
Bachia scaea, Jr, Mauro Teixeira, Vechio, Francisco Dal, Sales Nunes, Pedro M., Neto, Antonio Mollo, Lobo, Luciana Moreira, Storti, Luis Fernando, Gaiga, Renato Augusto Junqueira, Dias, Pedro Henrique Freire & Rodrigues, Miguel Trefaut, 2013
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3636.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:13E09793-A6C9-4CF2-9ED8-55A2A9F04541 |
|
DOI |
https://doi.org/10.5281/zenodo.5628249 |
|
persistent identifier |
https://treatment.plazi.org/id/03FF87F8-3819-FFBB-FF32-F93C904BFB75 |
|
treatment provided by |
Plazi |
|
scientific name |
Bachia scaea |
| status |
sp. nov. |
Bachia scaea View in CoL sp. nov.
( Figs. 1–2 View FIGURE 1 View FIGURE 2 )
Bachia flavescens Marçal et al. (2011: 262)
Holotype: MZUSP 101586, an adult male from the left bank of Madeira River, (9°26'55.30"S, 64°50'0.51"W, 101 m a.s.l., SAD69), Porto Velho municipality, state of Rondônia, Brazil, collected by the authors on 30th September 2010. Field number H 828.
Paratypes: MZUSP 103415, adult male and MZUSP 103414, juvenile; field numbers H 3133, H 3132, respectively (9°35'42.30"S, 65° 3'54.11"W, 96 m a.s.l., SAD69). MZUSP 100631, adult female; field number H 510 (9°26'16.18"S, 64°49'58.22"W, 123 m a.s.l., SAD69). MZUSP 103408, adult female; field number H 2479 (9°35'4.53"S, 65° 4'9.40"W, 129 m a.s.l., SAD69). MZUSP 101735, MZUSP 101736, MZUSP 103413, adult females; field numbers SAME 1130, SAME 1661, MTR 21266 (approximately 09°07' S, 64° 30'' W). All from Porto Velho municipality, state of Rondônia, Brazil.
Etymology: The specific epithet scaea is derived from the Greek word “ skaios ” which means “on the left”, referring to its geographical position regarding the Madeira River, as it is found only on its left bank, while its putative sister species, B. dorbignyi , is found only at its right bank.
Diagnosis: (1) A median-sized species of Bachia (maximum SVL= 69 mm); (2) prefrontals absent; (3) 5 supralabials; (3) supraoculars absent and 2-2 supraciliars; (4) femoral pores absent, 1-1 preanal pores in males; (5) 24–26 scales around midbody; (6) 51–54 dorsal rows of scales; (7) 40–43 ventral rows of scales; (8) 6–7 gulars; (9) 4 preanal shields; (10) no supralabials in contact with parietal; (11) dorsal scales hexagonal, imbricate, smooth; (12) fore and hindlimbs without clawed-digits; (13) first temporal present, separating fifth supralabial and parietal; (14) second chin shield not reaching oral border; (15) frontonasal present, reduced.
Description of the holotype: Body elongate, with a slight cervical constriction on head, snout rounded, tail longer than body. Rostral small, barely visible from above, contacting first supralabial, nasal and frontonasal. Viewed from above the rostral is about twice as wide as high; on lateral view it projects not far from the anterior margin of jaw. Frontonasal trapezoidal, as wide as long, wider posteriorly, contacting rostral, nasal and frontal. Prefrontals absent. Frontal roughly pentagonal, longer than wide, with anterior margin slightly convex, broader than, and in contact with frontonasal, and nasal; lateral margins straight to slightly concave, in contact with loreal, first and second superciliaries; posteriorly angulose, broadly contacting parietals. Frontoparietals and interparietal absent. Parietals large, longer than wide, slightly longer and slightly narrower than frontal, roughly pentagonal; their anterior margin deeply indented and in broad contact with frontal, contacting the second superciliary, three upper temporals and the dorsals. Posterior borders of parietals and dorsals coincides with a very slight transverse cervical constriction in the occipital region. Supraoculars absent. Two superciliaries of about the same size. Nasal large, longer than high. Nostril in the anterior lower margin of the nasal, invading the upper border of first supralabial. Loreal roughly squared, in contact with nasal, frontal, first superciliary, a small irregular anterior subocular and second and third supralabials. Frenocular absent. Six supralabials; third, fourth and fifth under the orbital region, fifth the highest and largest, not contacting parietal; fourth the smallest. Three suboculars; second longest; third one contacting second superciliary and an elongate anterior temporal. Postocular absent. Eyelid present with an undivided semitransparent disc. A small temporal scale between fifth and sixth supralabials and first and second temporal scales. Second upper temporal enlarged, longer than wide, diagonally disposed over a smaller scale contacting sixth supralabial. These two scales are followed by a similarly disposed third pair of temporal scales where the upper scale is the largest. Ear opening absent; its position marked by a longitudinal series of smaller granules. All head scales smooth and juxtaposed.
Mental roughly trapezoidal, wider than long, just broader than the ventral surface of rostral. Postmental roughly heptagonal; longer than wide. Two pairs of chin shields, both contacting infralabials; the anterior pair smaller, in broad contact at the midline; second pair in narrower contact at the midline, followed by three pairs of symmetric flat elongate pregulars, inner ones the largest. Five infralabials; first, second and third with about the same size. Gulars smooth, imbricate, rounded posteriorly, in seven transversal rows; scales of gular rows increasing gradually in size toward interbrachial region. Interbrachial region with two central, longer than wide central scales, emarginated at each side by a pair of two smaller scales, the lower one larger. Lateral scales of neck subrectangular, smooth, imbricate, slightly rounded posteriorly and longer than wide, disposed in regular transverse rows and becoming gradually similar to adjacent dorsal or ventral scales. Collar fold absent.
Dorsal scales imbricate and disposed in regular transversal rows; smooth, subrectangular and wider in occipital region, becoming progressively narrower, more elongate and rounded towards the level of the forelimbs and then on longer, hexagonal, lanceolate, smooth, with lateral sides almost juxtaposed. Fifty-three transverse rows between interparietal and the level of hind limbs. Lateral scales about the same size as the dorsals but smooth and less acuminate; those closer to ventrals slightly wider. A distinctive area with granular scales surrounds the area of forelimbs insertion and the posterior part of hindlimbs insertion. Twenty five scales around the midbody. Ventral scales smooth, longitudinally imbricate, laterally juxtaposed, almost squared just after the interbrachial row, becoming gradually longer than wide, rounded posteriorly, those after midbody narrower; forty transverse rows between interbrachials and preanals. Four preanal shields, divided in three longitudinal rows, central one with two scales aligned longitudinally; one small scale on each side of anal plate. One preanal pore, femoral pores absent.
Scales of tail similar to midbody dorsals, smooth, lanceolate, strongly imbricated, with 121 transverse rows.
Fore limbs stiliform, covered by smooth and imbricate scales, ending by three apical scales with no claws. Hind limbs also rudimentary, very reduced in size.
Background color of dorsal and lateral surfaces of body and tail dark brown with a pair of dorsolateral yellowish stripes extending from the lateral edge of parietals to the tip of the tail, and two paravertebral faded cream lines beginning at the fifth dorsal row, coalescing just after the hindlimb level and fading. Ventral parts of body and tail brown.
Measurements of the holotype (mm): SVL= 62.2 mm; TL= 107.4 mm
Variation: the sample is fairly homogeneous in scale counts and dorsal color pattern, however some variation in head scalation can be observed; in three specimens (MZUSP 103408, 103414–15,) the first temporal is followed directly by the second temporal, whereas in the five other specimens (MZUSP 100631, 101586, 101735 –36, 103413), there is a small scale between the first and second temporals and the supralabials. Ontogenetic variation on dorsal pattern can also be observed, with the two paravertebral cream lines merging at the cloacal level and entering the tail, fading posteriorly in juveniles; in adults after merging at the cloacal level the line disappears, rendering the dorsal surface of tail dark brown.
Comparison with other species (data from species in comparison are given in parentheses): the new species is a member of the Bachia dorbignyi group by having hexagonal, smooth, imbricate dorsal scales; quadrangular, juxtaposed ventral scales; supraoculars absent; interparietal and prefrontal absent; no femoral pores; 1-1 preanal pores; hindlimb stiliform and forelimbs ending in three fingers. These characters promptly distinguish B. scaea sp. nov. from all species of the B. bresslaui group (all with keeled dorsal scales, and supraoculars present), from the B. heteropa group (four digits on limbs and interparietal present) and from the B. flavescens group (quadrangular dorsal scales, supraoculars present).
Among the species of the Bachia dorbignyi group (diagnostic features for species of B. dorbignyi group are summarized in Table 1 View TABLE 1 ), B. scaea sp. nov. can be promptly distinguished from all species by the absence of clawed fingers ( Fig. 3 View FIGURE 3 ) (2–4 clawed fingers in combination for all other species). Additionally it differs from B. bicolor , B. huallagana , B. peruana , B. talpa and B. trisanale by having 6 supralabials (5) and from B. huallagana and B. trisanale by having 5 infralabials (4). By the absence of contact between supralabial and parietal it differs from B. dorbignyi (in the 5th), B. huallagana (3rd), B. peruana (4th), B. talpa (4th) and B. trisanale (4th). It also differs from B. dorbignyi , B. huallagana , B. peruana and B. trisanale by the presence of a first temporal (absent). By the absence of interparietal it differs from B. barbouri (present) and by the presence of a frontonasal it differs from B. trisanale (absent). The lower number of gulars, 6–7, distinguishes it from B. barbouri (8–10), B. bicolor , B. intermedia (both 7–9), B. talpa (9) and B. trisanale (7–8). By the lower number of scales around the body, 24–26, it can also be distinguished from B. barbouri (26–31), B. bicolor (27–31) and B. intermedia (28–35). By the number of dorsal scales, 51–54, it can also be distinguished from B. talpa (47–51). By the higher number of ventral scales, 42–45, it can be distinguished from B. barbouri (34–39), B. bicolor (34–40), B. intermedia (33–38) and B. talpa (36–38). Although information on the number of caudal scales is missing for most of the species, it can also be distinguished from B. dorbignyi by a higher number of rows, 111–121 (88–108). It can be further distinguished from B. bicolor , B. barbouri and B. talpa by the second chin shield not reaching the oral border (second chin shield reaches the oral border). It can also be distinguished from B. bicolor , B. talpa and B. trisanale by the presence of four preanal shields (three). Also differences in the dorsal color pattern distinguish it from all other Bachia species of the B. dorbignyi group (see Fig. 4 View FIGURE 4 for a summary of dorsal color patterns in the group, excluding B. talpa , for which we did not have data on its dorsal color).
Hemipenial morphology: The left hemipenis of MZUSP 103414 ( Fig. 5 View FIGURE 5 A) is unilobed with the distal tip of the retractor muscle divided. The organ is relatively small, extending along approximately four subcaudal rows (4 mm). The hemipenial body is cylindrical, slightly divided in the distal tip, indicating possibly vestigial lobes.
The sulcus spermaticus is a relatively broad channel, originating at the central region of the base of the organ, and proceeding in a straight line towards the apex. At the distal tip of the hemipenial body the sulcus is divided in two shallow branches. On the lobes, these branches run centrifugally ending at the external face of the tip. The full hemipenis is completely nude, with no evident plicae, papillae, ridges, calyces, mineralized spines or spinules in both faces (sulcate and asulcate), even after their immersion on Alizarin Red solution for 24 hours. The complete absence of evident hemipenial ornamentation on the genus Bachia is a common condition shared with other species, such as Bachia trisanale , B. intermedia (Presch 1978) , B. oxyrhina (Rodrigues et al. 2008) , B. heteropa alleni and B. bresslaui (Nunes 2011) .
The organ of B. dorbignyi ( Fig. 5 View FIGURE 5 B) is relatively similar with that of described above for B. scaea sp. nov., differing only in the shape of the hemipenial body, which is more globular, and in the width of sulcus spermaticus, wider than observed in the specimen of B. scaea sp. nov.
Distribution and Natural history: despite our large sampling effort in both banks of Madeira River, specimens of Bachia scaea sp. nov. were found only on its left bank ( Fig. 6 View FIGURE 6 ). The additional specimens from Porto Velho municipality were also found at the left bank. In the literature specimens referred to as B. gr. dorbignyi (Avila-Pires 2009) are recorded from a close locality, Guajará-Mirim, at the right bank of Madeira River. We have not examined these specimens, however the dorsal coloration matches that of B. dorbignyi , nonetheless it will be important to examine these specimens to attest if they belong to the new one or B. dorbignyi .
B. scaea B. B. B. B. B. B. B. B.
sp. nov. barbouri bicolor dorbignyi huallagana intermedia peruana talpa trisanale Caudals 111–121?? 88–108?????
SL 6 6 5 6 5 6 5 5 5
IL 5 5 5 5 4 5 5 5 4
SL-P none 5th none 5th 4th none 4th 4th 4th 1st temporal present absent present absent absent present absent absent absent Interparietal absent present absent absent absent absent absent absent absent Chin-oral no yes yes no no no no yes no Preanal shields 4 4 3 4 4 4 4 3 3
Hemipenis nude? flounced nude? nude?? nude Although no specimen was found active, one individual found during the day in a trail was supposed to be moving under the leaves. One individual was captured in a pitfall trap, while all others were found among the leaf litter and under rotten trunks and fallen logs, through active search. Although we managed to gather six specimens from the sampling areas, given the large effort, it seems to be rare.
The environment where B. scaea sp. nov. is found is dominated by a varzea forest (seasonally flooded forest by white-water rivers, such as the Madeira River), in some places with 35m tall trees, with dense leaf litter, crossed by small streams ( Fig. 7 View FIGURE 7 ), with the canopy frequently open due clearings. The altitude is about 90–100m a.s.l., however some higher grounds are also found, reaching up to about 300m a.s.l. Almost nothing is known about its biology, but a pregnant female (MZUSP 101735) carrying two eggs, observed through ventral skin, was found during the rainy season (December or January).
Phylogenetic relationships: The final standard deviation of split frequencies on the BA on the concatenate matrix had a very low value (0.002), and also for 16S (0.004) and c-mos (0.006) indicating stationarity. The concatenate dataset recovered Bachia scaea sp. nov. as sister to B. dorbignyi , but the support was not significantly high, in the 16S results they are still recovered as sisters but although higher the support are also not significant; for the c-mos results B. scaea sp. nov. is recovered in a polytomy together with B. intermedia , B. m. monodactylus , B. m. parkeri, B. bicolor , B. barbouri , B. peruana , B. trisanale and B. panoplia , but the support for this polytomy is also low. In fact, the overall support for most of the branches in all topologies are low, although some well supported structure is also observed, such as in the lineage that includes B. huallagana , B. scolecoides and B. heteropa alleni ( Fig. 8 View FIGURE 8 ).
Uncorrected p -distances ranged from 1 to 6% for c-mos, and 4 to 12% for 16S among Bachia species ( Table 2 View TABLE 2 ). Genetic distances between B. dorbignyi and B. scaea sp. nov. were 4% for 16S, and 2% for c-mos.
The description of Bachia scaea sp. nov. herein ends a long term stasis on the taxonomy of Amazonian Bachia species, especially in the B. dorbignyi group, which had its last species described by Dixon (1973) four decades ago.
Avila-Pires (1995) in her comprehensive work on the lizards from the Brazilian Amazonia commented that some Bachia species deserved a more careful examination. Regarding B. peruana , she argued that in a series of specimens from northern Acre state (Cruzeiro do Sul region), Brazil, although some features were coincident with the typical form, there were also other features that overlapped with characters of other species, such as the contact between fifth supralabial and parietal, similar to B. dorbignyi , the absence of frontonasal, similar to B. trisanale (Avila-Pires 1995) ; and also the absence of clawed fingers, which would distinguish it from other known species of the B. dorbignyi group. Nonetheless, as she examined only a few specimens from that region, she could not evaluate this issue. We have also examined two of three specimens she mentioned, and compared them to the examined B. peruana and Dixon’s (1973) data, who analysed nearlly 40 specimens of this species, and we are positive that this Cruzeiro do Sul population do not represent B. peruana . Indeed they are morphologically intermediate between B. dorbignyi and B. scaea sp. nov. However, as only two specimens (one sub-adult and one juvenile) are available, and variation between them is considerable, such as the presence of frontoparietal in one (ZUEC 436), while it is absent in the other (ZUEC 435) we prefer to wait for additional material in order to decide on their taxonomic status. At that time it should be also important to examine the previously referred specimens from this area (Avila-Pires 2009; Bernarde et al. 2011; SpeciesLink 2011), all referred to as Bachia sp. in our map ( Fig. 6 View FIGURE 6 ). A few individuals from southern Acre (Rio Branco region) were also made available to us. This sample fully matched the diagnostic features of B. trisanale ; a lower number of dorsal scales, frontonasal absent, presence of clawed-fingers, anal plate with three shields and the 4th supralabial touching parietal. Although the list of Brazilian reptiles (Bernils & Costa 2012) and IUCN Red List (Lehr & Doan 2010) indicate the presence of B. trisanale in Brazil, they do not have specific data on its occurrence (R. Bernils and E. Lehr per. comm.), thus to our knowledge this is the first unequivocal record of B. trisanale in Brazil.
The reported absence of clawed fingers in the northern Acre populations is also observed in all specimens of Bachia scaea sp. nov. In fact, clawed fingers are widespread among species of the B. dorbignyi group, thus its absence in B. scaea sp. nov. distinguishes it from all other species ( Fig. 3 View FIGURE 3 ), although its overall external morphology still resembles B. dorbignyi and B. peruana .
Nonetheless, the molecular data is not conclusive on Bachia scaea sp. nov. relationship, as the mitochondrial data suggests a close relationship with B. dorbignyi and the nuclear data place it in a polytomy with several species; thus only further analyses using additional genetic markers may elucidate this question. If confirmed, its relationship with B. dorbignyi will lead to an interesting biogeographical scenario, as both are geographically separated by the Madeira River. This river has already been recognized as a geographical barrier among closely related lineages within several groups (Cracraft 1985; Ayres & Cluttonbrock 1992; Haffer 1992; Avila-Pires 1995; Fernandes et al. 2012; Ribas et al. 2012; Tsuji-Nishikido et al. 2012), and could be associated with the break on the distributional range of the ancestor of B. scaea sp. nov. and B. dorbignyi , leading to their differentiation. Nonetheless a more comprehensive phylogenetic work is still needed to address this matter.
The upper Madeira River at the state of Rondônia crosses a lowland area that has been under a massive anthropic pressure. The varzea forests found at its right bank has been progressively anthropized, with the increasing of several destructive activities such as mining and pasturelands for cattle. Moreover, the installation of two hydroelectric dams, currently being built at the Madeira River, will probably transform drastically the landscape in the next few years. Fortunately, the left bank of Madeira River, where B. scaea sp. nov. is found, harbors more preserved forests, and also encompasses a National Park (Parque Nacional do Mapinguari) and higher lands, that will not be reached by the reservoirs, and hopefully some populations will not be affected.
The results presented here indicates that the diversity of Bachia at the Brazilian Amazonia is far from being comprehensively known, and it is likely to change in the future through further taxonomic studies on widespread species, and more surveys on poorly sampled areas, which will certainly find that its richness is even higher than currently known.
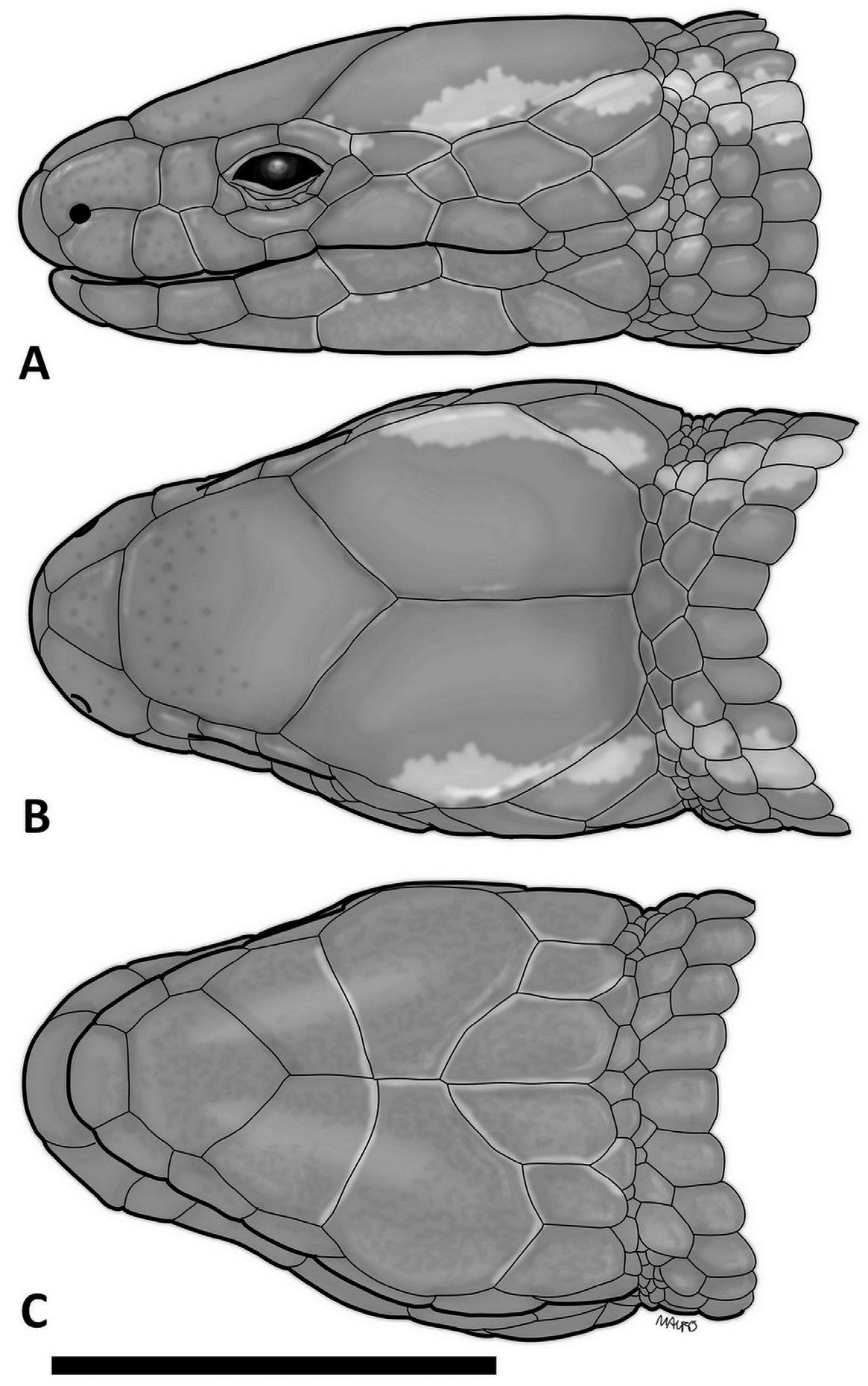

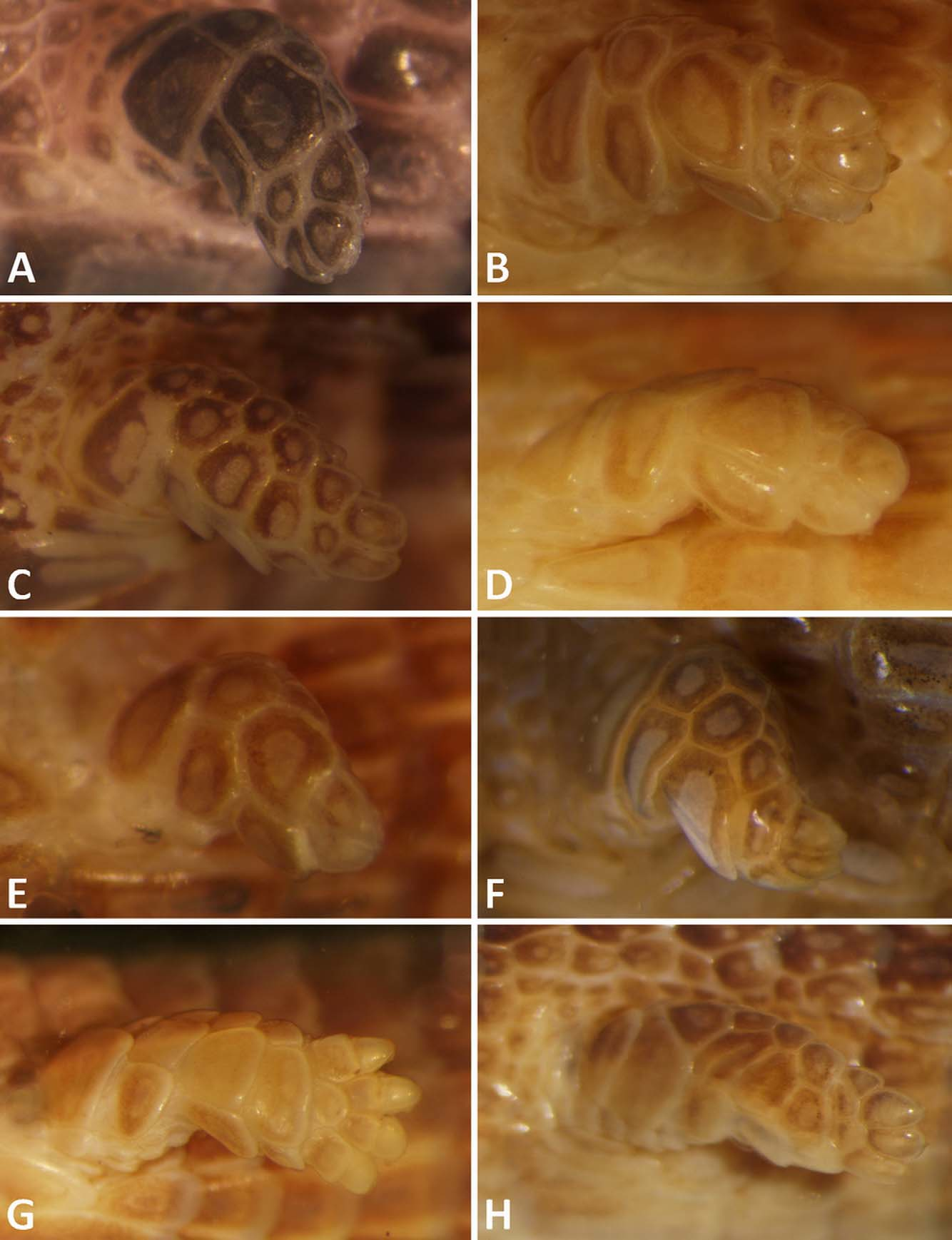
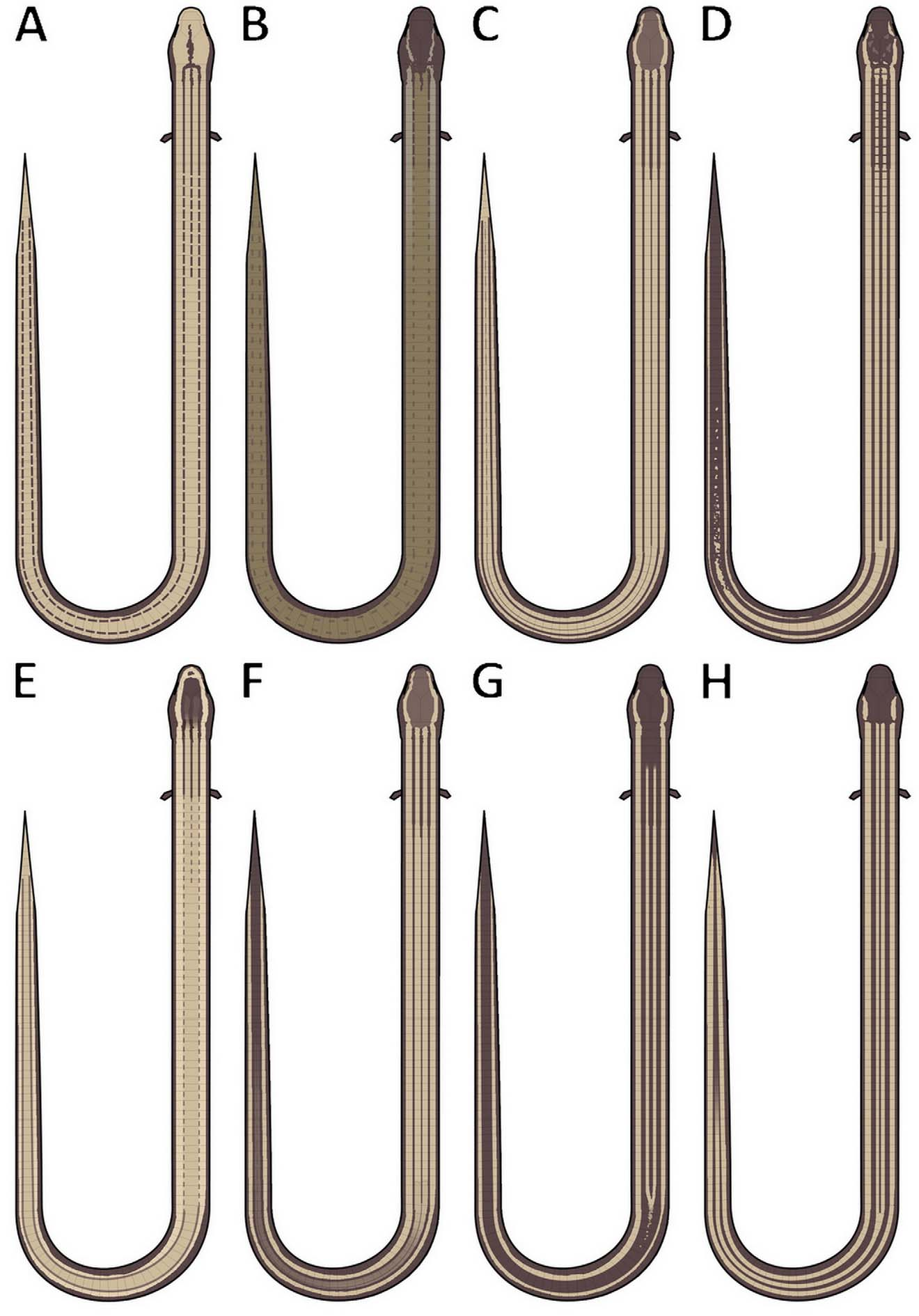
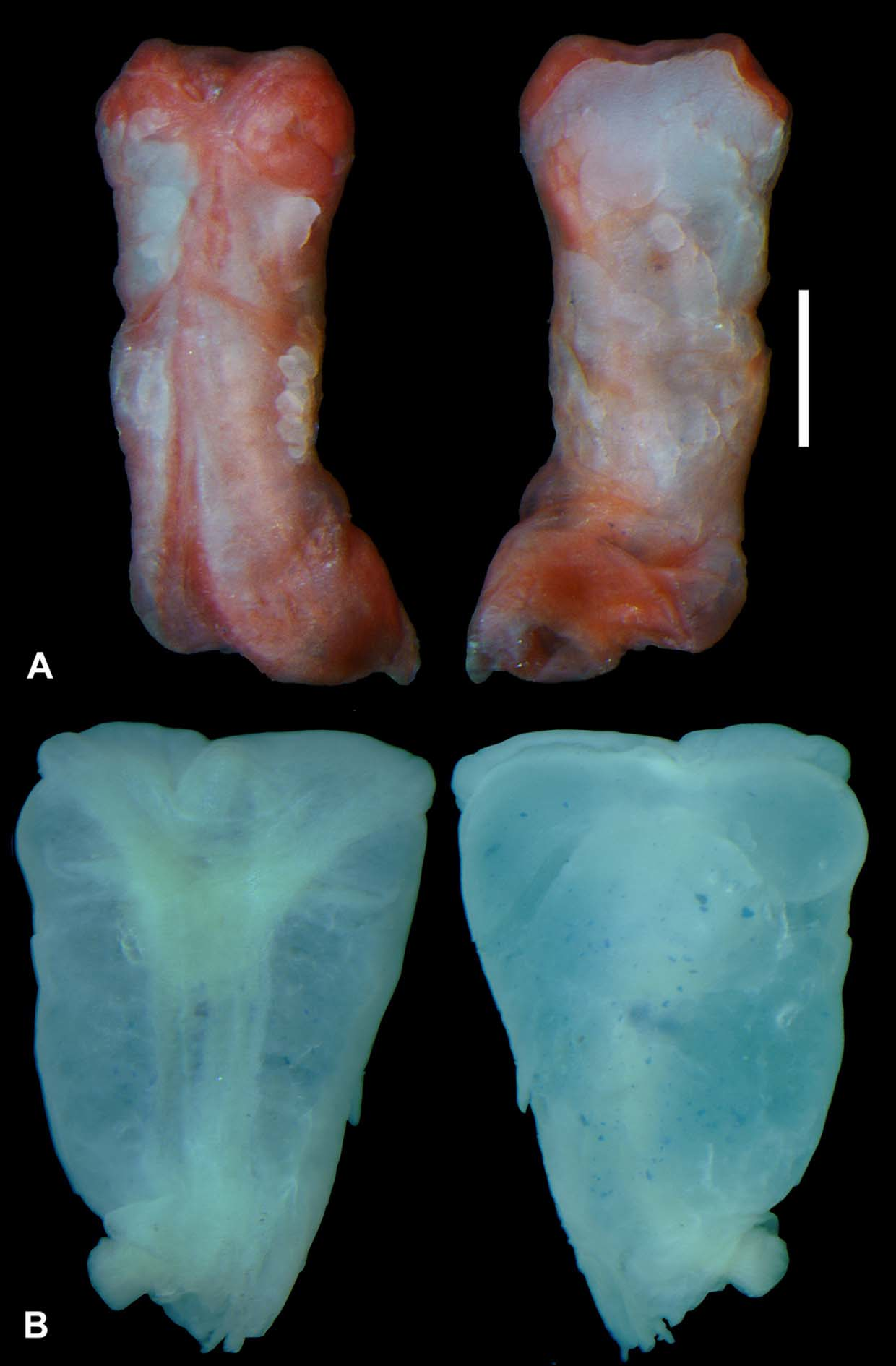
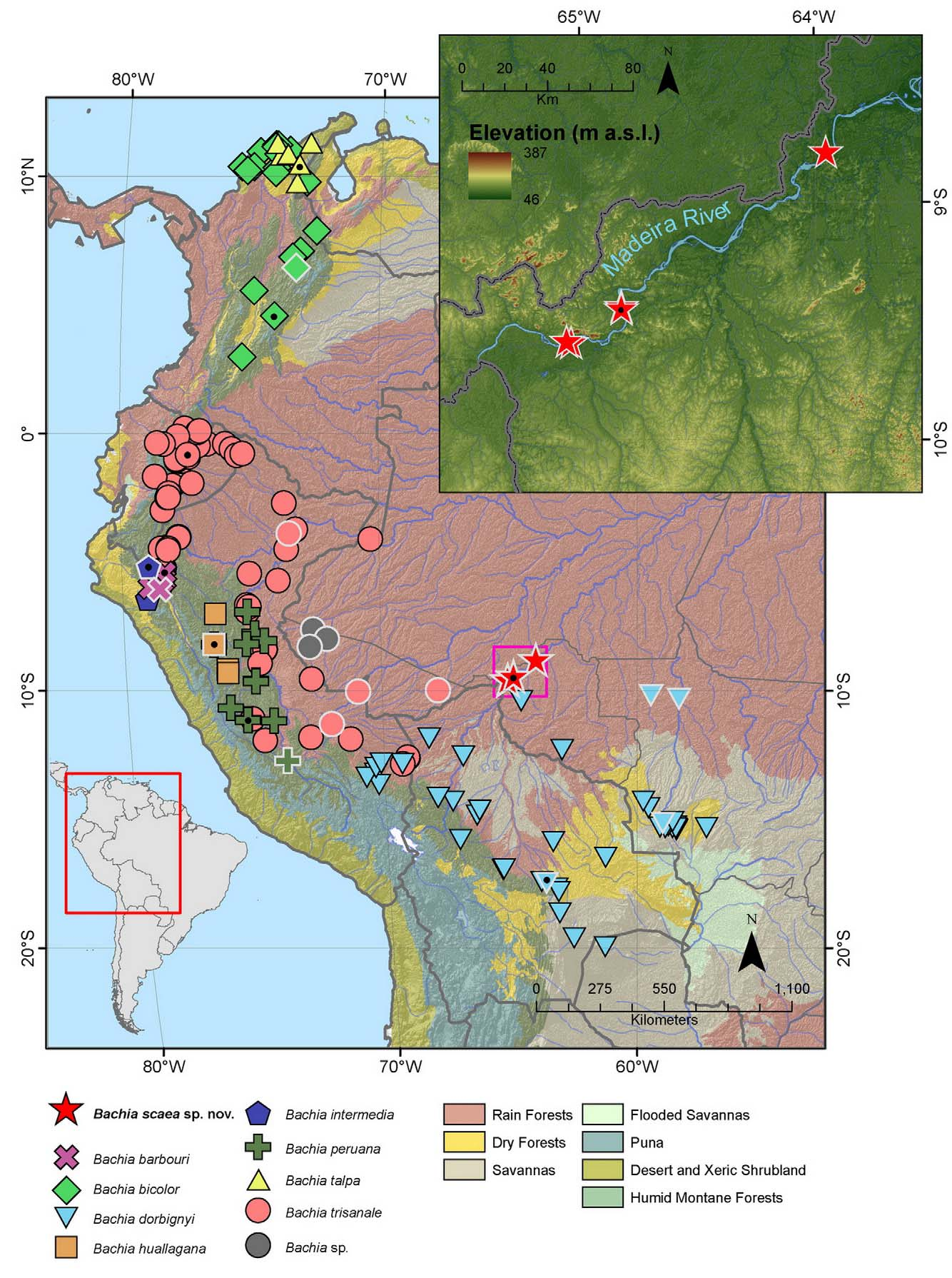

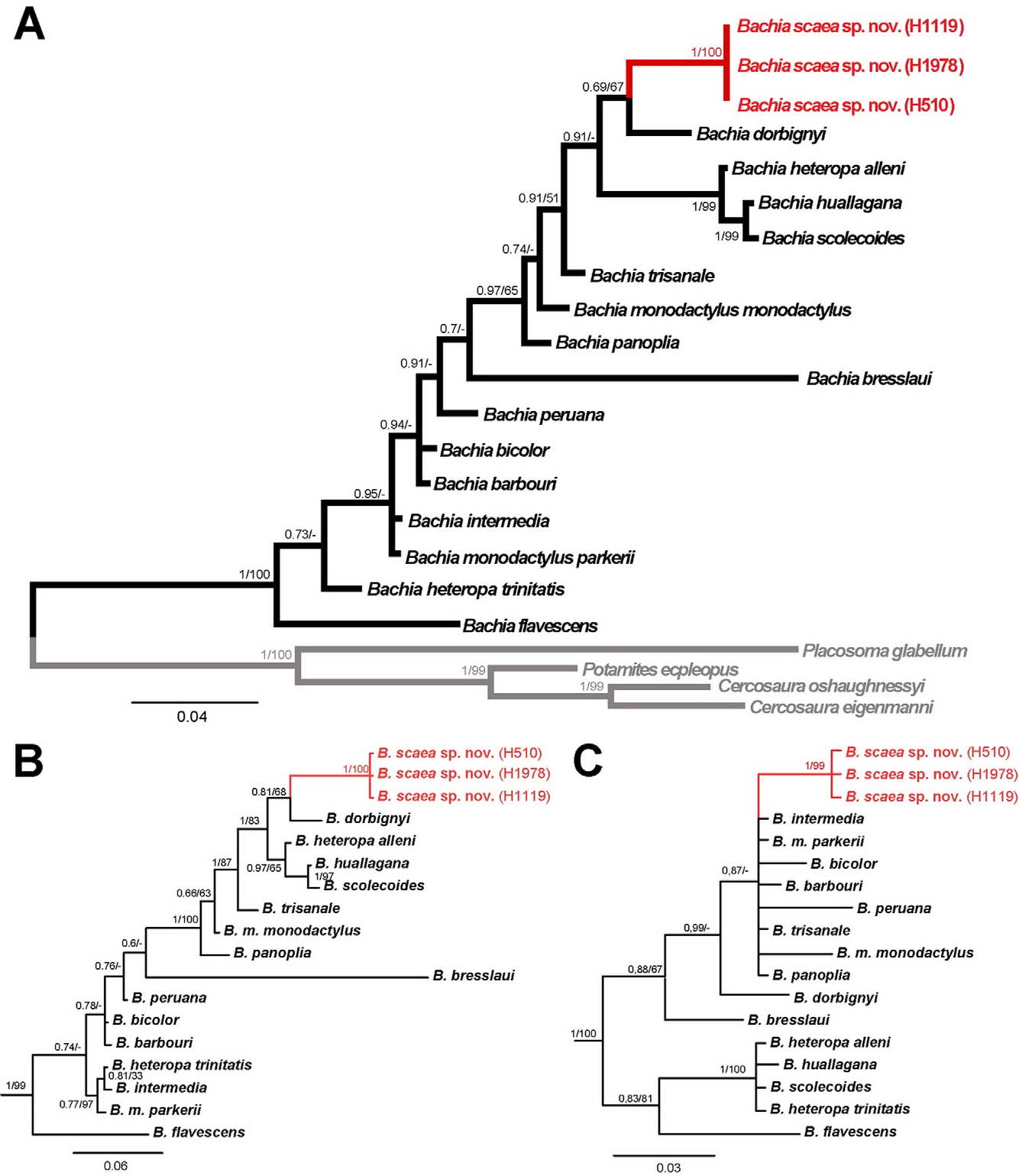
TABLE 1. Summary of diagnostic features among Bachia species of the B. dorbignyi group, derived from examined specimens (see Appendix I) and data from the literature (Cope 1896; Dixon 1973; Presch 1978; Nunes 2011). SAB = scales around body; SL = number of supralabials; SL-P = the supralabial scale contacting parietal; Chin-oral = second chin shield reaching oral border.
| Max SVL Claws | 69 mm none | 68 mm 2 | 75 mm 4 | 80 mm 3 | 73 mm 4 | 67 mm 3 | 107 mm 3 | 65 mm 3 | 79 mm 3 |
|---|---|---|---|---|---|---|---|---|---|
| Frontonasal SAB | present 24–26 | present 26–31 | present 27–31 | present 21–28 | present 24–30 | present 28–35 | present 23–26 | present 25–27 | absent 23–30 |
| Dorsal | 51–54 | 46–52 | 46–53 | 48–55 | 47–54 | 46–52 | 53–57 | 47–51 | 47–55 |
| Ventral | 40–43 | 34–39 | 34–40 | 37–42 | 35–42 | 33–38 | 41–45 | 36–38 | 34–42 |
| Gular | 6–7 | 8–10 | 7–9 | 6–7 | 7 | 7–9 | 6–7 | 9 | 7–8 |
TABLE 2. Pairwise genetic distances for 16 S gene (above) and c-mos (below) among sequences of Bachia analyzed. Values highlighted in bold represented those for the new species.
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1) B. barbouri | 0.00 | 0.11 | 0.08 | 0.06 | 0.06 | 0.01 | 0.07 | 0.01 | 0.05 | 0.02 | 0.05 | 0.01 | 0.08 | 0.08 | 0.08 | 0.08 | 0.07 |
| 2) B. bicolor 0.01 | 0.11 | 0.08 | 0.06 | 0.06 | 0.01 | 0.07 | 0.02 | 0.05 | 0.01 | 0.05 | 0.01 | 0.08 | 0.08 | 0.08 | 0.07 | 0.06 | |
| 3) B. bresslaui 0.03 | 0.03 | 0.11 | 0.12 | 0.12 | 0.12 | 0.12 | 0.12 | 0.11 | 0.12 | 0.11 | 0.11 | 0.12 | 0.12 | 0.12 | 0.13 | 0.12 | |
| 4) B. dorbignyi 0.02 | 0.02 | 0.03 | 0.10 | 0.03 | 0.09 | 0.03 | 0.09 | 0.05 | 0.09 | 0.05 | 0.07 | 0.04 | 0.04 | 0.04 | 0.04 | 0.04 | |
| 5) B. flavescens 0.06 | 0.05 | 0.05 | 0.06 | 0.09 | 0.06 | 0.10 | 0.06 | 0.08 | 0.06 | 0.09 | 0.07 | 0.09 | 0.09 | 0.09 | 0.10 | 0.10 | |
| B. heteropa 6) 0.05 alleni | 0.05 | 0.05 | 0.05 | 0.04 | 0.08 | 0.01 | 0.08 | 0.03 | 0.07 | 0.03 | 0.05 | 0.05 | 0.05 | 0.05 | 0.01 | 0.03 | |
| B. heteropa 7) 0.05 trinitatis | 0.05 | 0.05 | 0.05 | 0.04 | 0.00 | 0.08 | 0.00 | 0.06 | 0.00 | 0.06 | 0.02 | 0.09 | 0.09 | 0.09 | 0.09 | 0.08 | |
| 8) B. huallagana 0.06 | 0.06 | 0.05 | 0.06 | 0.04 | 0.00 | 0.00 | 0.08 | 0.03 | 0.08 | 0.04 | 0.06 | 0.06 | 0.06 | 0.06 | 0.00 | 0.03 | |
| 9) B. intermedia 0.00 | 0.01 | 0.03 | 0.02 | 0.05 | 0.05 | 0.05 | 0.05 | 0.06 | 0.01 | 0.06 | 0.03 | 0.09 | 0.09 | 0.09 | 0.09 | 0.08 | |
| B. m. 10) 0.02 monodactylus | 0.02 | 0.03 | 0.03 | 0.04 | 0.04 | 0.04 | 0.04 | 0.01 | 0.06 | 0.02 | 0.04 | 0.05 | 0.05 | 0.05 | 0.04 | 0.02 | |
| 11) B. m. parkerii 0.00 | 0.01 | 0.03 | 0.02 | 0.05 | 0.05 | 0.05 | 0.05 | 0.00 | 0.01 | 0.06 | 0.02 | 0.09 | 0.09 | 0.09 | 0.09 | 0.08 | |
| 12) B. panoplia 0.00 | 0.01 | 0.03 | 0.02 | 0.05 | 0.05 | 0.05 | 0.05 | 0.00 | 0.01 | 0.00 | 0.04 | 0.05 | 0.05 | 0.05 | 0.04 | 0.03 | |
| 13) B. peruana 0.01 | 0.01 | 0.03 | 0.02 | 0.05 | 0.05 | 0.05 | 0.06 | 0.01 | 0.02 | 0.01 | 0.01 | 0.07 | 0.07 | 0.07 | 0.06 | 0.06 | |
| B. scaea 14) 0.02 sp. nov. (H1119) | 0.02 | 0.04 | 0.02 | 0.06 | 0.06 | 0.06 | 0.06 | 0.01 | 0.03 | 0.01 | 0.01 | 0.01 | 0.00 | 0.00 | 0.06 | 0.05 | |
| B. scaea 15) 0.02 sp. nov. (H1978) | 0.02 | 0.04 | 0.02 | 0.06 | 0.06 | 0.06 | 0.06 | 0.01 | 0.03 | 0.01 | 0.01 | 0.01 | 0.00 | 0.00 | 0.06 | 0.05 | |
| B. scaea 16) 0.02 sp. nov. (H510) | 0.02 | 0.04 | 0.02 | 0.06 | 0.06 | 0.06 | 0.06 | 0.01 | 0.03 | 0.01 | 0.01 | 0.01 | 0.00 | 0.00 | 0.06 | 0.05 | |
| 17) B. scolecoides 0.05 | 0.05 | 0.05 | 0.05 | 0.04 | 0.00 | 0.00 | 0.00 | 0.05 | 0.04 | 0.05 | 0.05 | 0.05 | 0.06 | 0.06 | 0.06 | 0.04 | |
| 18) B. trisanale 0.00 | 0.01 | 0.03 | 0.02 | 0.05 | 0.05 | 0.05 | 0.05 | 0.00 | 0.01 | 0.00 | 0.00 | 0.01 | 0.01 | 0.01 | 0.01 | 0.05 | |
| Discussion |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.