
Physoschistura mango, Conway & Kottelat, 2023
|
publication ID |
https://doi.org/ 10.26107/RBZ-2023-0051 |
|
publication LSID |
lsid:zoobank.org:pub:EA56D49D-82C6-4135-B177-C812E6C657E2 |
|
persistent identifier |
https://treatment.plazi.org/id/042F87C3-FFE2-FF80-FC38-F84F7228F95B |
|
treatment provided by |
Felipe |
|
scientific name |
Physoschistura mango |
| status |
sp. nov. |
Physoschistura mango , new species
( Figs. 1 View Fig –11)
Petruichthys sp. (Hopong) – Kano et al. (2022). Holotype. MHNG 2790.086 View Materials , male, 18.5 mm SL; Myanmar: Shan State: pool at foot of a hill near Hopong, located 30 km to the east of Taunggyi; Kamphol Udomritthiruj (from aquarium-fish collectors), July 2008.
Paratypes. ZRC 65321 View Materials , 2 View Materials , 16.3–16.9 mm SL; CMK 20748, 5 , 17.1 –22.0 mm SL; collected with holotype .
Non-type material. TCWC 20683.01, 2 (C&S), 19.0– 23.5 mm SL; TCWC 20684.01, 1 ( SEM), 21.7 mm SL; TCWC 20685.01, 1 (DNA voucher), 20.1 mm SL; locality unknown (aquarium trade).
Diagnosis. Physoschistura mango is distinguished from congeners by features of the adult sexually dichromatic colour pattern, comprising a solid dark brown to black horizontal stripe along body side (distinct in mature males only), and a series of small spots or irregular markings in females (vs. colour pattern not known to be sexually dichromatic, comprising a series of vertical bars, most obvious on posterior half of body), and by having a shorter lateral-line canal on the body (5–6 pores vs. 5–10 in Ph. brunneana , 29 in Ph. raoi , 56–57 in Ph. rivulicola , 16–37 in “ Ph. ” pseudobrunneana ). It is further distinguished by a number of reductive features of the cephalic lateral-line canal system, including a shorter infraorbital canal, with 5–7 pores, restricted to a short series of ossicles associated with the ventral edge of the lachrymal (vs. extending around entire ventral margin of orbit, with 11–13 pores in Ph. brunneana , 11 in Ph. raoi , 10 in Ph. rivulicola and “ Ph. ” pseudobrunneana ), a shorter preoperculo-mandibular canal, restricted to preopercle only, with 2–4 pores (vs. preoperculo-mandibular canal present on both dentary and preopercle, with 9–10 pores in Ph. brunneana , 8 in Ph. raoi , 9 in Ph. rivulicola , and 7 in “ Ph. ” pseudobrunneana ). It is further distinguished from Ph. brunneana and Ph. raoi by having 8+8 principal caudal-fin rays (vs. 9+8), from P h. r aoi and Ph. rivulicola by the absence (vs. presence) of the axillary pelvic-lobe, and from “ Ph. ” pseudobrunneana by the absence (vs. presence) of a suborbital flap in males.
Physoschistura mango is also distinguished from the potentially closely related, and externally similar, Petruichthys brevis by the absence (vs. presence) of a suborbital flap in males, by having a shorter lateral-line canal on the body, with 5–6 pores (vs. 12 pores), and by a number of reductive features of the cephalic lateral-line canal system, including a shorter infraorbital canal, with 5–7 pores, restricted to a short series of ossicles associated with the ventral edge of the lachrymal (vs. with 11 pores, extending around entire ventral margin of orbit), by having the infraorbital canal separate from the otic canal (vs. connected), and by a shorter preoperculo-mandibular canal, restricted to preopercle only, with 2–4 pores (vs. preoperculo-mandibular canal present on both dentary and preopercle, with 9 pores). In life, Ph. mango is distinguished from Pe. brevis by features of the adult male nuptial colour pattern, including a solid orange-reddish background colour (vs. salmon to light cream background colour).
Description. Description based on type material, unless noted. See Figs. 1 View Fig and 2 View Fig for general appearance and Table 1 for select morphometric characters. A moderately elongate nemacheilid with body depth gradually increasing up to point slightly anterior to dorsal-fin origin, then decreasing below dorsal-fin base, and then almost uniform until caudal-fin base. Dorsal profile with a shallow concavity at nape. Head slightly compressed, as deep as anterior part of body, deeper than caudal peduncle; body slightly compressed anteriorly to compressed posteriorly. Eye large, diameter greater than snout length. Margin of orbit visible in both dorsal and ventral views. In lateral view, eye flush with dorsal profile of head. Snout short, ethmoid region raised, dome-like in dorsal and lateral views. Mature males without suborbital flap ( Fig. 3A View Fig ). Anus closer to anal-fin origin than base of pelvic fin. Caudal peduncle without dorsal or ventral keel, its depth uniform, 1.6 times in its length. Largest recorded size 23.5 mm SL (non-type material).
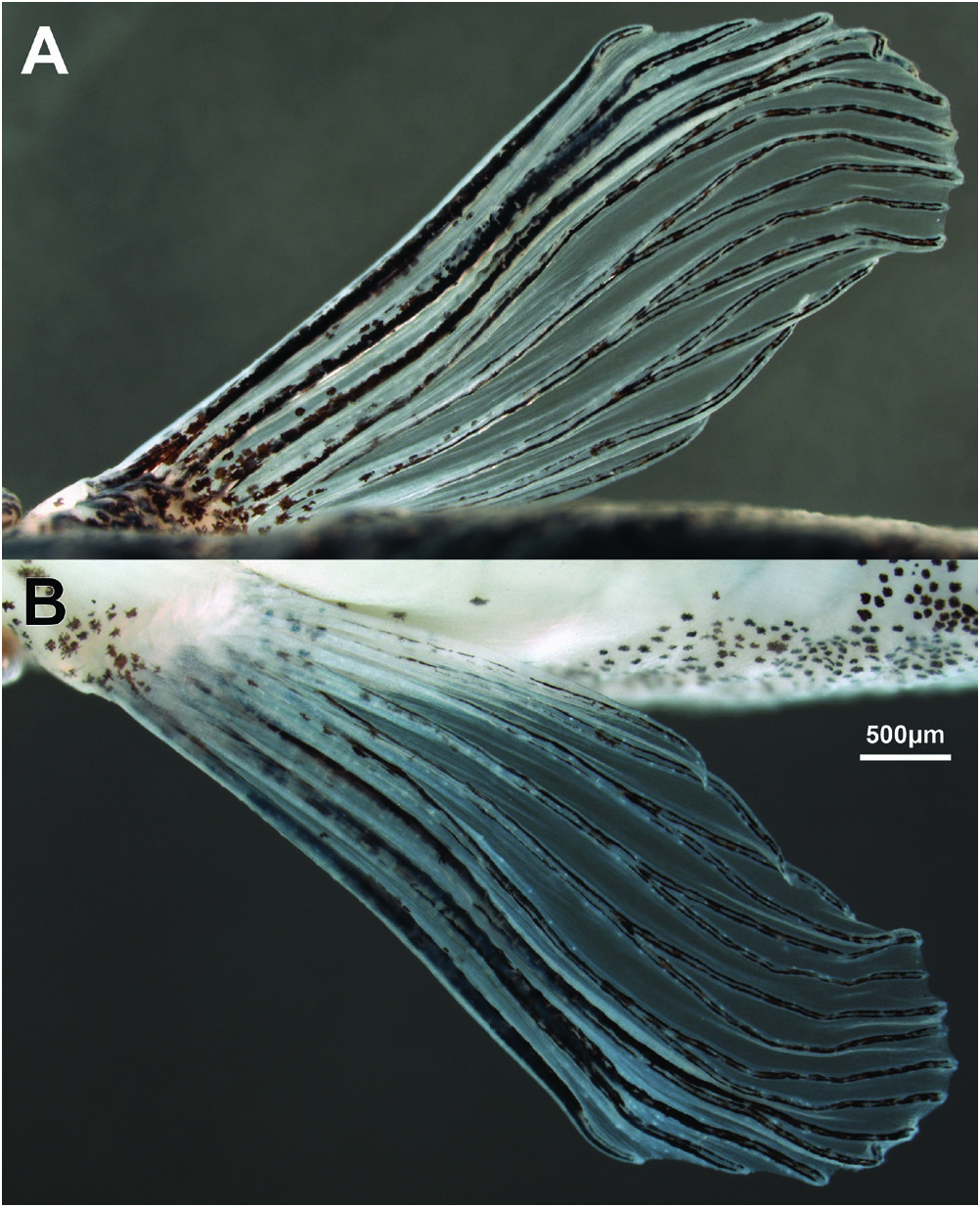
Following counts obtained from non-type C&S material (TCWC 20683.01). Dorsal fin with 3–4 unbranched and 8½ branched rays (iii-iv,8½); distal margin straight to slightly convex; first branched ray longest. Anal fin with 3 unbranched and 5½ branched rays (iii,5½); distal margin slightly concave. Caudal fin with 8+8 principal rays; 7–8 dorsal procurrent rays, 5–6 ventral procurrent rays; forked, lobes rounded, upper lobe 1.2 times length of lower lobe, 1.4 times length of median rays. Paired fins large. Pectoral fin with 1 unbranched, 8–9 branched rays, and 1 small unbranched ray (i,8–9,i); triangular, with obvious sexual dimorphism in fin size when pressed against body side, reaching past imaginary vertical line through base of second branched dorsal-fin ray in males, not reaching past imaginary vertical line through origin of dorsal fin in females; configuration of anterior branched pectoral-fin rays also sexually dimorphic (see under sexual dimorphism for further details). Axillary pectoral lobe absent. Pelvic fin with 1 unbranched, 6 branched, and 1 small unbranched rays (i,6,i); reaching mid-way between anus and anal-fin origin when pressed against body; posterior margin slightly convex; origin along imaginary vertical line through base of 4–5 th branched dorsal-fin ray; axillary lobe absent.
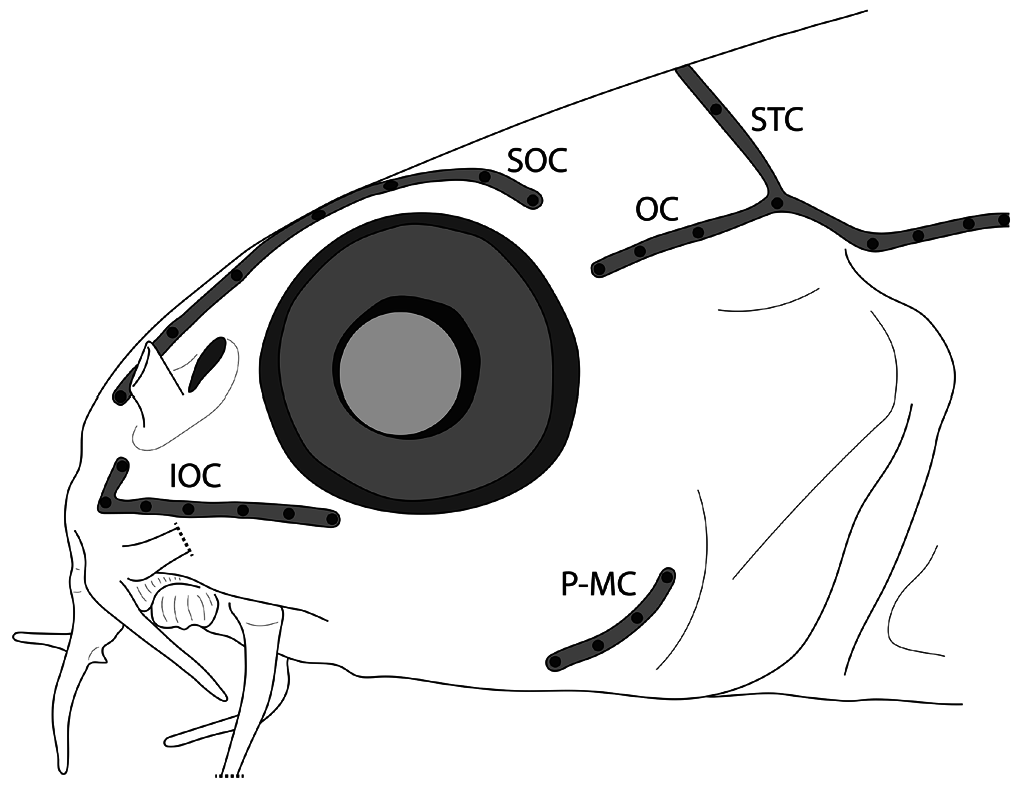
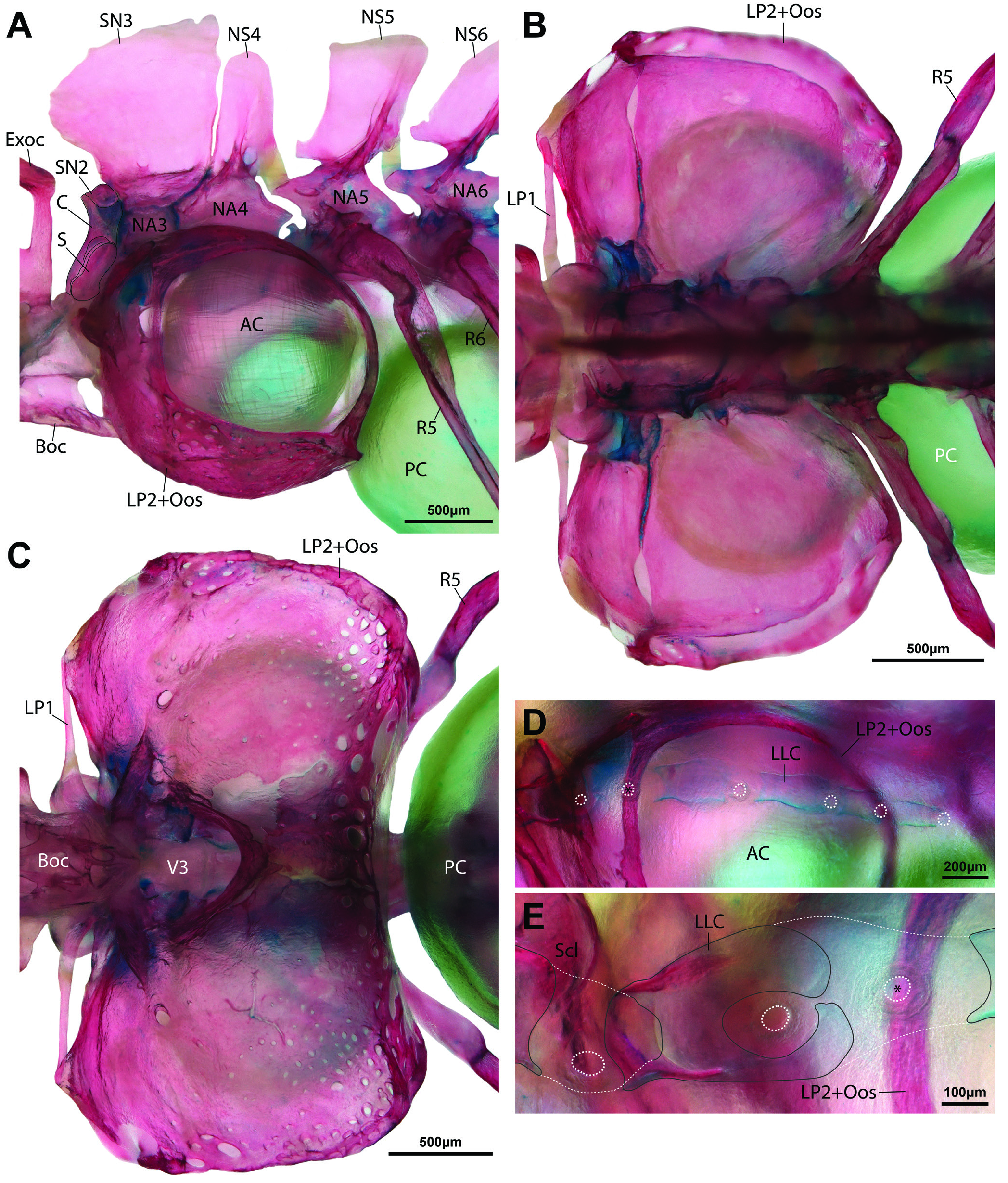
Body covered by tiny cycloid scales, excluding ventral surface between pectoral fins. Lateral-line canal on body incomplete, very short, with 5–6 pores only. Lateral-line canal on body supported by short, tube-like ossifications; two anteriormost larger than preceding ossifications ( Fig. 10D, E View Fig ). Cephalic lateral-line canal system with 7 supraorbital, 3–4 otic, 5–7 infraorbital, 2–4 preoperculo-mandibular, and 2+2, 3 or 5 supratemporal pores ( Fig. 3A View Fig ). Preoperculo-mandibular canal restricted to preopercle ( Fig. 9B). Infraorbital canal (=A2 portion of infraorbital canal of Kottelat, 1990) restricted to snout, with 5–7 pores; canal formed by one or two ossicles ventral to ribbon-like lachrymal, anteriormost with dorsally directed tubule anteriorly ( Fig. 9A). Otic canal (=A1 portion of infraorbital canal of Kottelat, 1990) disjunct from infraorbital canal, with 3–4 pores; connected posteriorly to supratemporal canal on both sides of head in 1 of 2 C&S specimens; connected on left side only in 1 C&S. Supratemporal canal interrupted medially in 1 paratype, with 2 pores on each side (2+2); complete across midline, with 3 or 5 pores in other material.
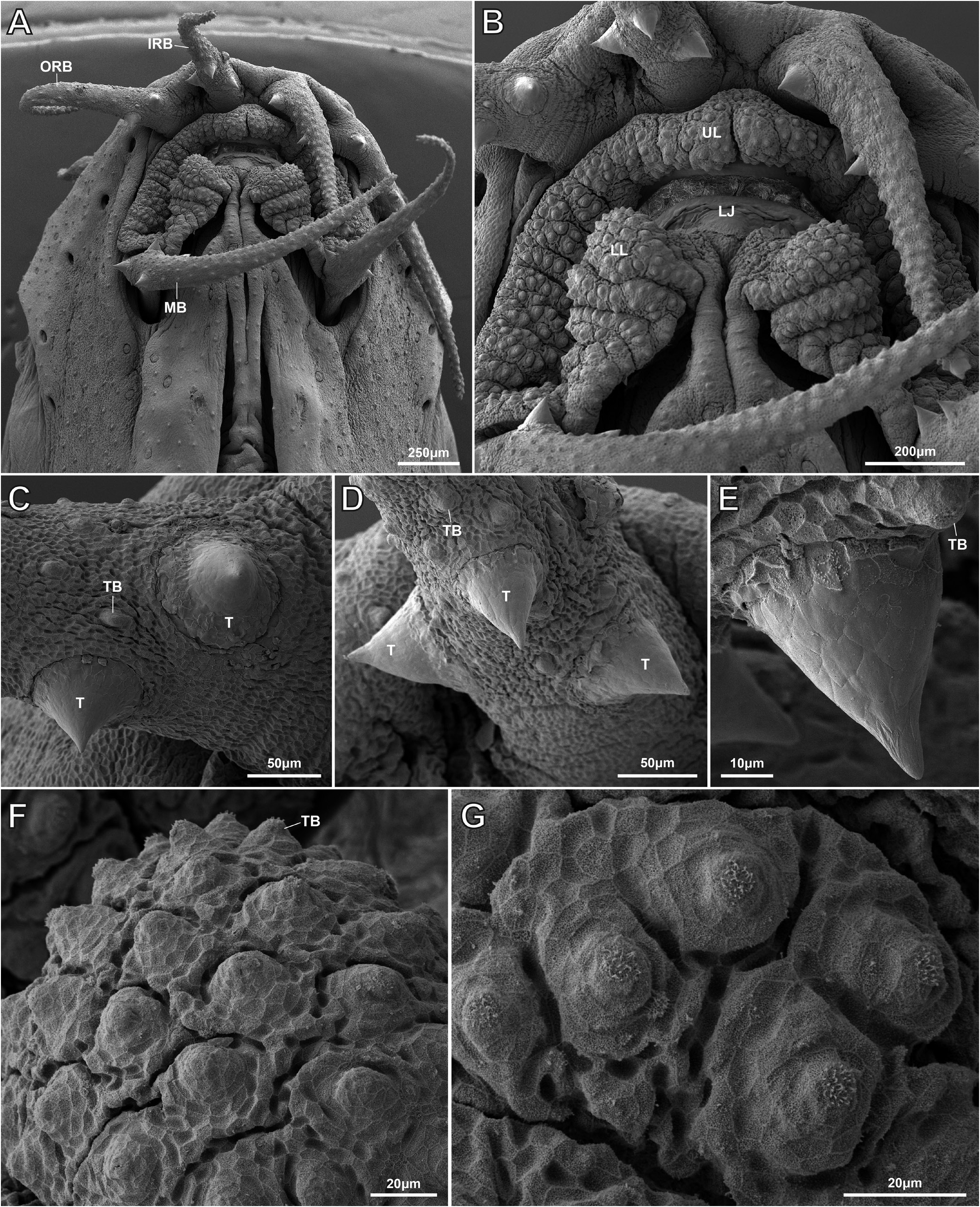
Anterior nostril pierced at tip of a short, anterodorsally directed tube. Posterior nostril large, adjacent to base of anterior nostril tube. Mouth arched, gape 1.3 times wider than long ( Fig. 4A, B View Fig ). Lips papiliferous; each papilla bearing a single tastebud at apex ( Fig. 4B, F, G View Fig ). Upper lip without centre notch ( Fig 4B View Fig ). Processus dentiformis well developed, hidden in ventral view by upper lip. Lower lip thicker than upper lip, with V-shaped interruption at centre; area of lip adjacent to interruption swollen, forming a triangular cushion, with 2–3 shallow folds, forming crenate lateral margin ( Fig. 4A, B View Fig ). Tip of lower jaw exposed by V-shaped median interruption in lower lip. Margin of lower jaw rounded, without median notch or concavity. Inner rostral barbel reaching past imaginary vertical line through anterior margin of eye, not reaching to imaginary vertical line through centre; outer rostral barbel reaching to imaginary vertical line through centre of eye. Maxillary barbel reaching to or slightly past imaginary vertical line through posterior margin of eye. Surface of all barbels covered with tastebuds ( Fig. 4C–E View Fig ).
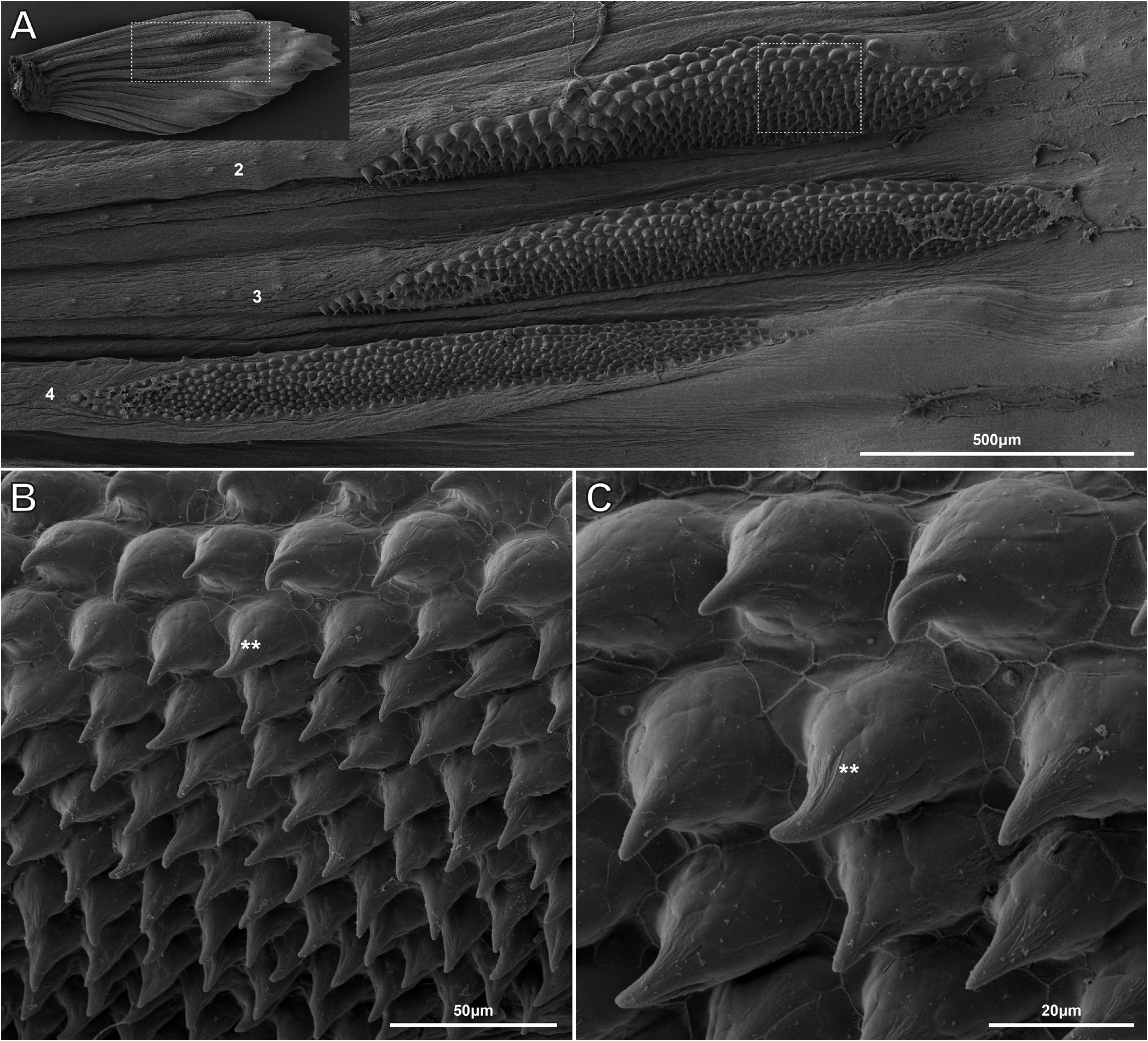
Following information based on non-type material (TCWC 20683.01). Intestine with bend posterior to stomach (Fig. 11). first branched ray; membrane between branches of second branched ray reduced in both sexes. Membrane between distal part of third and fourth branched ray in male wider than between fourth and fifth due to anterior displacement of third branched ray ( Fig. 5 View Fig ). First to third branched rays of male with an elongate patch of tiny tubercles on dorsal surface. Examination of a single mature male (TCWC 20684.01) using SEM revealed each elongate patch of tubercles to comprise 9–10 mediolateral rows of 20–50 tubercles, with a weak gradient in tubercle height from anterior to posterior within each patch (ranging from ~50 µm in anterior rows to ~30 µm in posterior rows) ( Fig. 6 View Fig ). Tubercles located within anteriormost rows of anteriormost patch (associated with first branched ray) appear larger than tubercles located within anteriormost rows of the two posterior patches (associated with second and third branched rays) ( Fig. 6 View Fig ). In male examined with SEM, 2–4 larger conical tubercles (~50–60 µm in height), each with pointed tip, located on inner face of each barbel, close to base ( Fig. 4C–E View Fig ). Similar tubercles not observed in other males (n=5) and their presence in a single individual may be associated with reproductive condition. No tubercles observed on other body surfaces or other fins in males. No tubercles observed in females.
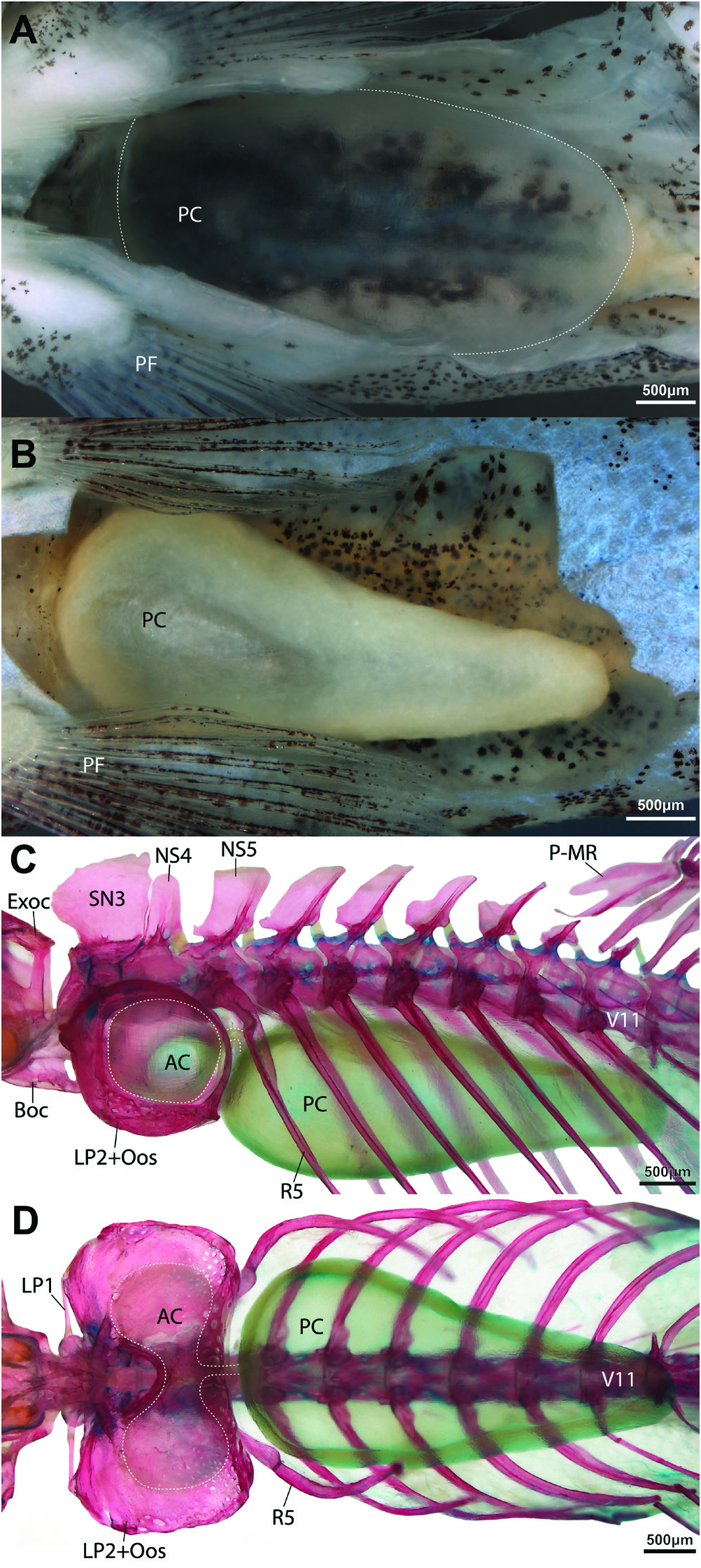
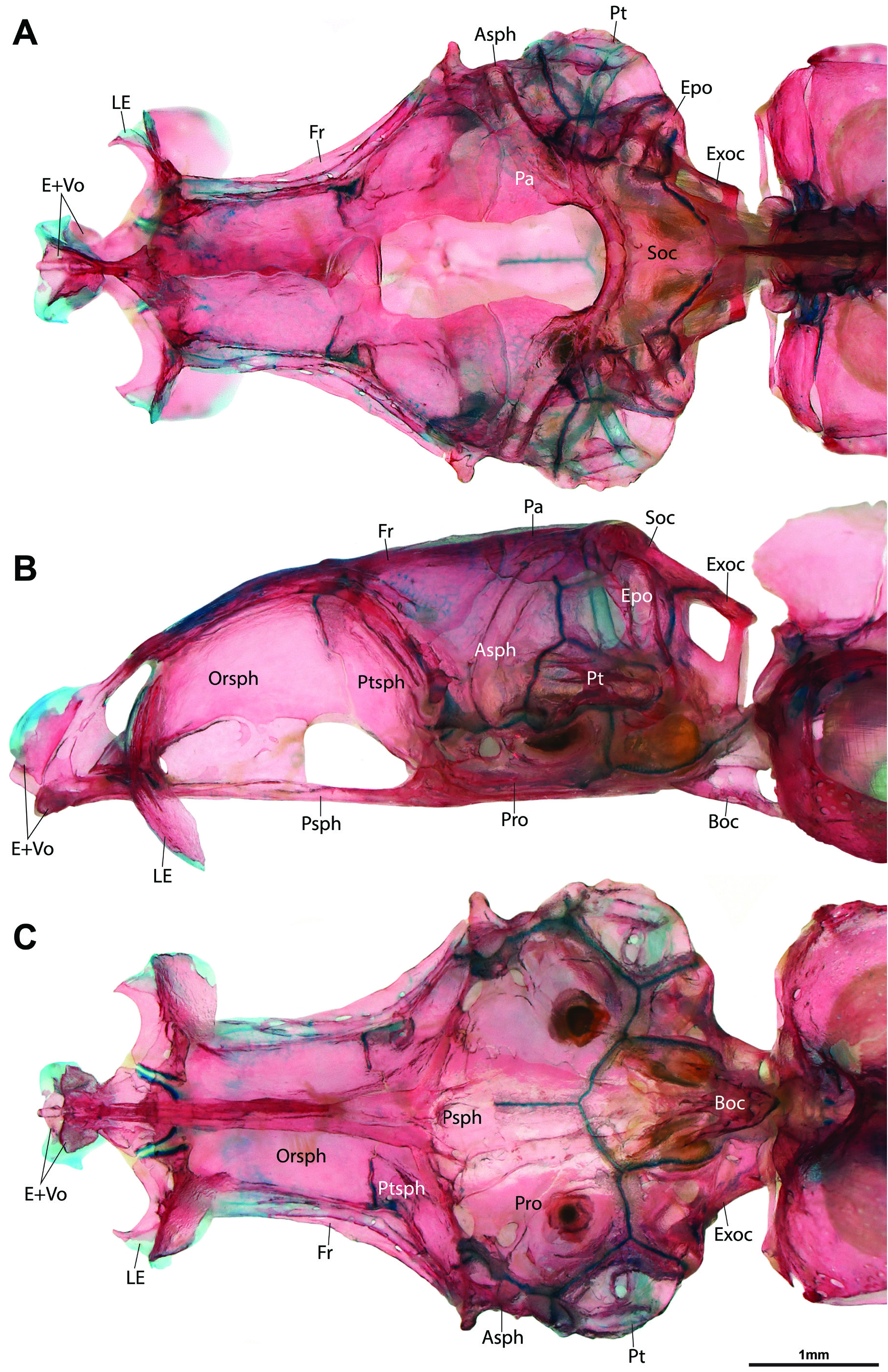
Anterior chamber of swimbladder bilobed, surrounded by bony swimbladder capsule ( Fig. 7C, D View Fig ). Posterior chamber of swimbladder large, occupying most of dorsal region of abdominal cavity in both sexes ( Fig. 7A, B View Fig ). Postepiphyseal fontanelle well-developed ( Fig. 8A View Fig ). Mesethmoid and vomer fused ( Fig 8 View Fig ). Preethmoid absent; preautopalatine and second preethmoid present ( Fig. 9B, C). Branchiostegal rays 3 ( Fig. 9F). Ceratobranchial 5 with ~10 conical pharyngeal teeth, arranged in single row around posteromedial edge of bone; 4 lowermost teeth approximately twice as large as other teeth ( Fig. 9D). Gill rakers present along posterior edge of ceratobranchial 1, anterior and posterior edge of ceratobranchial 2-4, and anterior edge of ceratobranchial 5; small, triangular fleshy projections, without obvious ossification ( Fig. 9D). Gill rakers absent from anterior edge of ceratobranchial 1. Total number of vertebrae 32 or 33, comprising 16 or 17 abdominal and 16 caudal vertebrae. Ribs not symmetrically developed, 9(left)/8(right) or 10(left)/9(right). Insertion of first dorsal-fin pterygiophore between neural spines of vertebrae 9/10. Insertion of first anal-fin pterygiophore between hemal spines of vertebrae 20/21 or 21/22. Caudal-fin skeleton with 5 hypurals.
Sexual dimorphism. Following information based on non-type material. Pectoral fin longer in males than in females, more pointed, more rigid and expanded laterally. First pectoral-fin branched ray (second pectoral-fin ray) of males thicker than unbranched ray (first pectoral-fin ray) and other branched rays ( Fig. 5 View Fig ). Membrane between branches of first branched ray reduced in both sexes. Distal part of second and third branched ray displaced anteriorly in male, approaching Size and shape of posterior chamber of swimbladder markedly different between two dissected specimens, one male, one female (TCWC 20683.01). In male, posterior chamber of swimbladder occupying most of abdominal cavity, rounded posteriorly, near-uniform diameter along its length ( Fig. 7A View Fig ). Wall of chamber in male also thin, appearing transparent. Posterior chamber in female smaller than that of male, exhibiting distinct reduction in diameter towards posterior (diameter of posterior region approximately half that of anterior region) ( Fig. 7B View Fig ). Wall of chamber in female opaque, thicker than that of male.
Colouration. In preservative, body background colour light cream to white ( Fig. 1 View Fig ). Darker pigment features faint in holotype and paratype series ( Fig. 1A, B View Fig ), including series of dark brown to black spots along dorsal surface, some extended ventrally to form short vertical bars; a horizontal row of dark brown to black spots or small irregular markings along body side, level with horizontal septum. Additional dark brown to black spots or small irregular markings along ventrolateral part in some females. A dark brown to black spot on caudal-fin base, slightly below midline, covering base of uppermost principal rays of lower lobe (weakly developed in holotype and paratype series; Fig. 1A, B View Fig ). Fin rays with weak scattering of dark brown to black pigment in all fins, most pronounced along first (unbranched) ray of pectoral fin in male. Dorsal and lateral surface of head scattered with dark brown to black pigment, most evident on snout where pigment forms a faint stripe anterior to eye. In mature males (non-type material; Fig. 1C View Fig ), markings of horizontal row fused together to form a solid horizontal stripe, similar in depth to pupil diameter.
Following information based on non-type material. In life ( Fig. 2 View Fig ), body semi-translucent, background colour light creamy brown to white in female, pale yellow to light orange in males. Features of pigmentation on body and head described above appear light brown in female ( Fig. 2B View Fig ), darker brown in male ( Fig. 2A View Fig ). In female, outermost principal caudal-fin rays appear light yellow, rays of other fins and membranes between rays of all fins transparent ( Fig. 2B View Fig ). In male, rays of all fins appear light orange, membranes between rays transparent ( Fig. 2A View Fig ). Based on Kano et al. (2022: fig. 3E), a specimen collected from Hopong and photographed when still alive exhibited a semi-translucent body, with a pale orange background colour in life. The markings on the dorsal and lateral surface of the body in this individual (similar to those of the holotype and paratypes; Fig. 1A, B View Fig ) appear light brown.
Distribution and habitat. The type locality is a small pool, near the town of Hopong, located 30km to the east of Taunggyi, Shan State, Myanmar. The type material was part of a batch of aquarium fishes exported from Myanmar to Thailand; the collector stated that they had been caught at the type locality of Danio margaritatus ( Roberts, 2007) . Roberts (2007: 138) reported that with D. margaritatus occurred “a small species of the nemacheiline loach genus Yunnanilus ”, which was likely Physoschistura mango . If true, the type locality is figured by Roberts (2007: fig. 5). Based on COI sequence data (see below), Ph. mango is the species reported from Hopong by Kano et al. (2022) and referred to as Petruichthys sp. (Hopong). In addition to Ph. mango, Kano et al. (2022 : table 1) report 24 other species of freshwater fishes from Hopong, including three other nemacheilids ( Petruichthy brevis , Physoschistura brunneana , and Physoschistura rivulicola ).
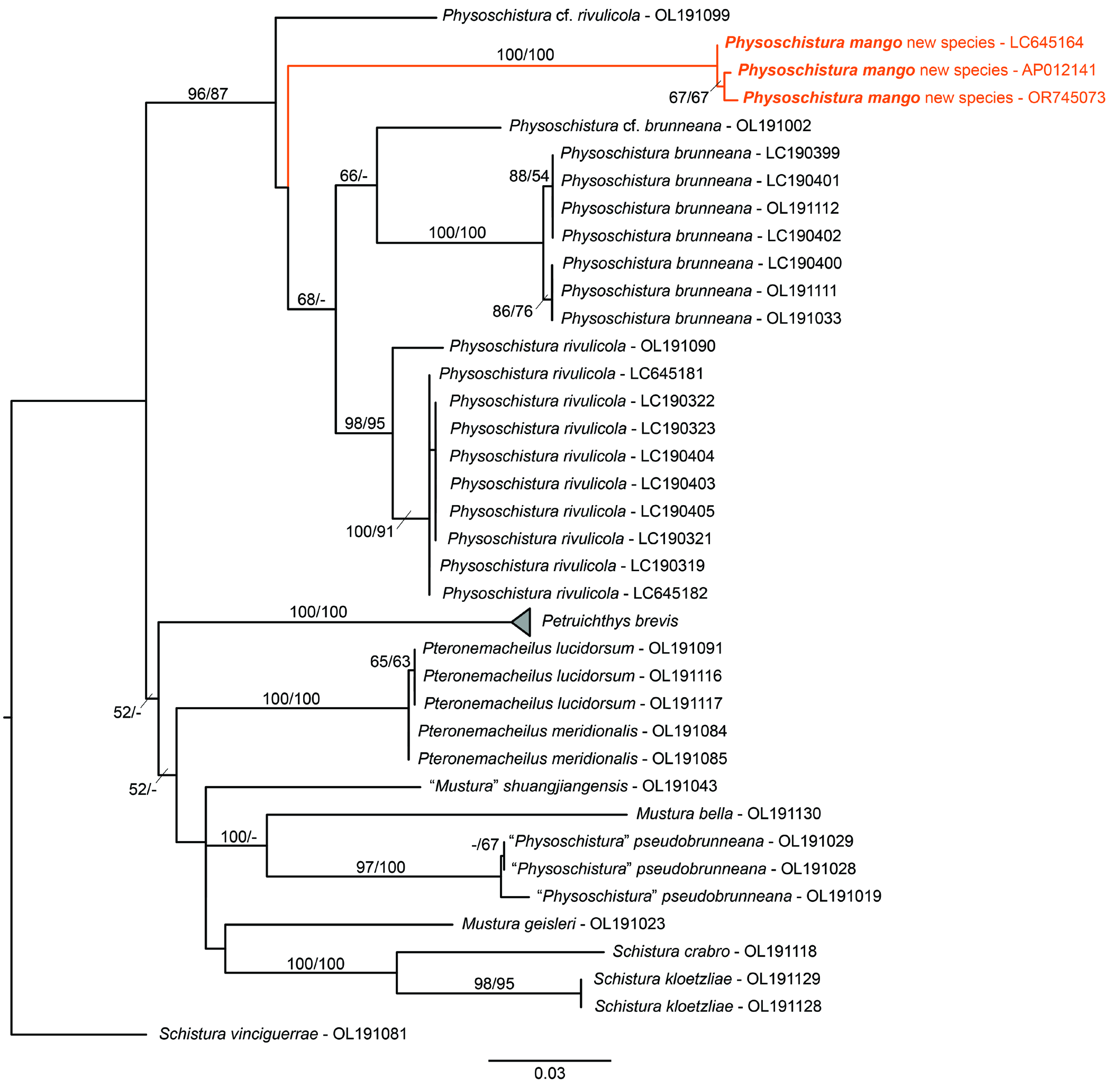
COI barcode. We obtained one 665 bp segment of the COI gene from an individual of Physoschistura mango obtained via the aquarium trade (non-type material, TCWC 20685.01; GenBank OR745073). This sequence is highly similar (as determined via Blast search) to two other COI sequences currently available on GenBank, including AP012141.1, obtained from an aquarium specimen identified as Yunnanilus sp. “rosy loach” (M. Miya, unpublished), and LC645164.1, obtained from an individual referred to as Petruichthys sp. (Hopong) by Kano et al. (2022). The mean uncorrected p-distance between the three sequences is 0.4%. Based on this information, we conclude that the new species, Petruichthys sp. (Hopong) of Kano et al. (2022) and the “Rosy” loach of the aquarium hobby are conspecific. The mean uncorrected p-distance between Ph. mango , four other species of Physoschistura and Petruichthys ranged between 8.7–12.6% ( Table 2).
Etymology. Mango, the tree Mangifera indica in reference to the orange-yellow flesh of its fruit. A noun in apposition.
Notes on biology. There is no information on biology in the wild. In aquarium (KWC, pers. obs.; Dederer, 2023), individuals swim actively in the lower part of the water column between short periods of rest on the substrate (while most nemacheilids dwell on the substrate). Mature males frequently chase each other and spar aggressively, involving nipping of the body and fins. Successful spawning and rearing have occurred in a densely planted aquarium ( Dederer, 2023).
Remarks. The new species exhibits the majority of the diagnostic characters of Physoschistura listed in the original description by Bănărescu & Nalbant (in Singh et al., 1982). This includes an incomplete lateral line, not reaching beyond the dorsal fin (lateral line with 5–6 pores only in Ph. mango ); a forked caudal fin (weakly forked in Ph. mango ); dorsal fin with 8 or 9 branched rays (8½ in Ph. mango , equal to 9 of Bănărescu & Nalbant); feeble to moderately developed processus dentiformis on upper jaw (moderate in Ph. mango ; Fig. 4B View Fig ); two halves of the swimbladder capsule joint and coalescent on their inner face (not connected by a manubrium) (see Figs. 7D View Fig , 10C View Fig ); and posterior chamber of swimbladder well developed, free, more or less conical, in direct contact with the capsule (see Fig. 7C,D View Fig ; in Ph. mango the well-developed posterior chamber of the swimbladder is separated from the posterior face of the swimbladder capsule by a short gap). The last character listed as diagnostic for Physoschistura by Bănărescu & Nalbant (in Singh et al., 1982: 208) relates to pigmentation: “body with brownish crossbars.” Physoschistura mango does not exhibit crossbars (which we interpret as vertical bars), and instead exhibits a sexually dichromatic colour pattern that is quite different from the other species currently placed in Physoschistura and more reminiscent of Petruichthys brevis ( Fig. 12 View Fig ). This includes a horizontal row of dark brown to black blotches along the centre of the body, more prominent in males, especially in mature males, which develop a solid horizontal stripe, likely formed via fusion of the blotches or darkening of the intervening pigment.
Kottelat (1990) expanded the diagnosis for Physoschistura to also include characters of the mouthparts (“mouth strongly arched, 1.5–2.0 times wider than long; lower lip with a median interruption forming two laterally broadly triangular pads with deep furrows”). These characters, especially the “triangular pad”, appear to have been misinterpreted or misunderstood by many authors (see Kottelat, 2018), and this character was later amended to “lower lip with wide median interruption, the two halves forming an acute angle, not in contact medially, wide and fleshy medially, forming a more or less triangular cushion, partly free from jaw and connected to isthmus by a frenum” ( Kottelat, 2018: 290, fig. 2). This arrangement of the lower lip is not unique to Physoschistura , and is found also in the members of Mustura , Pteronemacheilus , Protonemacheilus , and also Petruichthys , though in the later the triangular cushion is traversed with deep folds ( Kottelat, 2018: fig. 9). Physoschistura mango exhibits the aforementioned condition ( Fig. 4A, B View Fig ), and like Pe. brevis also exhibits deep folds across the surface of the triangular cushion.
Although Physoschistura mango is similar in overall appearance to Petruichthys brevis ( Fig. 12 View Fig ), similar enough to be considered congeners by Kano et al. (2022), the new species appears to be more closely related to species of Physoschistura than to Pe. brevis . Both ML and MP analyses of our COI dataset ( Fig. 13 View Fig ) recover Ph. mango as a member of a clade that includes also Ph. brunneana (type species of Physoschistura ), Ph. rivulicola , and the potentially undescribed Ph. cf. rivulicola and Ph. cf. brunneana (reported by Dvořák et al., 2022). Though branch support for this clade was high (87–96% bootstrap support), the relationships within are largely unresolved. We note here that all members of this clade are found within or in the vicinity of Lake Inle, and water bodies of the surrounding He-Ho plain, Shan State, Myanmar. The position of Pe. brevis , also from this area of Myanmar, is unresolved based on our analyses ( Fig. 13 View Fig ) (though see Dvorák et al. [2022, 2023] for a wellsupported sister group relationship between Pe. brevis and the aforementioned Physoschistura based on a larger dataset). As reported earlier by Dvořák et al. (2022, 2023), we did not recover “ Physoschistura” pseudobrunneana as a close relative of the other members of Physoschistura included in our data set, rendering Physoschistura polyphyletic as currently recognised ( Fig. 13 View Fig ). It is clear based on the results presented herein, and those of other recent molecular phylogenetic studies on South Asian nemacheilids (e.g., Dvořák et al., 2022, 2023), that additional work is needed to refine the membership of Physoschistura , and the closely related genera Mustura , Pteronemacheilus , and Petruichthys . This work will likely require the erection of multiple new genera (e.g., see Kottelat, 2018) but is beyond the scope of the present study.
An interesting, potentially sexually dimorphic feature of Physoschistura mango that deserves further investigation is the presence of tubercles at the base of the barbels ( Fig. 4A–E View Fig ). We observed barbel tubercles only in a single nuptial male (TCWC 20684.01, male, 21.7 mm SL), which also had well-developed patches of tiny tubercles on the anteriormost branched pectoral-fin rays ( Fig. 6 View Fig ). The absence of barbel tubercles in other males (and females) that we the number of miniature species that have been described from this region in the course of the last three decades (e.g., Kullander & Britz, 2002; Britz, 2003, 2009; Kottelat et al., 2006; Roberts, 2007; Britz & Kottelat, 2008; Britz et al., 2009, 2012, 2021, 2022; Conway & Kottelat, 2011; Conway et al., 2011; Ou et al., 2011; Anoop et al., 2019; Parenti et al., 2023). Miniature fishes typically exhibit reductions in the lateral-line system ( Myers, 1958; Weitzman & Vari, 1988); the reductions that we have documented in this system for Ph. mango , in comparison with larger bodied close relatives, including other members of Physoschistura and potentially also Pe. brevis , may be linked to the process of miniaturisation ( Hanken & Wake, 1993). Fig. 11. Physoschistura mango, TCWC 20683.01, male, 23.4 mm
SL; digestive tract.
examined may be related to reproductive condition (i.e., barbel tubercles are present only in reproductively active males), or less likely, an aberration in a single individual. To the best of our knowledge, this is the first report of barbel tubercles in a nemacheilid, and only the second record within Cypriniformes , where they are known only from some members of the genus Psilorhynchus (Conway et al., 2013) . A second interesting, and potentially sexually dimorphic feature of Ph. mango involves the size and shape of the swimbladder (larger and with a rounded posterior part in male vs. smaller and with a pointed posterior part in female; Fig. 7A, B View Fig ). Sexual dimorphism of the swimbladder is rare in cypriniform fishes (see Conway et al., 2014) and to the best of our knowledge has not been reported for the Nemacheilidae . The examination of additional specimens of Ph. mango will be needed to confirm if the variation reported herein represents sexual dimorphism or individual variation.
The largest individual of Physoschistura mango that we have examined is a nuptial male, 23.4 mm SL. Using the criteria of Weitzman & Vari (1988), Ph. mango represents a miniature species and would be qualified to be added to the list of miniature freshwater fishes from South and Southeast Asia compiled by Kottelat & Vidthayanon (1993) over 30 years ago, and certainly in need of an update considering The novel, and potentially sexually dimorphic, barbel tubercles of male Physoschistura mango , a characteristic otherwise unknown from the 800 or so described species of nemacheilid loaches, should also be considered within the context of miniaturisation (i.e., morphological novelty; Hanken & Wake, 1993: 506). Morphological novelties are common in miniature Cypriniformes and include for example, the modified pectoral and pelvic fins, and pelvic pad in male Paedocypris ( Kottelat et al., 2006; Britz and Conway, 2009); modified pectoral fin, drumming muscle and sexually dimorphic Weberian apparatus and fifth rib of male Sundadanio ( Conway & Britz, 2007; Conway et al., 2011); the anteroventral tip of left cleithrum projecting into a strong spine in Fangfangia ( Britz et al., 2012) ; the anterior anus and urogenital opening, enlarged paired fins, drumming muscle and sexually dimorphic Weberian apparatus and fifth rib of male Danionella ( Britz, 2003, 2009; Britz & Conway, 2016; Britz et al., 2009, 2021); and the strange humeral organ of male Barboides ( Loiselle & Welcomme, 1971; Conway & Moritz, 2006; Conway et al., 2017). The vast majority of the aforementioned morphological novelties are sexually dimorphic and likely serve different roles in reproduction, as confirmed recently for Paedocypris ( Britz & Kottelat, 2008) and Danionella ( Schulze et al., 2018; Tatarsky et al., 2022). Given the popularity of Ph. mango in the aquarium hobby, we expect more detailed information on courtship and reproduction under aquarium conditions is likely to become available in the future.
Comparative material: See Kottelat (1990, 2018) for list of material of Petruichthys and Physoschistura examined, plus the following material. Petruichthys brevis – Lake Inle; CAS 93216, 30 (of 35), 20.0–46.0 mm SL; NRM 28473, 2, 28.6–32.1 mm SL.



| TCWC |
Texas Cooperative Wildlife Collection |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.