
Lema (Lema) cyanella ( Linnaeus, 1758 )
|
publication ID |
https://doi.org/ 10.1080/00222933.2011.559594 |
|
persistent identifier |
https://treatment.plazi.org/id/03FF8795-1F13-FFB2-FE72-4ECB84A3FAD1 |
|
treatment provided by |
Felipe |
|
scientific name |
Lema (Lema) cyanella ( Linnaeus, 1758 ) |
| status |
|
Lema (Lema) cyanella ( Linnaeus, 1758)
( Figures 1B View Figure 1 , 6–9 View Figure 6 View Figure 7 View Figure 8 View Figure 9 )
Chrysomela cyanella Linnaeus, 1758: 376 View Cited Treatment [Europe] (LCL).
Crioceris cyanella: Fabricius, 1775: 121 .
Lema cyanella: Fabricius, 1798: 93 ; Winkler, 1924 –1932:? [ Japan].
Lema rugicollis Suffrian, 1841: 97 [ Germany] (?), treated as a synonym of L. puncticollis in Jacoby and Clavareau, 1904: 5; treated as a synonym of L. cyanella in Clavareau, 1913: 61; Monrós, 1959: 182; Warchalowski, 1985: 191; Schmitt, 2010:362.
Lema obscurior Pic, 1897: 3 [?] (MNHN), synonymized by Monrós (1959); Schmitt (2010). Since we have not detected the original paper written by Pic yet, according to Monros (1959) and Schmitt (2010) we cited the species in the list.
Lema inaequalicollis Pic, 1924: 13 [ China] (MNHN), syn. nov.
Lema cyanella ab. nigricans Jacobs, 1926: 166 [ Germany] (?) [unavailable name].
Lema concinnipennis var. ventralis Kuwayama, 1932: 69 [Kôyadaira, Prov. Awa] (SEHU), partim, syn. nov.
Lema (Lema) cyanella: Monrós, 1959 ; Mohr, 1985: 243.
Lema nigricans Monrós, 1959: 181 , [unavailable name].
Lema concinnipennis: Kimoto, 1964: 127 [Senkaku islands].
Japanese name: Sasaki-kubiboso-hamushi.
Specimens examined
Lectotype of Lema (Lema) concinnipennis var. ventralis (designated here): // Shikoku: Awa / Koyadaïra; / Japon: 11-8-13 / Edme Gallois // Lema concinnipennis / BALY var. ventralis / KUWAYAMA (1931) / Det. SATORU KUWAYAMA / COTYPE // (SEHU). [Although the author wrote “Cotype” on the identified label, he did not designate a cotype in Kuwayama (1932).]
Paralectotype of Lema (Lema) concinnipennis var. ventralis (designated here), partim. 1 ♂ // Shikoku: Awa / Koyadaïra; / Japon: 11-8-13 / Edme Gallois //; 1 ♀ // Shikoku: Awa / Koyadaïra ; / Japon: 27-7-13 / Edme Gallois //; 1 ♀ // Shikoku: Awa / Koyadaïra ; / Japon: 4-8-13 / Edme Gallois //; 1 ♀ // Shikoku: Awa / Koyadaïra ; / Japon: 9-8-13 / Edme Gallois //; 1 ex. // Shikoku: Awa / Koyadaïra ; / Japon: 4-8-13 / Edme Gallois //.
Holotype specimen of Lema inaequalicollis Pic, 1924 ( China) . 1 ♀ [questionable], // type // Chine bor chi fou // inaequalicollis n. sp. [Photographs taken by Takuma Yoshida in MNHN and a description were investigated.]
Others. [Europe] Bulgaria: 1 ♂, 23 April 1971, R. Druma [ KM] ; 1 ♂, Razgiad s. Senovo, 6 May 1972 [ KM] ; 1 ♂, Strand ja Zvesdez, 3 June 1968, V. Tomov [ KM]. Czech Republic : 1 ♀ [questionable], Chlum u Treb., Bohemia [ JPC]. [ East Asia, Mongolia] : 1 ♂, Inn Shan [ KM]. [ North-eastern China] 1 ♂, 30 May 1938, Mokasai, I. Okada [ SEHU]. [ East Asia, Senkaku islands ] 2 ♂, 3 April 1951, T. Takara [ KM]. [ Honshu , Japan] Aichi Pref. : 1 ♂, Shikatsu-mura, Nishi-kasugai-gun, 20 April 1946, S. Osawa [ SEHU]. Kyoto Pref. : 1 ♂, 1 ♀, Sayama, Kumiyama-chô, 5 May 2007, H. Suenaga [ SHC]. [ Shikoku , Japan] Ehime Pref. : 1 ♂, Iwayaji, 1 June 1969, M. Sakai [ EUM] ; 1 ex., Yotsuga-mine, 18 July 1947, M. Miyatake [ EUM]. [ Kyushu , Japan] Ôita Pref. : 1 ♂, 2 ♀, Kamitsue-machi, Hita-shi, 6–8 May 2009, S. Sasaki [ SSC] ; 1 ♂, 3 ♀, 2–4 June 2009, same locality and collector as the preceding [ SSC] ; 2 ♂, 28–30 August 2009, same locality and collector as the preceding [ SSC]. Nagasaki Pref. : 1 ♂, Oonobaru, Higashi-sonogi, 30 August 2008, S. Imasaka [ ISC] ; 1 ♂, 25 June 2009, same locality and collector as the preceding [ ISC] .
Diagnosis
Body nonparallel and very slightly swollen at middle in dorsal view. Vertex swollen. Frontoclypeus bearing relatively long setae on two posterior sides. Ventral surface of thorax and abdominal segment 1 with coarse setation.
Redescription
Body coloration ( Figure 1B View Figure 1 ). Dorsal: head throughout dark indigo-blue, without red spot on vertex, antenna black; pronotum, elytra and scutellum shining indigo-blue; legs unicolored black. Ventral: black throughout. Setae white. Bulgaria and Senkaku islands individuals with deep metallic green on ventral surface.
Head ( Figures 6A, B View Figure 6 ). Length greater than width; postocular region strongly constricted, rear region forming noticeable neck; vertex groove very deep; top of vertex noticeably swollen, with median fovea elongated longitudinally, its surface covered with fine transverse wrinkles and a few setae arising from large punctures; area between vertex groove and compound eye with large punctures bearing setae; frontoclypeus triangular, enclosed by grooves, bearing relatively long setae on two posterior sides, central region and along anterior suture hairless; labrum with six relatively long setae along posterior margin; antenna filiform, 0.6 times as long as body length, segments 1–2 with a few long setae, segments 3–10 bearing velutinous setation, segments 3 + 4 slightly longer than segment 5, 5th segment longest, segments 1 and 2 globular, each segment stout, segments 3–6 cylindrical but thickening toward apex, segments 7–11 cylindrical and uniform in diameter, apex of segment 11 conically prominent.
Prothorax (lateral and ventral, Figure 6B View Figure 6 ). Slightly wider than long, constriction located a little posterior of the middle; surface bearing punctures of variable size with coarse arrangement and unclear transverse groove near base, fovea at middle, anterior and basal margins narrowly ridged, posterior margin internally bearing close, dense setation. A long seta arising from each anterior and posterior corner.
Mesonotum ( Figure 6B View Figure 6 ). Scutellum trapezoid in shape, posterior corners round; surface closely bearing setae on basal half; posterior and lateral margins of scutellum fringed by bulging. Area right anterior scutellum covered with dense setation.
Elytra ( Figure 1B View Figure 1 ). Solid build in shape, 1.7 times longer than wide; slightly depressed in anterior region but in Ôita population no depression recognizable; strial punctures being subobsolete distally, interstrial area smooth.
Pygidium . Bearing very dense setation except for area with many transverse rows of ridges (stridulatory devices).
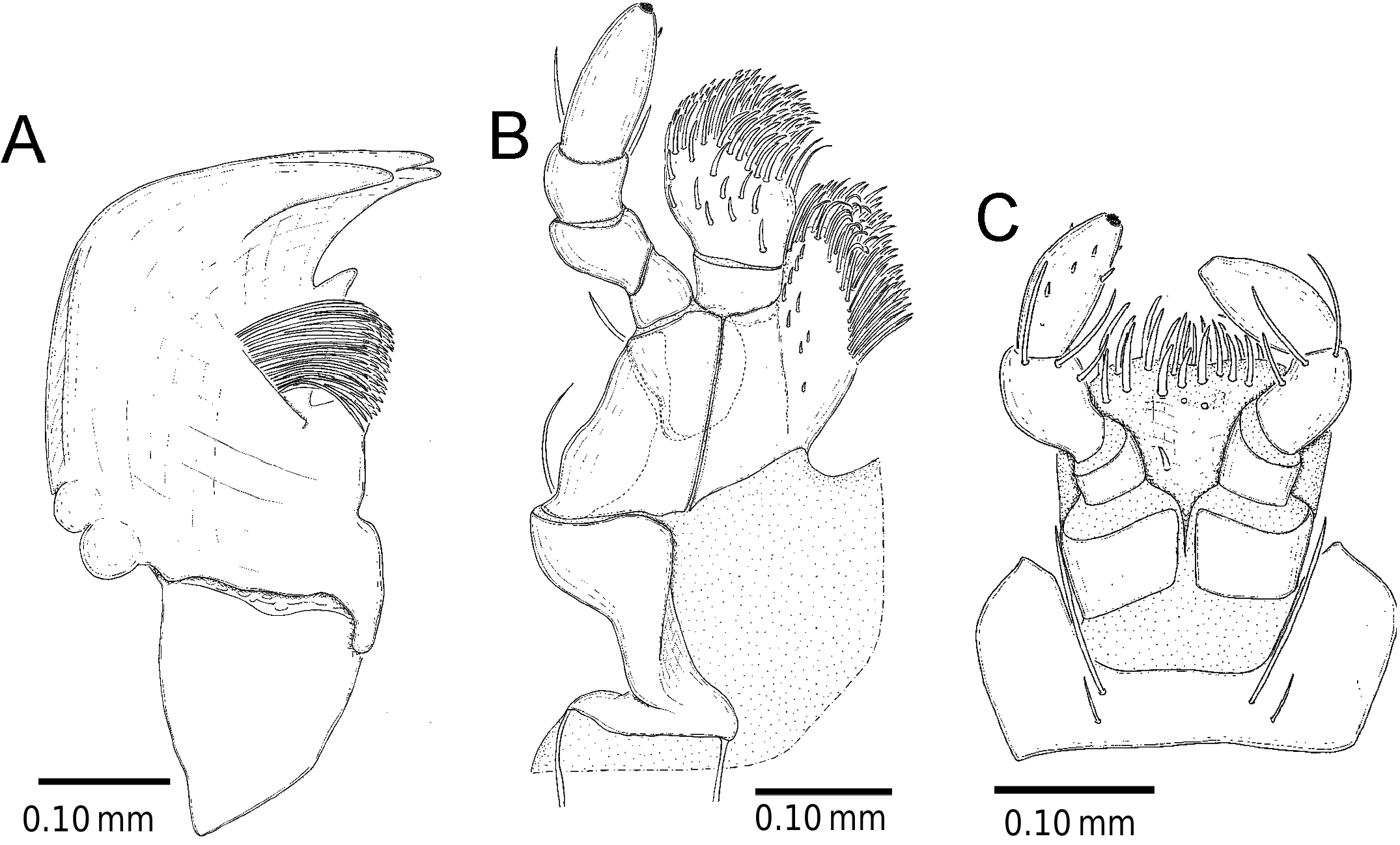
Mouthparts (not all setae were detected) ( Figure 7 View Figure 7 ). Mandible short and stout, strongly sclerotized; anterior region armed with three teeth; mola well-developed; bristles between teeth and mola relatively long, basal half in cluster of bristles curved basally; apodeme relatively short. Maxilla short; palpus five-segmented, segment 1 conico-cylindrical, apex of segment 1 covered with small projections, segments 2–5 cylindrical and short; galea two-segmented, apical segment bearing very thick setae; lacinia bearing stout setae on apical and inner ridge-line; base of cardo with two slen- der apodemes. Labium palpiger four-segmented, apex of palpiger segment covered with small projections; ligula weakly sclerotized, membranous part anterior to ligula covered with very thick setae. Gula and genae wholly covered with relatively coarse setation with large punctures.
Prosternum ( Figure 6C View Figure 6 ). Anterior part of prosternum transversely oblong, basal half bearing relatively denser setation than anterior half; prosternal process very narrow and higher than anterior part, bearing setae on ridge-line, posterior ends widened with dense setation. Surface of pronotal hypomeron smooth except at middle with complex sculpture; posterior margin of pronotal hypomeron not closed and forming arms, but prosternal process bridges them. Anterior and posterior ends of prothorax with setae on fringe: anterior with curved seta and posterior with straight one.
Mesothorax ( Figure 6C View Figure 6 ). Surface of mesosternum bearing microsculpture and relatively coarse setation, with ridge along posterior margin. Mesepisternum and mesepimeron covered with relatively dense setation except for basal margin.
Metathorax ( Figure 6C View Figure 6 ). Metasternum oblong with ridge on each side; surface bearing relatively coarse setation, but its density being weaker toward midline where glabrous, anterior ridge coarsely covered by setae, lateral ridge with no seta, lateral half of posterior ridge with setae. Metepisternum with longitudinal groove at middle; surface bearing moderately dense setation, setation of inner half being coarse toward base, lateral half with glabrous area where covered by elytra.
Legs. Forecoxae conico-cylindrical in shape, bearing dense and fine setae, foretrochanters glabrous but with a long sensillum and a few short ones on anterior ridgeline; profemora almost glabrous except lateral apex with coarse setation in ventral view, in dorsal view bearing relatively dense setation except for basal area where glabrous. Midcoxae spherical, bearing dense, short setae on ventral half. Mid- and hindtrochanters glabrous but with a few setae on posterior ridge-line; mid- and hindfemora bearing coarse setation in ventral view, and in dorsal view glabrous except for lateral apex with coarse setation. Tibiae slender but thickening toward apex, covered with dense setation, in basal area with curved setae, in mid to apical area with straight and slightly hardened setae; its apex bordered with spine-like, translucent brown projection, and also armed with a pair of very short spines on ventral, spine longest in midtibiae than other tibiae.
Abdominal sternite ( Figures 6D, E View Figure 6 ). Surface of segment 1 entirely smooth and polished, but lateral margin with dense setation, near middle covered with very coarse setation. Sternites 2 and 3 covered with relatively dense setation but three glabrous patches exist in middle and near lateral areas. Sternites 4 and 5 uniformly covered with relatively dense setation in male, in female with glabrous area at middle of sternite 5.
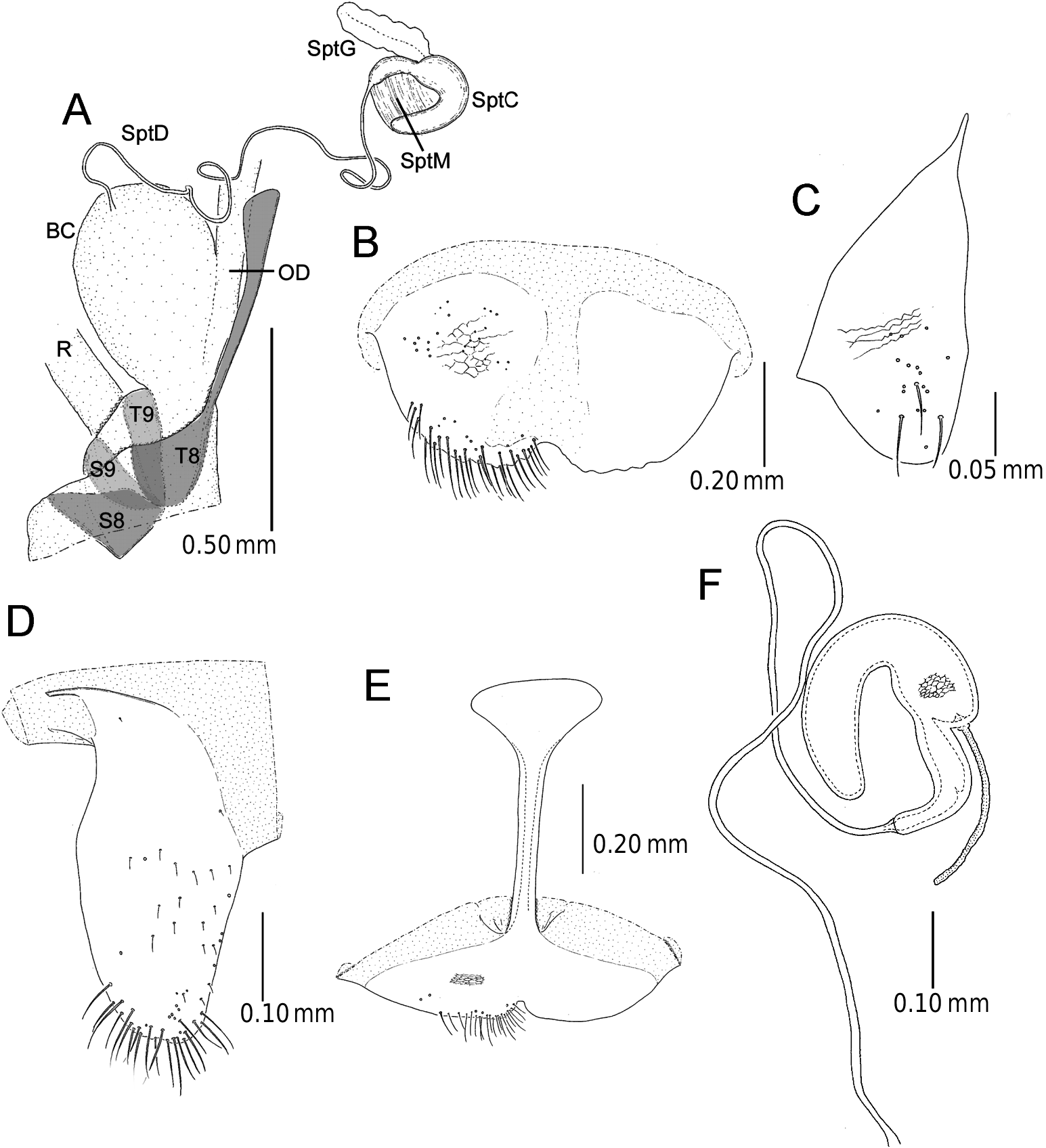
Male genitalia ( Figure 8 View Figure 8 ). Consisting mainly of five parts: tergite 8, second spiculum, tegmen, median lobe and internal sac. Tergite 8 similar to that of female as described below. Second spiculum consisting of two pairs of twig-like sclerites, one pair longer than the other, longer pair asymmetrical. Proximal half of tegmen rectangular in lateral view, tapering toward apex. Median lobe stout, median foramen expanding and occupying one-third of ventral surface in lateral view, ventral surface of median orifice round with no protrusion at its apex. Internal sac showing specialized morphology as in other members of the subgenus Lema ; sclerite having long median ejaculatory guide; basal part of lateral lobe not separated, shape anchor-like in ventral view; membrane having horny process on dorsal side of sclerite.
Female genitalia. ( Figure 9 View Figure 9 ). Consisting of four parts: tergites 8 and 9, and sternites 8 and 9. Tergites 8 and 9 and sternite 9 consisting of a pair of sclerites; sternite 8 with stick-like apodeme; tergites 8 and 9 and base of sternite 8 covered by scale-like or transverse wrinkles. Spermatheca with relatively long duct with no specialized junction to bursa copulatrix. Distal part of spermathecal capsule hook-shaped, inner surface completely covered by scale-like sculpture; proximal part of spermathecal capsule curving at right angle, with spine-like projection on inner wall at middle.
Measurements
Elytral length: male: 3.54 ± 0.17 mm (mean ± SD), female: 3.30 ± 0.14 mm. Elytral width: male: 2.13 ± 0.10 mm, female: 1.94 ± 0.07 mm. Pronotum length: male: 1.14 ± 0.07 mm, female: 1.05 ± 0.08 mm. Pronotum width: male: 1.20 ± 0.06 mm, female: 1.13 ± 0.04 mm. Ratio (length of antennal segments 3 + 4 / 5): 1.33 ± 0.12 (range: 1.11–1.71). Median ejaculatory guide (MEG) length (Japanese population): 2.05 ± 0.20 (range: 1.63–2.05). Spermathecal duct length (Japanese population): 1.90 ± 0.23 (range: 1.62–2.16). MEG length ( Mongolia, n = 1): 2.13 mm, ( Bulgaria, n = 3): 1.60 mm, (Senkaku Island, n = 2): 2.14 mm.
Distribution
Japan (Honshu, Shikoku, Kyushu), Europe, China, Mongolia, Korea, Senkaku Islands. The species is new to the Senkaku Islands.
It has been recorded from the Nearctic region; however, the species was recognized as a previously introduced one (cf. Schmitt (2010).
Host plants
The specimens from Kyushu were collected by sweeping or hand capturing of the beetles from deferent Cirsium plants depending on seasons. The presumed host plants are
Cirsium japonicum ( Figure 1D View Figure 1 ) and C. suffultum from May to June and C. lineare from August to September.
Remarks
Validity of identification of L. (L.) cyanella . Although we did not examine the type specimens of Chrysomela cyanella Linnaeus, 1758 (= L. cyanella ), the descriptions of the specimens examined in the present study from Japan, Bulgaria, the Czech Republic, China and Korea agreed with the original descriptions of L. cyanella and L. puncticollis Curtis, 1830 (a junior synonym of L. cyanella ). In addition, because L. cyanella is the only species of the subgenus Lema recorded from Europe (e.g. Reitter 1912; Warchalowski 1985), we identified the present species, from Japan, Bulgaria, the Czech Republic, China and Korea, as L. cyanella .
Justification for synonymizing the two species with L. (L.) cyanella . Gressitt and Kimoto (1961) synonymized L. inaequalicollis Pic, 1924 with L. concinnipennis Baly, 1865 without providing reasons. We examined photographs of the type specimen of L. inaequalicollis and identified it as L. cyanella . Monrós (1959) synonymized L. concinnipennis var. ventralis with L. concinnipennis without providing reasons. However, the syntypes of L. concinnipennis var. ventralis Kuwayama, 1932 contain two species, L. concinnipennis and L. cyanella , and the lectotype was designated as the latter species in the present study to avoid further nomenclatural confusion. Because the description in Kuwayama (1932) agreed with those of latter species and the syntype belonging to L. cyanella had a “cotype” label, we selected that specimen as the lectotype. Therefore, we propose that L. inaequalicollis and L. concinnipennis var. ventralis are junior synonyms of L. cyanella .
Generic position of L. (L.) cyanella . Crowson (1967) stated that the type specimen of L. cyanella is identical to Oulema lichenis (Voet, 1806) (see White (1981) for nomenclatural problems of O. lichenis ). However, the types of Chrysomela cyanella (= L. cyanella ) are not conspecific with O. lichenis ( Schmitt 1990; M. Schmitt, personal communication in 2010).
Distribution records from Japan. Winkler (1924 –1932) recorded this species from Japan (cf. Özdikmen and Turgut 2008), and Baly (1873) recorded this species as Lema puncticollis from Japan. Winkler’s work has not been mentioned in the taxonomic literature of Japanese Lema species ( Chûjô and Kimoto 1961; Kimoto 1964; Kimoto and Takizawa 1994), and Baly’s report had been considered to be a misidentification ( Chûjô and Kimoto 1961, Kimoto and Takizawa 1994). Chûjô and Kimoto (1961) reclassified Baly’s record of L. puncticollis from Japan as L. cirsicola , although Baly’s specimens were not included in their work. Since we were not able to investigate the materials used by Winkler (1924 –1932) or Baly (1873), the validity of these reports is not judged.
Kuwayama (1932) recorded L. puncticollis from Japan. However, we examined his materials deposited in SEHU, and they were identified as L. cirsicola . Kuwayama (1932) treated Lema sapporensis Matsumura, 1911 as a subspecies of L. cyanella , and Monrós (1959) synonymized L. sapporensis with L. cyanella without providing reasons. We examined his materials deposited in SEHU, including the type specimens of L. sapporensis , and they were concluded to be a different species from L. cyanella . Therefore, we resurrect L. sapporensis from synonymy with L. cyanella .



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Lema (Lema) cyanella ( Linnaeus, 1758 )
| Matsumura, Y., Sasaki, S., Imasaka, S., Sano, M. & Ôhara, M. 2011 |
Lema (Lema) cyanella: Monrós, 1959
| Mohr K-H 1985: 243 |
Lema concinnipennis: Kimoto, 1964: 127
| Kimoto S 1964: 127 |
Lema nigricans Monrós, 1959: 181
| Monros F 1959: 181 |
Lema concinnipennis var. ventralis
| Kuwayama S 1932: 69 |
Lema cyanella
| Jacobs W 1926: 166 |
Lema inaequalicollis
| Pic M 1924: 13 |
Lema rugicollis
| Schmitt M 2010: 362 |
| Warchalowski A 1985: 191 |
| Monros F 1959: 182 |
| Clavareau H 1913: 61 |
| Jacoby M & Clavareau H 1904: 5 |
| Suffrian E 1841: 97 |
Lema cyanella: Fabricius, 1798: 93
| Fabricius JC 1798: 93 |
Crioceris cyanella:
| Fabricius JC 1775: 121 |