three syntypes. Ventral profile from snout tip to anal-fin base convex, straight from that point to penultimate ventral plate, caudal fin damaged on three syntypes.
Snout tip pointed in dorsal view, lateral margins of head curved anteriorly. Odontodes of head and trunk small, thick, aligned. Dorsum of head smooth, without ridges. Infraorbital series with six bones, infraorbital 1 with a sensorial pore exposed ventrally, infraorbitals 2, 3, and 4 convexly raised from anterior portion of snout to eye. Rostral plate with an area of naked skin without odontodes at tip of snout, narrow, horizontally elongated, short, not reaching sensorial pore ventrally exposed of infraorbital 1. Posterior margin of parieto-supraoccipital slightly concave to straight, lateral margins truncate. Predorsal plates and first three lateral plates of dorsal series with ridges or keels moderately developed. Superior margin of orbit slightly raised, postorbital notch large. Eye large, rounded to slightly oval horizontally.
Lower lip narrow, ventral surface covered by short papillae with rounded tip, papillae slightly bigger close to dentary. Lower lip margin with long fringes; upper lip margin with short fringes. Maxillary barbel long, surpassing distal border of lower lip. Teeth bicuspid, long, mesial cusp greater and wider than lateral, dentary teeth larger than premaxillary. Premaxilla with 4(1) or 6*(2) teeth; dentary with 7(1), 8(1), or 9*(1) teeth. Sexual dimorphism on teeth not seen (see sexual dimorphism).
Body narrow and long, five lateral plate series in longitudinal rows below the dorsal fin: dorsal, mid-dorsal, median, mid-ventral, and ventral. Mid-dorsal series with moderately-developed keels, last plate extended around end of dorsal fin. Median and mid-ventral series with rough well-developed keels, last plate of mid-ventral series extended around end of anal fin. Median series bearing sensory canal between two proximate keels on caudal peduncle. Longitudinal lateral plates 30(1) or 31*(2); posterior plates of median series 15(1) or 16*(2). Lateral line complete. Lateral abdominal plates 6*(46). Lateral region of body posterior to pectoral-fin insertion between borders of lateral plates of mid-ventral series and of lateral abdominal plates with a slender area of naked skin, narrower than caudal peduncle width. Abdominal plates of posterior complex with a large well-defined preanal plate bordered by three plates and these by five plates. Abdominal plates of median complex parallel and longitudinally arranged in three series posteriorly and increasing series anteriorly. Abdominal plates of anterior complex irregular, smaller anteriorly, projected toward lip; anterior margin convex, surpassing horizontal line of anterior margin of cleithrum exposed ventrally.
Dorsal fin ii,7*(3), dorsal-fin spinelet present, locking mechanism not functional; depressed unbranched ray reaching fifth plate posterior to its base; depressed last branched ray reaching third or fourth plate. Pectoral fin i,6*(3); depressed unbranched ray evidently surpassing pelvic-fin origin; distal margin truncate. Pelvic fin i,5*(3); depressed unbranched ray slightly surpassing anal-fin origin; distal margin truncate. Anal fin i,5*(3); depressed unbranched ray reaching sixth plate posterior to its base; depressed last branched ray reaching third plate; distal margin truncate. Caudal fin damaged.
Color in alcohol. Based on lectotype and two paralectotypes, which are poorly preserved (color of specimens as originally described on “Remarks”). Dorsal surface of body with dark brown ground coloration; same coloring on ventral surface. Dorsal surface of head from anterior margin of snout to last dorsal plate with large and irregular dark brown spots in lectotype, small and irregular spots visible only on head in paralectotypes. Dorsal and pectoral fins with dark brown bars on rays, pectoral-fin membranes with dark brown pigments, caudal fin uniformly dark brown pigmented on branched rays and membranes, unbranched rays pale brown.
Sexual dimorphism. No sexual dimorphism is known.
Distribution and habitat. Rineloricaria cacerensis is known from its type locality, Cáceres, Mato Grosso State, Brazil ( Paraguay River basin). See additional comments on Discussion.
Remarks. The lectotype and paralectotypes of Rineloricaria cacerensis are highly discolored, with the caudal fin partly damaged not allowing confirmation of some characters related to caudal-fin filaments and color pattern mentioned in the original description of Miranda Ribeiro (1912). The caudal fin was described with the external rays prolonged as in R. parva , but with the superior one much longer. The dorsal overall ground coloration was described as light brown (pardo barrento); with a stripe on head around eyes; a blotch on predorsal region (triângulo cervical); five black transversal bands, the first around dorsal-fin insertion, the second broader and located posteriorly to dorsal fin, the remaining three equidistantly on caudal peduncle; caudal fin black, excepting outer rays; and dorsum of body from head to end of caudal peduncle with black spots, a series of larger black spots behind dorsal-fin insertion along midline. Unfortunately, the caudal fins of the lectotype and paralectotypes are broken, but we suspect that it should be interpreted as only the superior unbranched ray extended as a long filament. During this revision, we examined numerous species of Rineloricaria and found filaments on both unbranched rays only in R. parva , R. castroi , and R. wolfei . These species presents both caudal unbranched rays extended as long filaments of similar lengths (see Fig. 11 View Fig ), and also other characters as short sexually- dimorphic odontodes on lateral margins of the head and dorsum of pectoral fins in breeding male specimens, four lateral plate series, snout tip not raised, straight lateral margins of the head. However, species of Rineloricaria with five lateral plate series, snout tip raised, and lateral margins of the head curved anteriorly were found in R. fallax , R. formosa , R. hasemani , and R. morrowi . These species presented the caudal fin with only the superior unbranched ray extended as a long filament. Because R. cacerensis also present these charaters we suspect that the caudal fin could be similar. The stripes around eyes are no longer visible on lectotype and paralectotypes, and we do not know if they were arranged as vertical stripes or as horizontal cross bands.
Examined material. Type specimens: MNRJ 645 View Materials , lectotype, (1) , 67.7 mm SL, Cáceres , Mato Grosso (Aguas do Paraguay), Brazil. MNRJ 30030 View Materials , paralectotypes, (2) , 55.0- 56.4 mm SL, same data of lectotype .


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Order |
|
|
Family |
|
|
Genus |
Rineloricaria cacerensis
| Vera-Alcaraz, Héctor S., Pavanelli, Carla S. & Zawadzki, Cláudio H. 2012 |
Hemiloricaria cacerensis
| Ferraris, C. J. Jr. 2007: 245 |
| Isbrucker, I. J. H. & I. Seidel & J. P. Michels & E. Schraml & A. Werner 2001: 23 |
Rineloricaria cacerensis
| Britski, H. A. & K. Z. S. Silimon & B. S. Lopes 2007: 160 |
| Ghazzi, M. S. & O. T. Oyakawa 2007: 90 |
| Ferraris, C. J. Jr. 2003: 341 |
| Britski, H. A. & K. Z. S. Silimon & B. S. Lopes 1999: 130 |
| Isbrucker, I. J. H. & H. Nijssen 1976: 111 |
| Fowler, H. W. 1954: 113 |