
Trilobodrilus windansea, Kerbl & Vereide & Gonzalez & Rouse & Worsaae, 2018
|
publication ID |
https://doi.org/ 10.5852/ejt.2018.421 |
|
publication LSID |
lsid:zoobank.org:pub:BC3D3D2B-7EBB-4A5C-8FB8-25D33EA4E524 |
|
DOI |
https://doi.org/10.5281/zenodo.5690082 |
|
persistent identifier |
https://treatment.plazi.org/id/03FFD60D-FFC1-FF92-FD9E-8679FB2FFBDC |
|
treatment provided by |
Plazi |
|
scientific name |
Trilobodrilus windansea |
| status |
sp. nov. |
Trilobodrilus windansea View in CoL sp. nov.
urn:lsid:zoobank.org:act:A9DB66C3-C548-45C1-98F6-6EFBA019CF40
Fig. 1 View Fig. 1 , Tables 1, 3–7
Diagnosis
Trilobodrilus with two pairs of long prostomial compound cilia (1.5–2 times longer than locomotory cilia), encircled by five pairs of intermediate ciliary tufts anterior to the prostomial ciliary bands. First ciliary band with small dorsal gap. Dorsally incomplete second ciliary band with mid-dorsal ciliary tuft and additional posterior ciliary row. Dorsally incomplete third ciliary band posterior to nuchal organs, elongated epidermal inclusions, no spindle glands.
Etymology
This species is named after the beach where it was collected, Windansea Beach.
Material examined
Holotype
UNITED STATES OF AMERICA: complete adult, 805 µm long (platinum-palladinium-coated and mounted on stub for SEM), Windansea Beach, La Jolla, San Diego , California, 32°49′46″ N, 117°16′51″ W, coarse sand in holes at 0.5 m and 1 m at the mid ‐ and high ‐water mark, respectively, 14 Apr. 2009, K Worsaae and G Rouse leg. ( SIO-BIC A8206 ).
GoogleMapsParatypes
UNITED STATES OF AMERICA: 4 specimens (2 mounted on same stub as holotype for SEM, 2 in 70% EtOH), same locality and sampling site as the holotype, 14 and 17 April 2009 (SIO-BIC A8207, SIO-BIC A8208, NHMD- 210468). Additional specimens mounted on slides were unfortunately lost after conducting the measurements.
Description
Measurements given from holotype, ranges given in parenthesis include paratypes and lost specimens.
Transparent body, light brown to dark brown tint (live and fixed specimens) ( Fig. 1A View Fig. 1 ). Body length 715 µm (499–1040 µm, n = 12), width 86 µm (84–189 µm, n = 12), body segments indistinct ( Fig. 1A– C View Fig. 1 , Tables 1, 7 View Table 7 ).
Prostomial shape square ( Fig. 1A–F View Fig. 1 , Table 7 View Table 7 ). Eyes lacking. Middle of mouth located 110 µm (77– 133 µm, n = 9) posterior of the apical tip (mo, Fig. 1A, C–D View Fig. 1 , Table 1). Four compound cilia terminally on prostomium (pcc, Fig. 1B, E–F View Fig. 1 ), each consisting of approximately 50–60 cilia (n = 3), spaced 32 µm apart (18–32 µm, n = 10). Compound cilia substantially (up to two times) longer than locomotory cilia in ciliary bands (19–43 µm relative to 8–20 µm in ciliary bands, n = 10, Fig. 1A, D View Fig. 1 , Table 1). Prostomial compound cilia surrounded by five pairs of semicircularly arranged apical ciliary tufts (act, Fig. 1B, E–F View Fig. 1 , Tables 1, 7 View Table 7 ). Two ciliary bands on prostomium and one ciliary band posterior to the nuchal organs (cb1–3, Fig. 1A–F View Fig. 1 , Table 7 View Table 7 ). First ciliary band (12–19 µm wide, n = 3) encircles prostomium with 7–8 µm wide dorsal gap (cb1, Fig. 1A–B, E–F View Fig. 1 , Table 7 View Table 7 ). One pair of intermediate ciliary tufts located laterally between first and second ciliary band (ict, Fig. 1B, E View Fig. 1 ). Second ciliary band (13–19 µm wide) dorsally incomplete with 43–46 µm wide gap and one mid-dorsal ciliary tuft in center of gap (mdt, Fig. 1B, E–F View Fig. 1 , Table 7 View Table 7 ; consisting of 50–70 cilia, n = 3). One additional thin row of cilia (5–7 µm wide, n = 2) lines the second ciliary band postero-laterally, not extending dorsally for the entire length of the second ciliary band (acr, Fig. 1E–F View Fig. 1 , Table 7 View Table 7 ).
One lateral pair of nuchal organs located between the second and third ciliary band (no, Fig. 1B, E–F View Fig. 1 ). Third ciliary band dorsally incomplete with wide gap (cb3, Fig. 1B, E–F View Fig. 1 , Table 7 View Table 7 ); width of third ciliary band 10–17 µm, width of gap 39–50 µm (n = 3). Individual cilia scattered across the body ( Fig. 1B, E View Fig. 1 ). The ventral ciliary tract extends from posterior prostomium to posterior pygidium (vct, Fig. 1B–C, E View Fig. 1 ; width of tract relative to total body width approximately 0.6 (n = 3)). Anus opening dorso-anteriorly on pygidium.
Eggs 126–143 µm long, located 534–464 µm posterior to the prostomial tip and 209–194 µm (n = 2) anterior to pygidial tip ( Fig. 1G View Fig. 1 ).
Epidermal inclusions in the epidermis ( Fig. 1H View Fig. 1 , Table 7 View Table 7 ); average measurement of envelope 4 × 9 µm, 4–8 spherules per envelope (n = 12); no spindle glands.
Molecular information
The following sequences were determined by standard sequencing from a single, non-type specimen collected on 14 Apr 2009, for which no morphological voucher remains: 18S rDNA, MG588089 View Materials (1857 nucleotides (nt), Table 4); 28S rDNA, MG588091 View Materials (1126 nt, Table 5); COI, MG588093 View Materials (644 nt, Table 6); CytB, MG588095 View Materials (421 nt). In the following, the sequences of T. windansea sp. nov. are first compared to the most similar sequences found in T. ellenscrippsae sp. nov., and subsequently the range of similarities to the respective species is listed.
Trilobodrilus windansea sp. nov. 18S rDNA fragment is 99.9% similar to the 18S rDNA of T. ellenscrippsae sp. nov., and 99.1% ( T. nipponicus ) – 99.5% ( T. axi ) similar to the 18S rDNA fragments of the other sequenced species ( Table 4). Its 28S rDNA fragment is 99.9% similar to the 28S rDNA of T. ellenscrippsae sp. nov., and 98.6% ( T. axi ) – 99.4% ( T. nipponicus ) similar to the 28S rDNA fragments of the other sequenced species ( Table 5). COI is 84.9% similar to the COI of T. ellenscrippsae sp. nov., and 76.9% ( T. itoi ) – 78.6% ( T. nipponicus ) similar to the sequences of the other species ( Table 6). Cytochrome B is 85.6% similar to T. ellenscrippsae sp. nov.
In both the maximum-likelihood as well as the Bayesian analyses, T. windansea sp. nov. was found to be the sister species of T. ellenscrippsae sp. nov. with full support, the two of them forming the sister group to the T. itoi – T. nipponicus clade ( Fig. 2 View Fig. 2 ). These Pacific species were shown to be the sister clade to the group formed by the Atlantic species T. axi and T. heideri ( Fig. 2 View Fig. 2 ). All Trilobodrilus -species form a sister-clade to Dinophilus sp., a representative of the second dinophilid genus ( Fig. 2 View Fig. 2 ).
Habitat
Intertidal zone of a clean, coarse, well-sorted sandy beach, 0.5–1 m below the mid- and high water mark.
Distribution
Trilobodrilus windansea sp. nov. is only known from Windansea Beach, La Jolla, San Diego, California.
Remarks
Trilobodrilus windansea sp. nov. most closely resembles T. itoi in morphology (but T. ellenscrippsae sp. nov. in its molecular profile), but still differs by having more apical ciliary tufts, by having an additional row of cilia posterior to the second ciliary band, and by lacking a fourth ciliary band as well as segmentally arranged ciliary tufts along the body ( Fig. 1 View Fig. 1 , Table 7 View Table 7 ). Furthermore, the epidermal inclusions are more elongated and include fewer spherules than in T. itoi ( Table 7 View Table 7 ). Trilobodrilus windansea sp. nov. resembles the other Californian species, T. ellenscrippsae sp. nov., in having a dorsally incomplete second ciliary band with mid ‐dorsal tuft, but differs in having longer prostomial compound cilia, one more pair of apical ciliary tufts, a small dorsal gap in the first ciliary band, an additional row of cilia posterior to the second ciliary band, a dorsally incomplete third ciliary band, and by lacking a fourth ciliary band ( Fig. 1 View Fig. 1 , Table 7 View Table 7 ).
Trilobodrilus windansea sp. nov. further differs from T. ellenscrippsae sp. nov., T. itoi , T. axi and T. heideri in comparison of gene fragments, see Molecular information above or Tables 4–6.
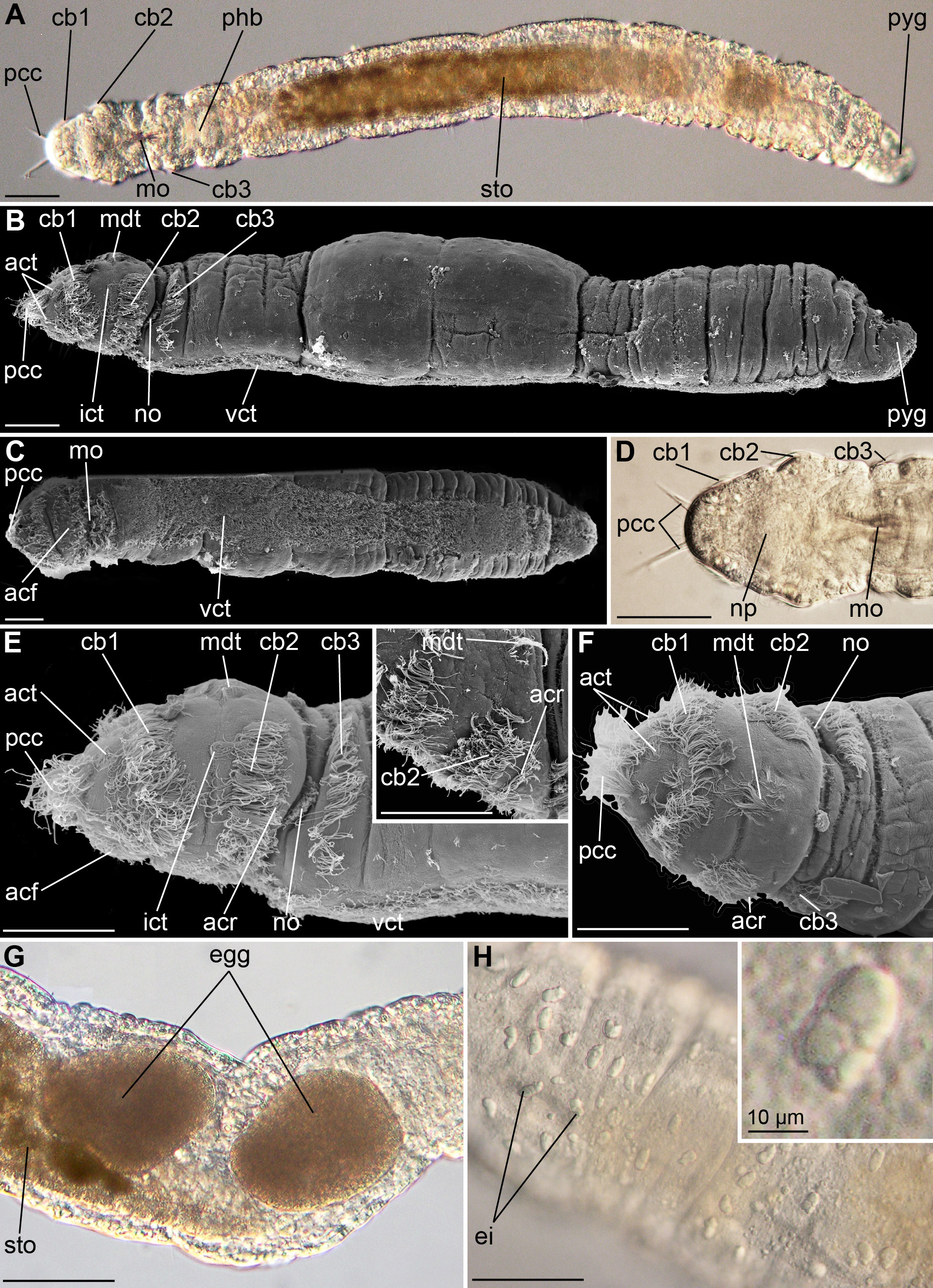
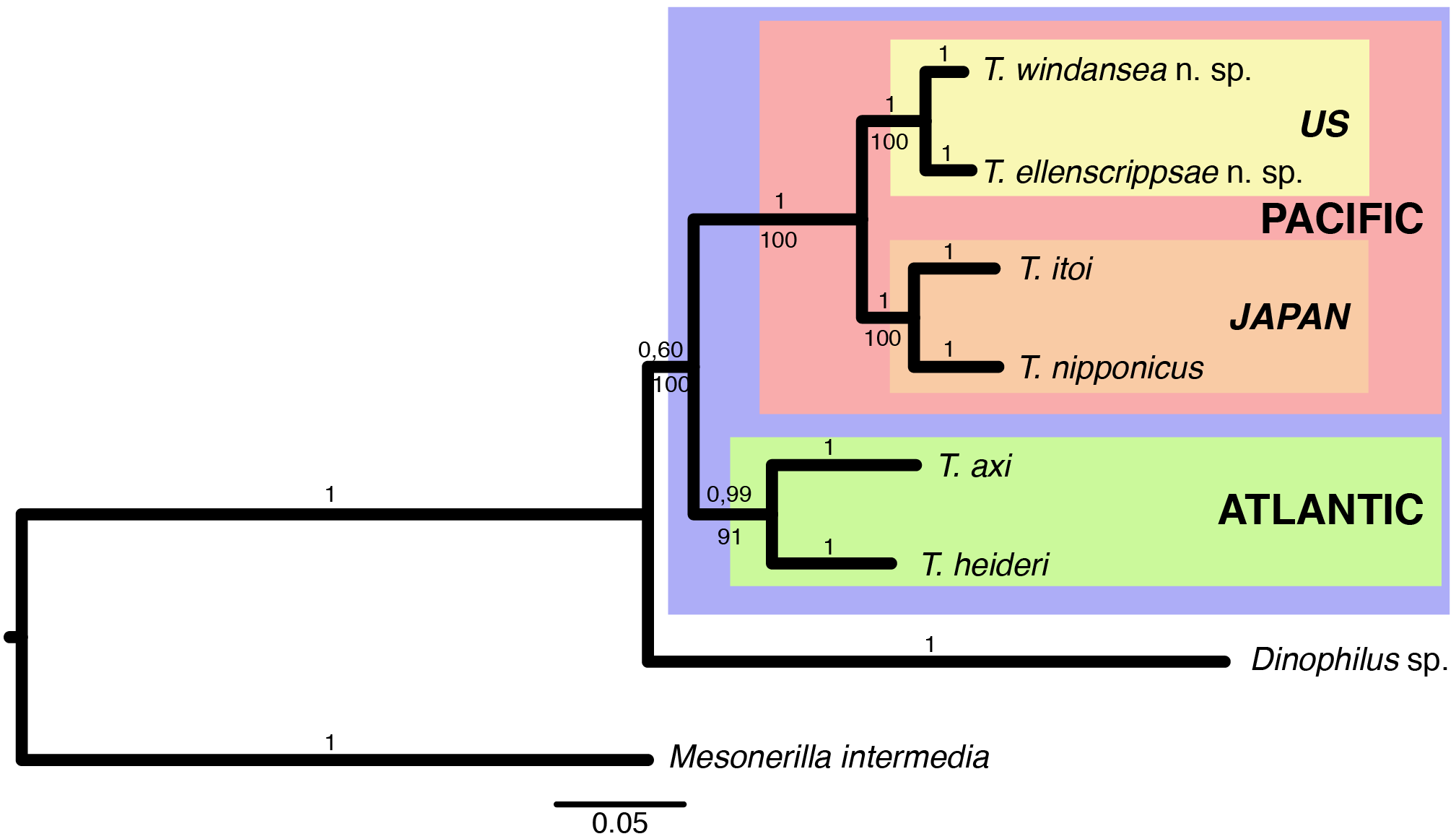
Table 7 (next page). Morphological characters and their states in all described species of Trilobodrilus Remane, 1925. Measurements are given in mm for body length and width, and in µm for the size of epidermal inclusions; the latterare takenfromKajiharaet al. (2015),andfrom theoriginal sources(Rao 1973; Remane 1925; Riser 1999; Uchida & Okuda 1943; Westheide 1967, 2008). Abbreviations: G = gonochoristic; H = hermaphroditic.
| Species | T. windansea | T. ellenscrippsae | T. heideri | T. axi | T. hermaphroditus | T. indicus | T. nipponicus | T. itoi |
|---|---|---|---|---|---|---|---|---|
| sp. nov. | sp. nov. | Remane, 1925 | Westheide, Riser, 1999 1967 | Rao, 1973 | Uchida & Okuda, 1943 | Kajihara, Ikoma, Yamasaki & | ||
| Hiruta, 2015 | ||||||||
| Body length [mm] | 0.5–1.0 | 0.5–0.7 | 1.5–1.9 | 1 | 1.7 | 1.0–1.2 | 0.7–1.4 | 0.9–1.3 |
| Body width [mm] | 0.08–0.2 | 0.09–0.2 | 0.1–0.2 | 0.1 | 0.22 | 0.1–0.12 | 0.09–0.16 | 0.08–0.12 |
| Prostomial shape | Squared | Conical | Squared | Conical | Conical | Conical | Conical | Squared |
| Pairs of apical ciliary tufts | 5 | 4 | ? | ? | ? | ? | 2–3 | 3–4 |
| Mid-dorsal tufts on 2nd ciliary | 1 | 1 | 0 | 0 | 0 (but 2 stereocilia) | 0 | 1 | 1 |
| band (#) | ||||||||
| Interm. ciliary tufts (#) | present (1) | present (>2) | ? (absent) | ? (present) | ? | ? | present (1) | present (>2) |
| Add. ciliary row(s) posterior of 2nd ciliary band | present | absent | present | present | absent | ? | absent | absent |
| 3rd ciliary band dorsally | absent | present | absent | absent | present (but not | present | present | absent |
| continuous | meeting ventral | |||||||
| ciliary tract) | ||||||||
| 4th ciliary band | absent | present | absent | present | ? | ? | present | present |
| Segmentally arranged lateral | absent | present (not as | ? | ? | present | ? (absent) | present | present |
| ciliary tufts | regular as in other | |||||||
| species) | ||||||||
| Epidermal inclusions [µm × µm] 4 × 9 | ? | ? | 10 × 15 | 6 × 9 | 3 × 12 | 5.5 × 16 | 7 × 9 | |
| Spherules/ envelope (#) | 4–8 | ? | 3–10 | 5–19 | 3–7 | 7–11 | 9–13 | 3–22 |
| Length/ width of epidermal inclusions (mean) | 2.2 | ? | 1.41 | 1.81 | 1.42 | 2.42 | 2.91 | 1.29 |
| Spindle glands | absent | ? | present | present | present | present | absent | absent |
| Sex | G | G | G | G | H | G | G | G |
| Habitat | intertidal | subtidal | subtidal | intertidal | intertidal/ subtidal | intertidal | intertidal | intertidal |
| Type locality | Windansea | La Jolla Cove, La | Helgo-land, | Sylt, | Nahant Bay, Palm Beach, Akkeshi Bay, | Ishikari Beach, | ||
| Beach, La Jolla, Jolla, California, | Germany | Germany | near middle of | Andhra | Japan | Japan | ||
| California, USA USA 9 m depth | shor of Nahant, | Pradesh, | ||||||
| Massachusetts, USA India | ||||||||
| 6 m depth | ||||||||
| Source | This study | This study | Remane (1925) | Westheide | Riser (1999) | Rao (1973) Uchida & Okuda Kajihara et al. | ||
| (1967) | (1943) | (2015) | ||||||
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |