
Guimaraesiella (Dicrurobates) Gustafsson & Bush, 2020
|
publication ID |
https://doi.org/10.11646/zootaxa.4885.2.1 |
|
publication LSID |
lsid:zoobank.org:pub:081203D8-39FF-41C3-A79A-BB63F47AB3B1 |
|
DOI |
https://doi.org/10.5281/zenodo.4332162 |
|
persistent identifier |
https://treatment.plazi.org/id/3487B0CA-0E09-4473-AB17-8DBB7D75DEA2 |
|
taxon LSID |
lsid:zoobank.org:act:3487B0CA-0E09-4473-AB17-8DBB7D75DEA2 |
|
treatment provided by |
Plazi |
|
scientific name |
Guimaraesiella (Dicrurobates) Gustafsson & Bush |
| status |
subgen. nov. |
Guimaraesiella (Dicrurobates) Gustafsson & Bush , new subgenus
urn:lsid:zoobank.org:act:3487B0CA-0E09-4473-AB17-8DBB7D75DEA2
Brueelia Kéler, 1936: 257 (in part).
Brueelia “clade C” Bush et al. 2016: 743, fig. 3c.
Type species: Brueelia dicruri Ansari, 1955 ex Dicrurus macrocercus albirictus (Hodgson, 1836) View in CoL .
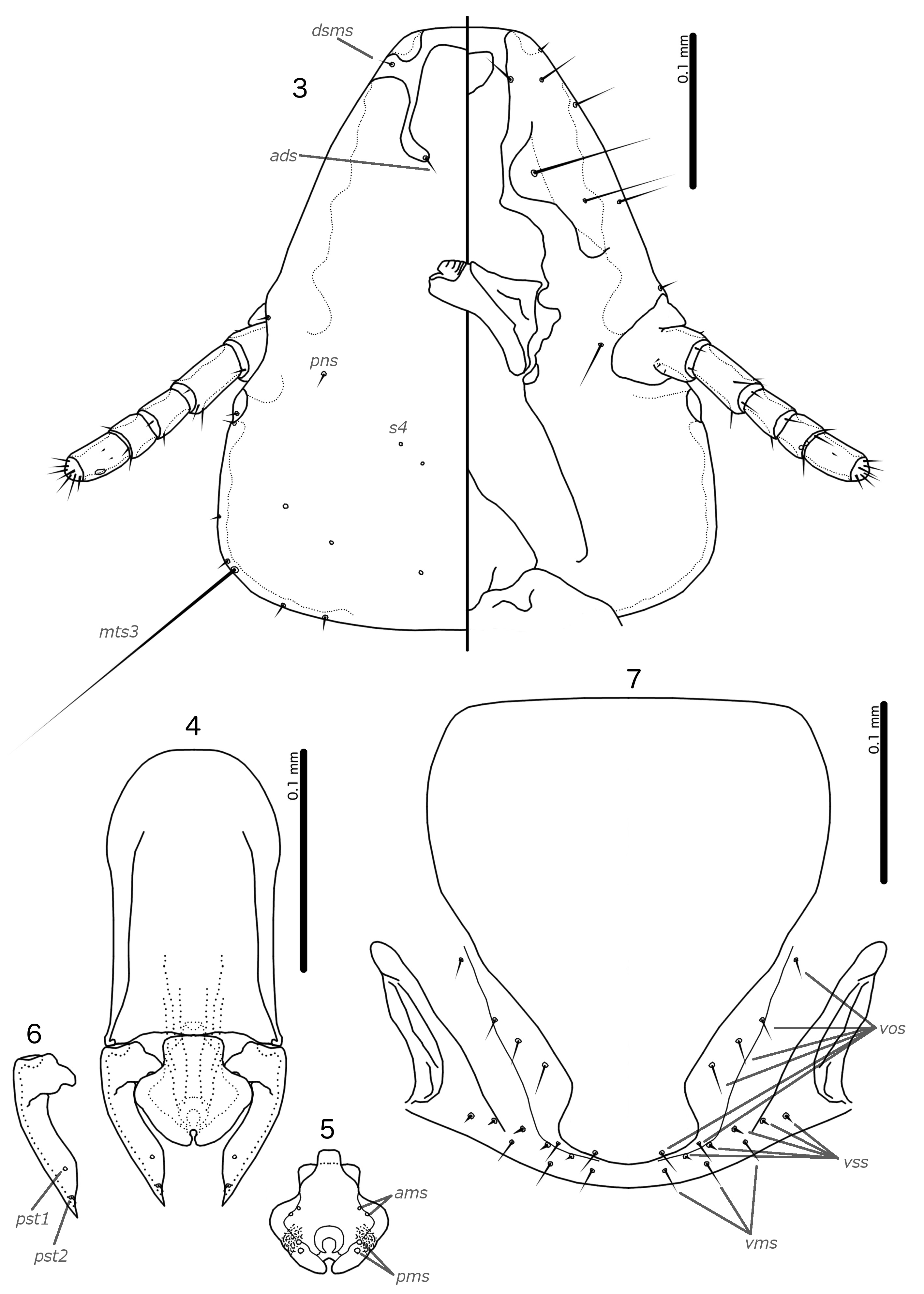
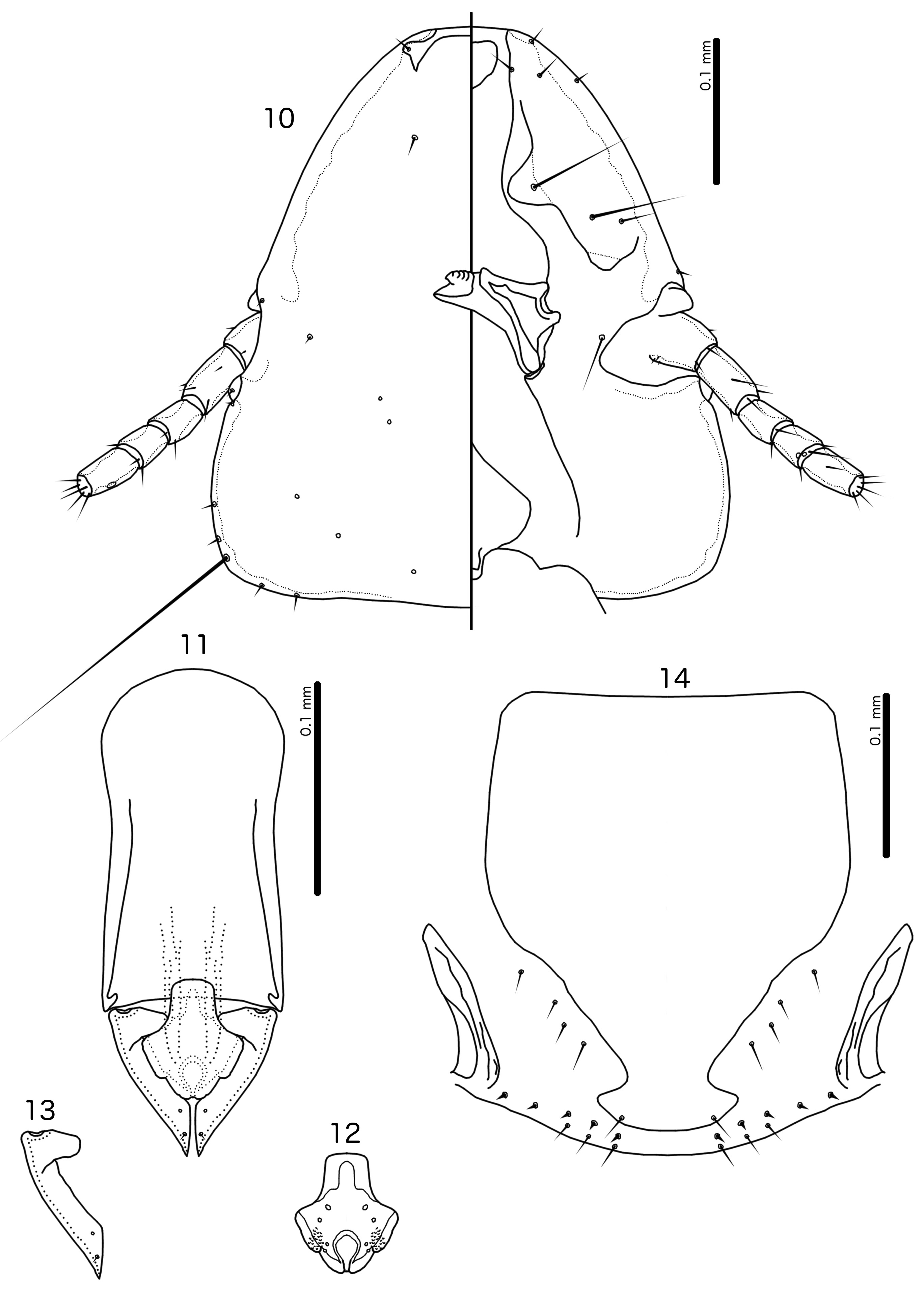
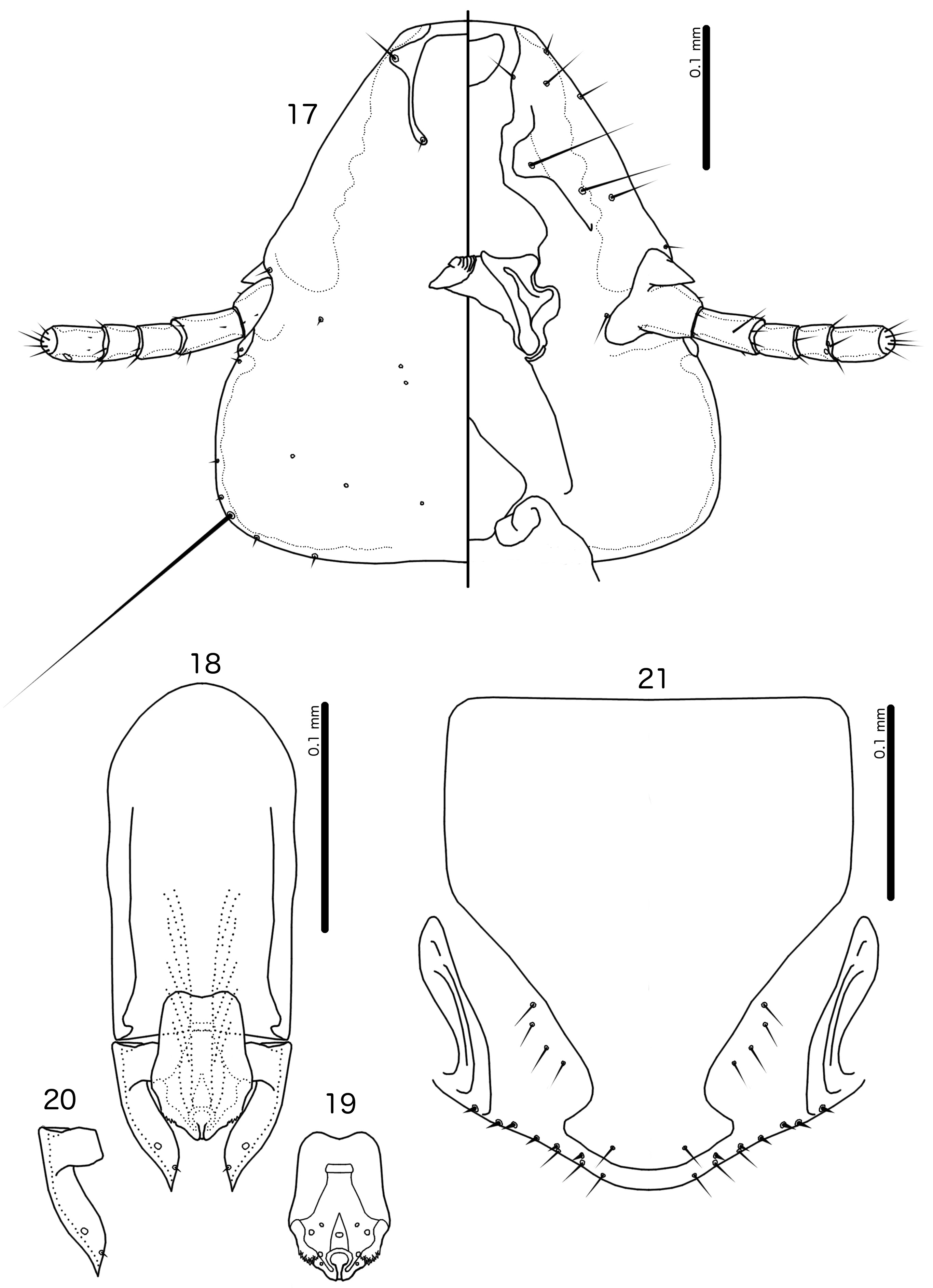
Diagnosis. The subgenus Guimaraesiella (Dicrurobates) shares the following characters with the nominate subgenus: dorsal preantennal suture present; marginal carina interrupted at least medianly; pns and s4 present; as3 absent; psps present on tergopleurites IV–VII; setal rows absent on all tergopleurites in both sexes; ss present on tergopleurites II–VIII in both sexes; aps present on male tergopleurite VII; parameral heads folded medianly; gonopore open distally. However, species included in Guimaraesiella (Dicrurobates) can be separated from those in the nominate subgenus by two characters of the male genitalia: (1) gonopore positioned terminally in species of Guimaraesiella (Guimaraesiella) , but subterminally in species of Guimaraesiella (Dicrurobates) (e.g. Figs 5 View FIGURES 3–7 , 12 View FIGURES 10–14 , 19 View FIGURES 17–21 ); (2) rugose nodi present in species of Guimaraesiella (Dicrurobates) (e.g. Figs 5 View FIGURES 3–7 , 12 View FIGURES 10–14 , 19 View FIGURES 17–21 ), but absent in species of Guimaraesiella (Guimaraesiella) . Both these characters are found in at least some species included in the subgenera Guimaraesiella (Mohoaticus) Mey, 2017 and Guimaraesiella (Cicchinella) Gustafsson et al., 2019a (gombakensis and tenella species groups). Hence, Guimaraesiella (Dicrurobates) may be closer to one or both of these subgenera than to Guimaraesiella sensu stricto. Notably, in the phylogeny of Bush et al. (2016: 742, fig. 3(b), clade A-5), the subgenus Guimaraesiella (Cicchinella) was not placed close to the nominate subgenus; however, no member of the subgenus Guimaraesiella (Mohoaticus) was represented in that phylogeny, and relationships among the deeper nodes within Guimaraesiella sensu lato were poorly resolved.
Species of Guimaraesiella (Dicrurobates) can be separated from species of Guimaraesiella (Cicchinella) with rugose nodi by the following characters: (1) female subgenital plate with cross-piece in Guimaraesiella (Cicchinella) (gombakensis and tenella species groups), but without cross-piece in Guimaraesiella (Dicrurobates) (e.g. Figs 7 View FIGURES 3–7 , 14 View FIGURES 10–14 , 21 View FIGURES 17–21 ); (2) male gonopore ventral, near center of mesosome in Guimaraesiella (Cicchinella) (both species groups), but subterminal in Guimaraesiella (Dicrurobates) (e.g. Figs 5 View FIGURES 3–7 , 12 View FIGURES 10–14 , 19 View FIGURES 17–21 ); (3) ventral sclerite absent in Guimaraesiella (Cicchinella) (gombakensis species group), but present in Guimaraesiella (Dicrurobates) (e.g. Figs 5 View FIGURES 3–7 , 12 View FIGURES 10–14 , 19 View FIGURES 17–21 ); (4) parameral heads with corrugated section in Guimaraesiella (Cicchinella) (both species groups), but without such corrugation in Guimaraesiella (Dicrurobates) (e.g. Figs 6 View FIGURES 3–7 , 13 View FIGURES 10–14 , 20 View FIGURES 17–21 ); (5) dorsal preantennal suture completely separating dorsal anterior plate in Guimaraesiella (Cicchinella) (tenella species group), but not separating the dorsal anterior plate in Guimaraesiella (Dicrurobates) (e.g. Figs 3 View FIGURES 3–7 , 10 View FIGURES 10–14 , 17 View FIGURES 17–21 ).
Species of Guimaraesiella (Mohoaticus) can be separated from Guimaraesiella (Dicrurobates) by the following characters: (1) dorsal preantennal suture not medially continuous median to ads and dorsal anterior plate continuous with roof of head in Guimaraesiella (Dicrurobates) (e.g. Figs 3 View FIGURES 3–7 , 10 View FIGURES 10–14 , 17 View FIGURES 17–21 ), but suture medially continuous median to ads and dorsal anterior plate separated from roof of head in Guimaraesiella (Mohoaticus) ; (2) mesosomal lobes with straight or bulging lateral margins and rugose nodi either poorly delimited or on bulge in Guimaraesiella (Dicrurobates) (e.g. Figs 5 View FIGURES 3–7 , 12 View FIGURES 10–14 , 19 View FIGURES 17–21 ), but with deeply sinuous lateral margins and rugose nodi clearly delimited ventrally by a noticeable ridge in Guimaraesiella (Mohoaticus) ; (3) gonopore smooth anteriorly in Guimaraesiella (Dicrurobates) (e.g. Figs 5 View FIGURES 3–7 , 12 View FIGURES 10–14 , 19 View FIGURES 17–21 ), but serrated anteriorly in Guimaraesiella (Mohoaticus) .
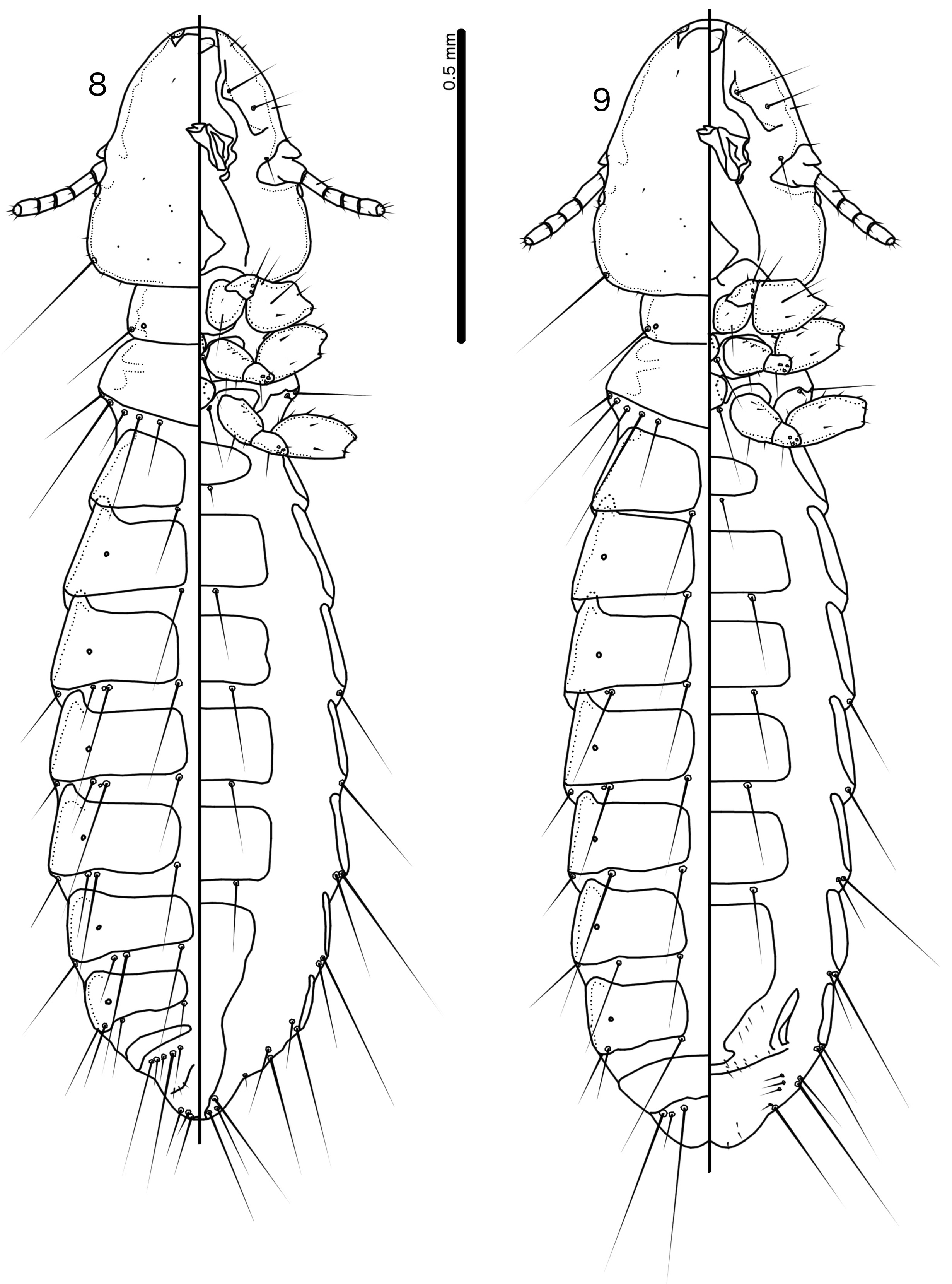
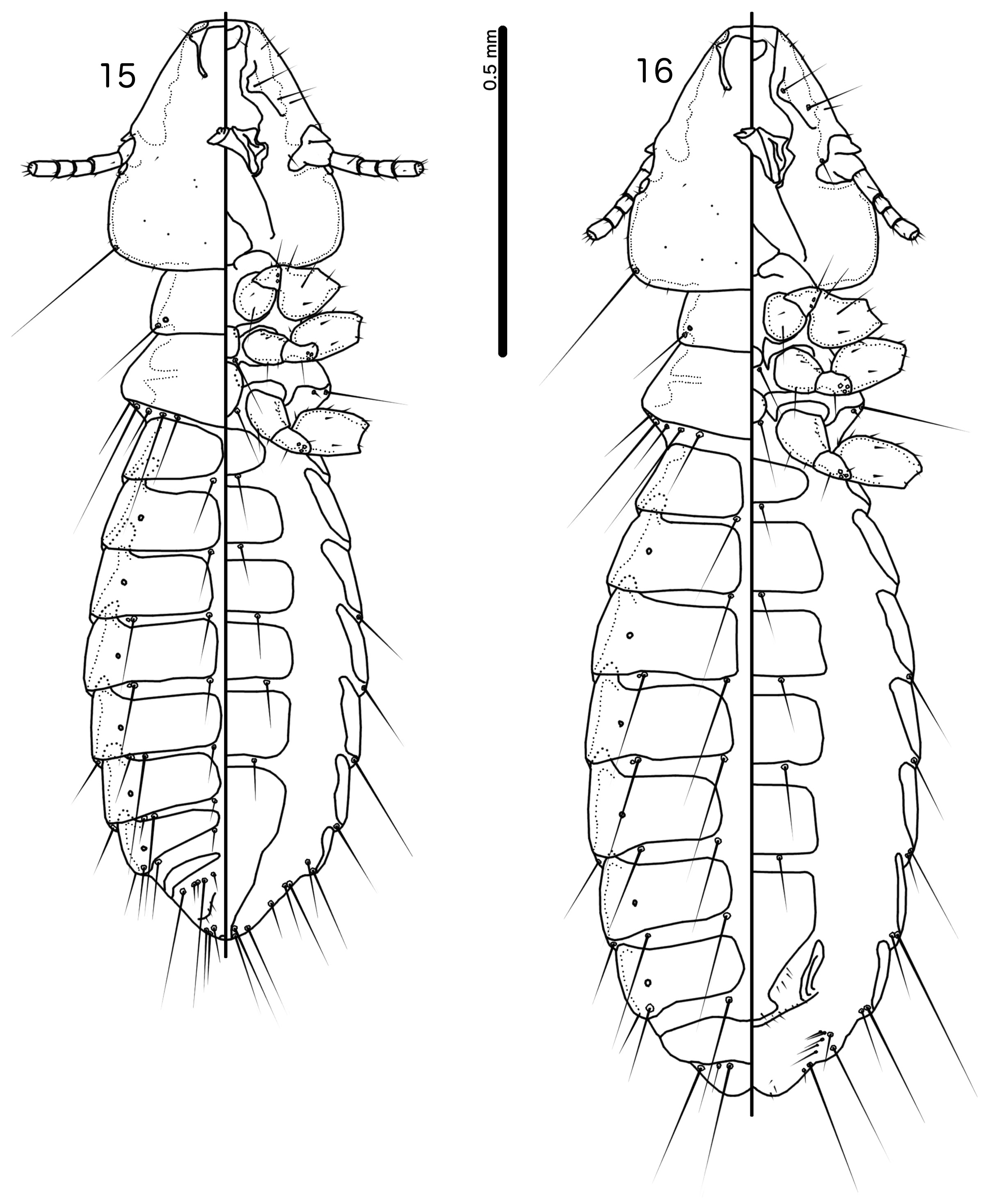
Description. Both sexes. Head shape variable, but preantennal area typically long and roughly trapezoidal, with flattened frons (e.g. Figs 3 View FIGURES 3–7 , 10 View FIGURES 10–14 , 17 View FIGURES 17–21 ). Marginal carina broad, with irregular inner margins, interrupted medianly but not laterally (except in species where dorsal preantennal suture reaches lateral margins of head); frons hyaline, continuous with dorsal preantennal suture which reaches at least dsms, and may reach ads and lateral margins of head; ventral anterior plate present; temporal and occipital carinae not visible; antennae sexually monomorphic; temples gently rounded. Head chaetotaxy as in e.g. Figs 3 View FIGURES 3–7 , 10 View FIGURES 10–14 , 17 View FIGURES 17–21 ; as3 absent; mts3 only temporal macrosetae.
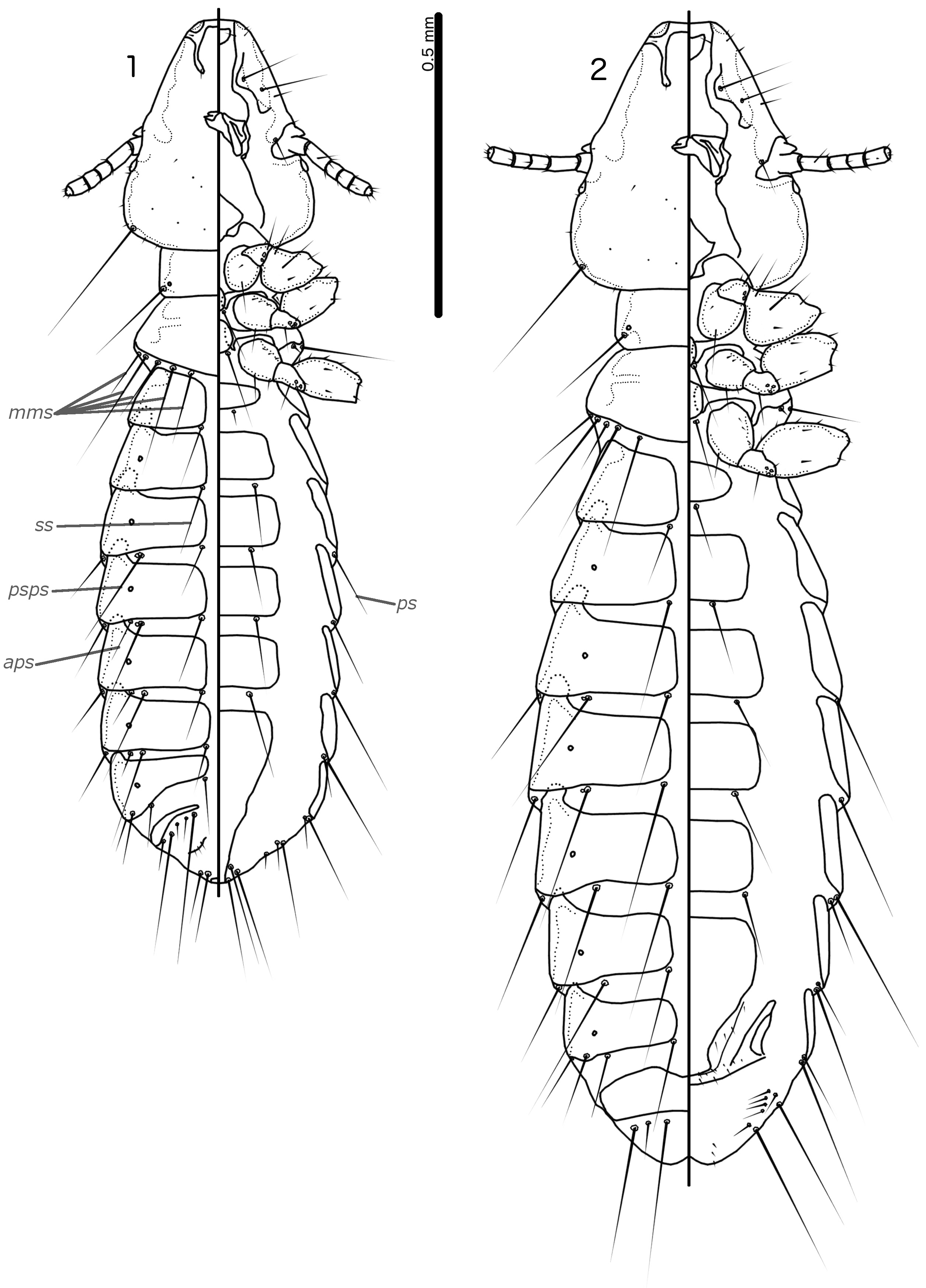
Prothorax rectangular; psps on postero-lateral corners. Pterothorax roughly pentagonal, with lateral margins divergent and posterior margin either rounded or convergent to median point; mms moderately separated medianly. Meso- and metasterna not fused, each with 1 seta on each side on postero-lateral corners (e.g. Figs 1–2 View FIGURES 1–2 , 8–9 View FIGURES 8–9 , 15–16 View FIGURES 15–16 ).
Male tergopleurites II–IX+X and female tergopleurites II–VIII divided medianly; ventral sections of tergopleurites generally slender. Sternal plates rectangular, not approaching ventral sections of tergopleurites; accessory sternal plates absent (e.g. Figs 1–2 View FIGURES 1–2 , 8–9 View FIGURES 8–9 , 15–16 View FIGURES 15–16 ).
Male. Abdominal chaetotaxy sparse, differing slightly between species. Subgenital plate roughly triangular, lateral margins typically irregular (e.g. Figs 1 View FIGURES 1–2 , 8 View FIGURES 8–9 , 15 View FIGURES 15–16 ). Genitalia: basal apodeme rectangular, with rounded anterior end, often constricted at mid-length (e.g. Figs 4 View FIGURES 3–7 , 11 View FIGURES 10–14 , 18 View FIGURES 17–21 ). Proximal mesosome variable, typically quadratic or rectangular, narrow compared to distal mesosome; ventral sclerite present, variable between species; mesosomal lobes gener-ally wider than proximal mesosome, with distinct bulging nodi lateral to gonopore; these nodi are typically at least partially rugose; 2 ames sensilla and 2 pmes sensilla on each side (e.g. Figs 5 View FIGURES 3–7 , 12 View FIGURES 10–14 , 19 View FIGURES 17–21 ). Parameral heads variable (e.g. Figs 6 View FIGURES 3–7 , 13 View FIGURES 10–14 , 20 View FIGURES 17–21 ). Parameral blades tapering only distally, may be slightly elongated distal to mesosome; pst1 sensillus located proximal to pst2; pst2 microseta, near distal end of paramere (e.g. Figs 4, 6 View FIGURES 3–7 , 11, 13 View FIGURES 10–14 , 18, 20 View FIGURES 17–21 ).
Female. Abdominal chaetotaxy sparse, differing slightly among species. Subgenital plate broad in anterior half, narrowing in posterior half; lateral margins of posterior half often irregular; subgenital plate does not reach vulval margin, but expands distally into lateral submarginal bulges; few vms and vss on each side; vos follow lateral margins of subgenital plate, with at least 1 distal vos separated from other vos by a gap, and distal most vos typically situated on or near distal margin of subgenital plate, near vss (e.g. Figs 7 View FIGURES 3–7 , 14 View FIGURES 10–14 , 21 View FIGURES 17–21 ).
Host distribution. Species of Dicruridae , Oriolidae and Vangidae .
Geographical range. Afrotropical, Indo-Malayan and Australo-Papuan regions; presumably present outside these regions wherever drongos occur.
Etymology. The name Dicrurobates is formed by the host genus Dicrurus Vieillot, 1816 —from Greek “ dikros ” = “forked”, and “ oura ” = “tail”—combined with “ bates ”, Greek for “one who walks on something”.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Guimaraesiella (Dicrurobates) Gustafsson & Bush
| Gustafsson, Daniel R. & Bush, Sarah E. 2020 |
Brueelia
| Bush, S. E. & Weckstein, J. D. & Gustafsson, D. R. & Allen, J. & DiBlasi, E. & Shreve, S. C. & Boldt, R. & Skeen, H. R. & Johnson, K. P. 2016: 743 |
Brueelia Kéler, 1936: 257
| Keler, S. von 1936: 257 |