
Geotrupacarus mycotrupetes (Krantz & Mellott)
|
publication ID |
https://doi.org/ 10.24349/4h94-gv61 |
|
persistent identifier |
https://treatment.plazi.org/id/070B87FB-FFC6-FFBA-FE14-30215FA9F878 |
|
treatment provided by |
Felipe |
|
scientific name |
Geotrupacarus mycotrupetes (Krantz & Mellott) |
| status |
|
Geotrupacarus mycotrupetes (Krantz & Mellott)
Scarabaeoid beetles worldwide serve as hosts for a variety of mites, and Mycotrupes is no exception. Geotrupacarus mycotrupetes ( Fig. 4 View Figure 4 ), a mite originally described as a member
of the genus Macrocheles by Krantz & Mellott (1968), is a large (>1,000 μm), distinctive phoretic form that attaches preferentially behind coxae I or in the gular area Myco of. gaigei
in northwestern peninsular Florida. Like its beetle host, G. mycotrupetes is arenicolous and burrows with ease through sand substrates. In laboratory studies, it was observed to use its formidable burrowing capabilities in locating Myco. gaigei when the beetles and the mites
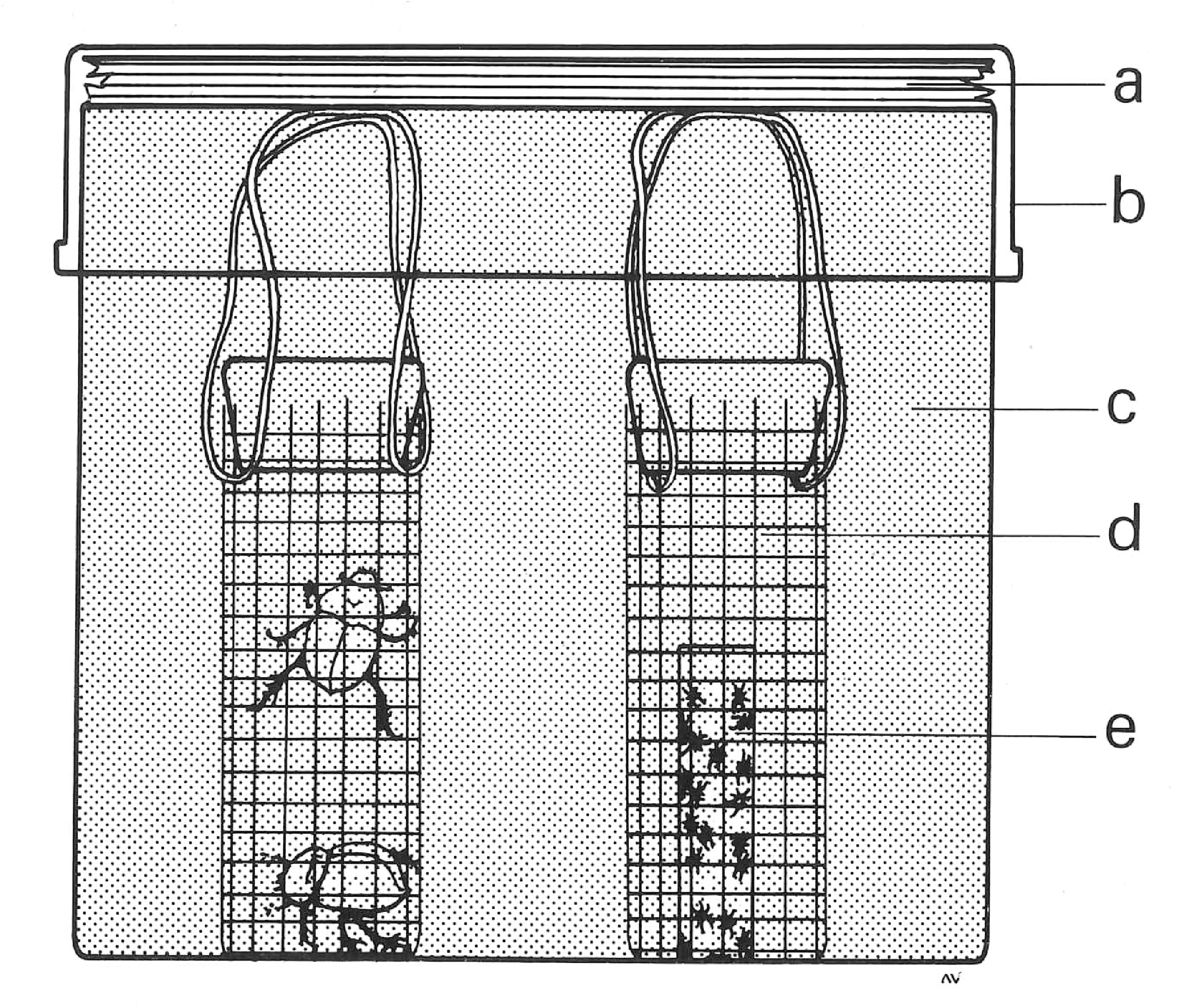
were sequestered in separate wire mesh cages beneath the sand surface ( Fig. 5 View Figure 5 ), presumably in response to recognition of a kairomonal cue generated by the beetle host ( Krantz & Royce 1994). Although it is clearly phoretic, the morphology and reproductive strategy G. of mycotrupetes
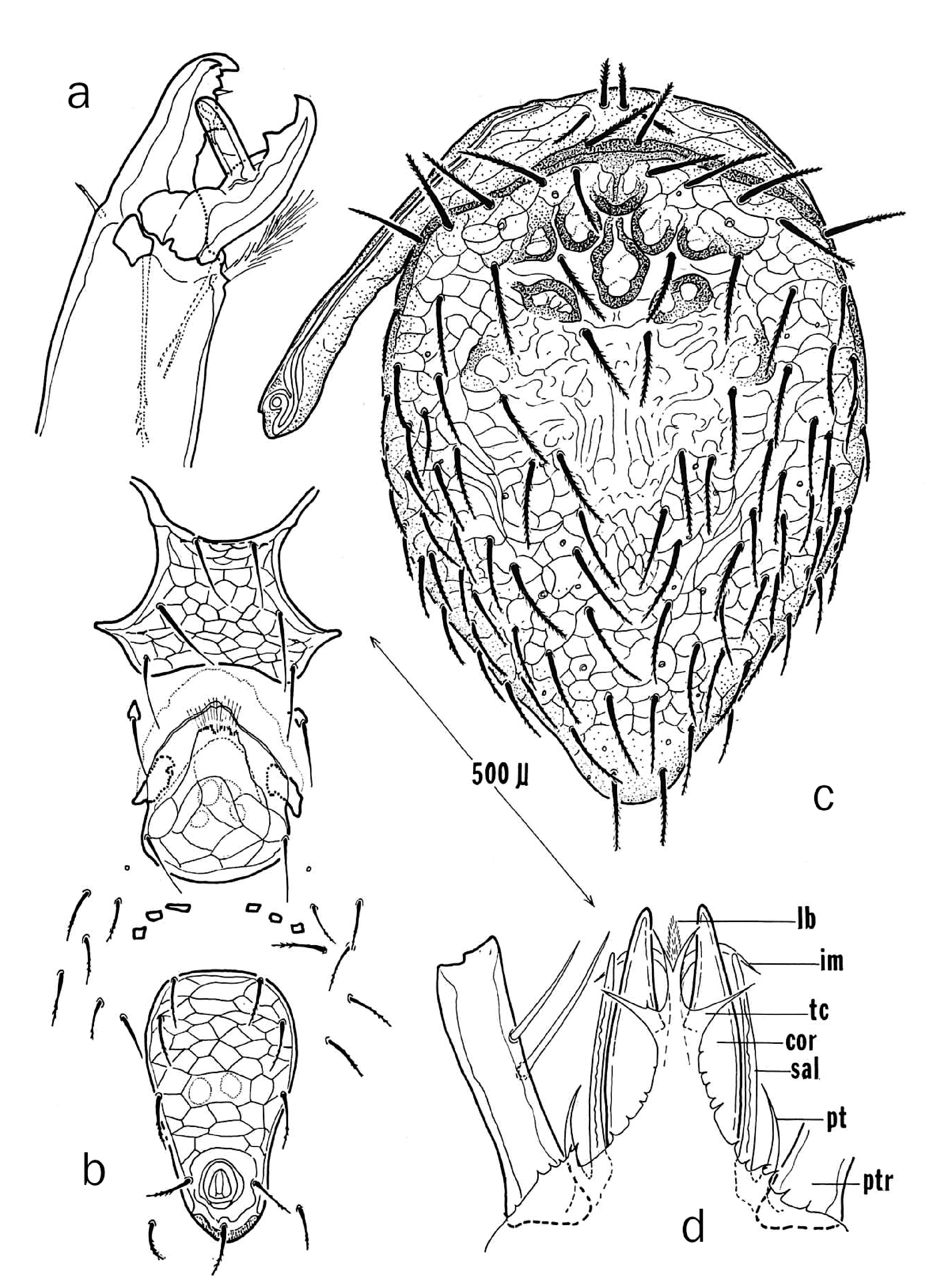
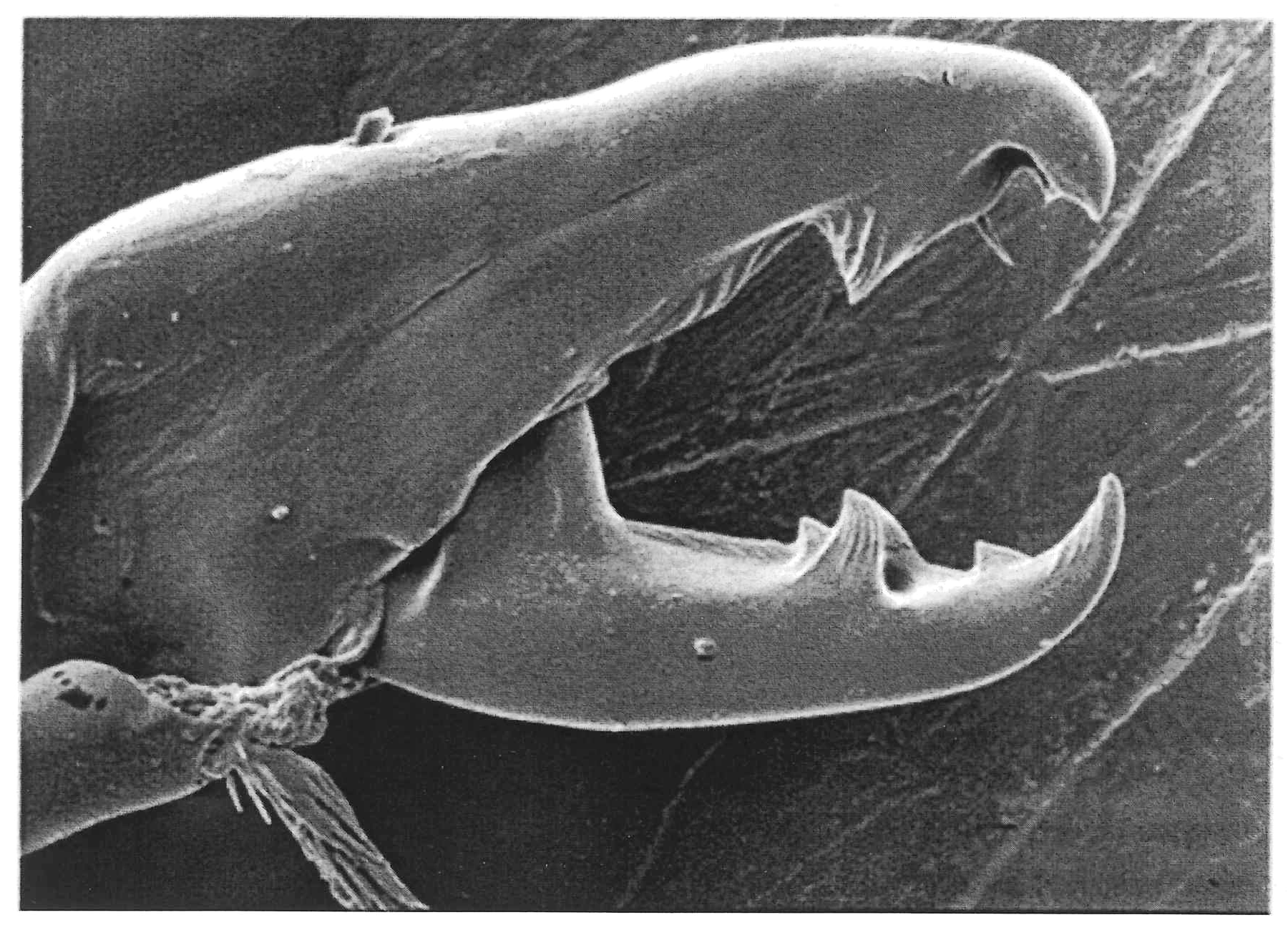
point to a close phylogenetic relationship with early derivative, free-living macrochelid taxa best exemplified by the genus Nothrholaspis Berlese ( Emberson 2010). Like Nothrholaspis , G. mycotrupetes lacks the subterminal bidentate tooth typically present on the cheliceral movable digit of more highly derived phoretic macrochelids, a structure coupled with an opposing ribbed platform on the fixed digit to form a complex for grasping a single hair of its host ( Fig. 6 View Figure 6 ). In addition, the lateral elements of the gnathotectum G of. mycotrupetes , like
those of Nothrholaspis species, are broadly fused to a bifurcated central element ( Fig. 4d View Figure 4 ) rather than being free and flaglike as in more highly derived forms. The female ventrianal shield ( Fig. 4b View Figure 4 ) is narrow and oblong rather than being subtriangular or broadly rounded as in Nothrholaspis species ( Emberson 2010), and the cribrum has narrow paranal extensions ( Krantz, 2009) (overly abbreviated in Krantz & Mellott, 1968, Fig. 4b View Figure 4 ) that are more typical
of early derivative lineages rather than being totally reduced to a narrow band of spicules confined to the posterior margin of the shield as in more highly derived macrochelids. Similar cribral extensions also occur in all postlarval immatures of G. mycotrupetes (Krantz 1990,
Krantz & Royce 1992). Postepigynal platelets, again characteristic of more primitive lineages,
ter of female; c – dorsal and peritrematic shields of female; d – prodorsal aspect of female (tc=gnathotectum, cor=corniculum, lb=labrum, im=internal mala, sal=salivary stylus, pt=patrocinium, ptr=palptrochanter I) (from Krantz & Mellott 1968) (courtesy J. Kansas Entomol.
Soc.).
are present in the female ( Krantz 1998). Among the less obvious morphological features that define both described species of Geotrupacarus ( G. mycotrupetes and peltotrupetes) are the infrequent and often obscure barbs that ornament the subcapitular and ventral shield setae. Their obscurity is a consequence of some barbs being so closely aligned to their given seta that they are easily overlooked, as sometimes occurred in the original species descriptions ( Krantz & Mellott 1968). Finally, the reproductive strategy of G. mycotrupetes is more suggestive of
a sand barrier (a=moistened paper towel layer, b=carton lid, c=sand, d=wire cylinder, e=shell vial) (after Krantz & Royce 1994).
primitive diplodiploidy or parahaploidy sensu ( Norton et al. 1993) than of the arrhenotokous form of haplodiploidy common to most insect-associated phoretic macrochelids ( Krantz & Royce 1994).
Overall, the primitive characteristics of G. mycotrupetes , coupled with its unusual phoretic proclivities, lends credence to the concept that phoresy has arisen more than once in the Macrochelidae (Krantz & Royce 1992) , and at the same time raises the question as to whether a phoretic association with a non-vagile host like Myco. gaigei could involve something other than transport to a fresh food substrate.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.