
Mediomastus opertaculeus, Tomioka & Hiruta & Kajihara, 2013
|
publication ID |
https://doi.org/ 10.12782/sd.18.1.105 |
|
publication LSID |
lsid:zoobank.org:pub:95FE7B70-A37B-4F24-8236-9A8FEA2261B2 |
|
persistent identifier |
https://treatment.plazi.org/id/CEEBF297-ACB3-44E2-9BAF-812DBC787671 |
|
taxon LSID |
lsid:zoobank.org:act:CEEBF297-ACB3-44E2-9BAF-812DBC787671 |
|
treatment provided by |
Felipe |
|
scientific name |
Mediomastus opertaculeus |
| status |
sp. nov. |
Mediomastus opertaculeus View in CoL sp. nov.
( Figs 1–5 View Fig View Fig View Fig View Fig View Fig )
Material examined. Holotype: ZIHU 4369 View Materials , whole specimen, fixed in 10% formalin seawater, preserved in 70% ethanol . Paratypes (seven specimens): ZIHU 4370, mount- ed on SEM stub; ZIHU 4371, some parts of body mounted on slides, remaining parts preserved in 70% ethanol; ZIHU 4372–4375, thoracic regions only, fixed in 10% formalin seawater, preserved in 70% ethanol; abdominal parts were used for DNA extraction; ZIHU 4376, whole specimen, fixed in 10% formalin seawater, preserved in 70% ethanol. Other material examined: ZIHU 4377–4379, three whole specimens, fixed in 10% formalin seawater, preserved in 70% ethanol.
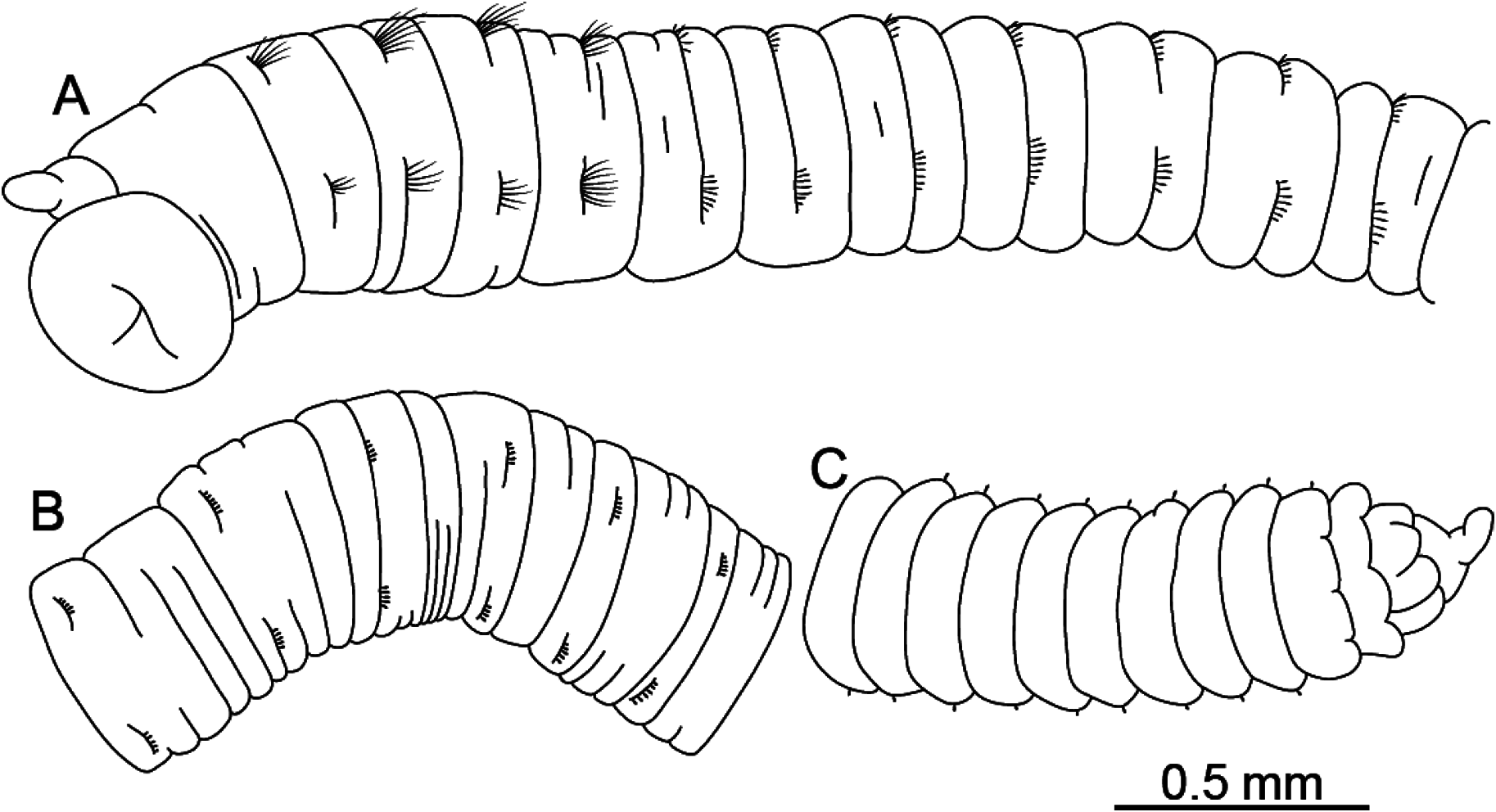
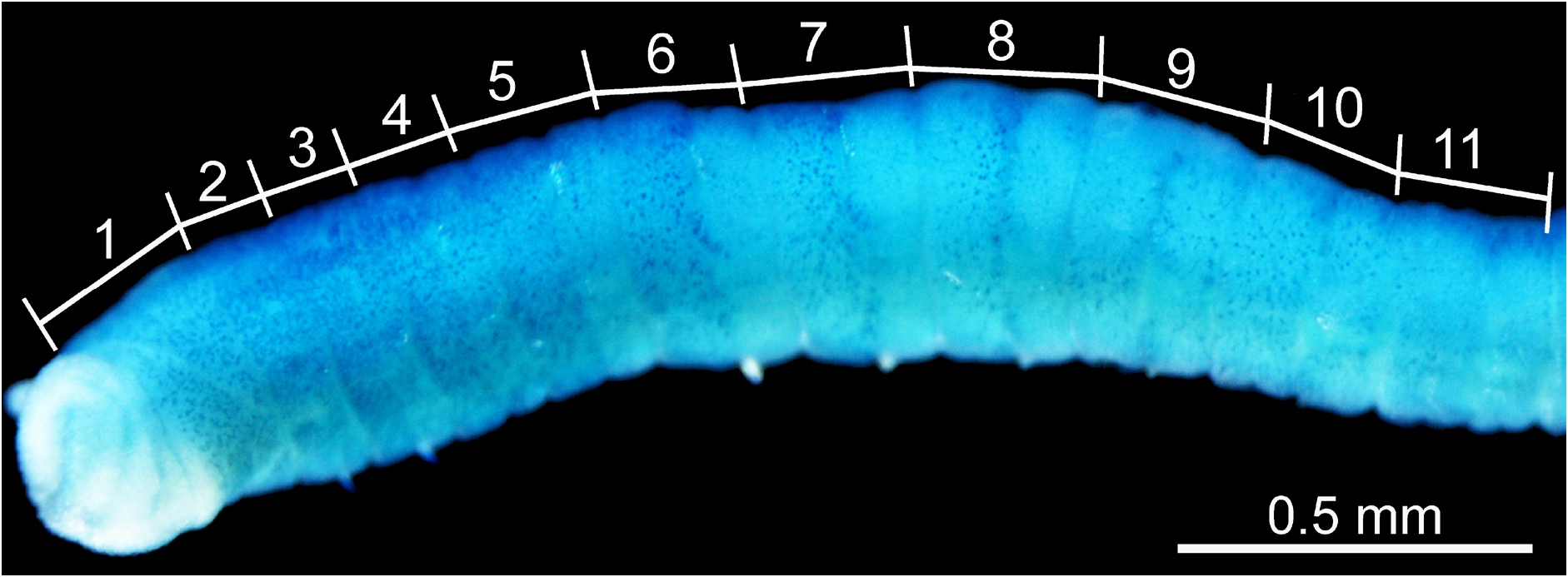
Description. Holotype complete, sex uncertain, with 92 segments; 15.7 mm in length; 0.47 mm in maximum width. Body color red in life, whitish pink in ethanol. Epithelium smooth. Nephridiopores not visible. Branchiae absent. All segments cylindrical.
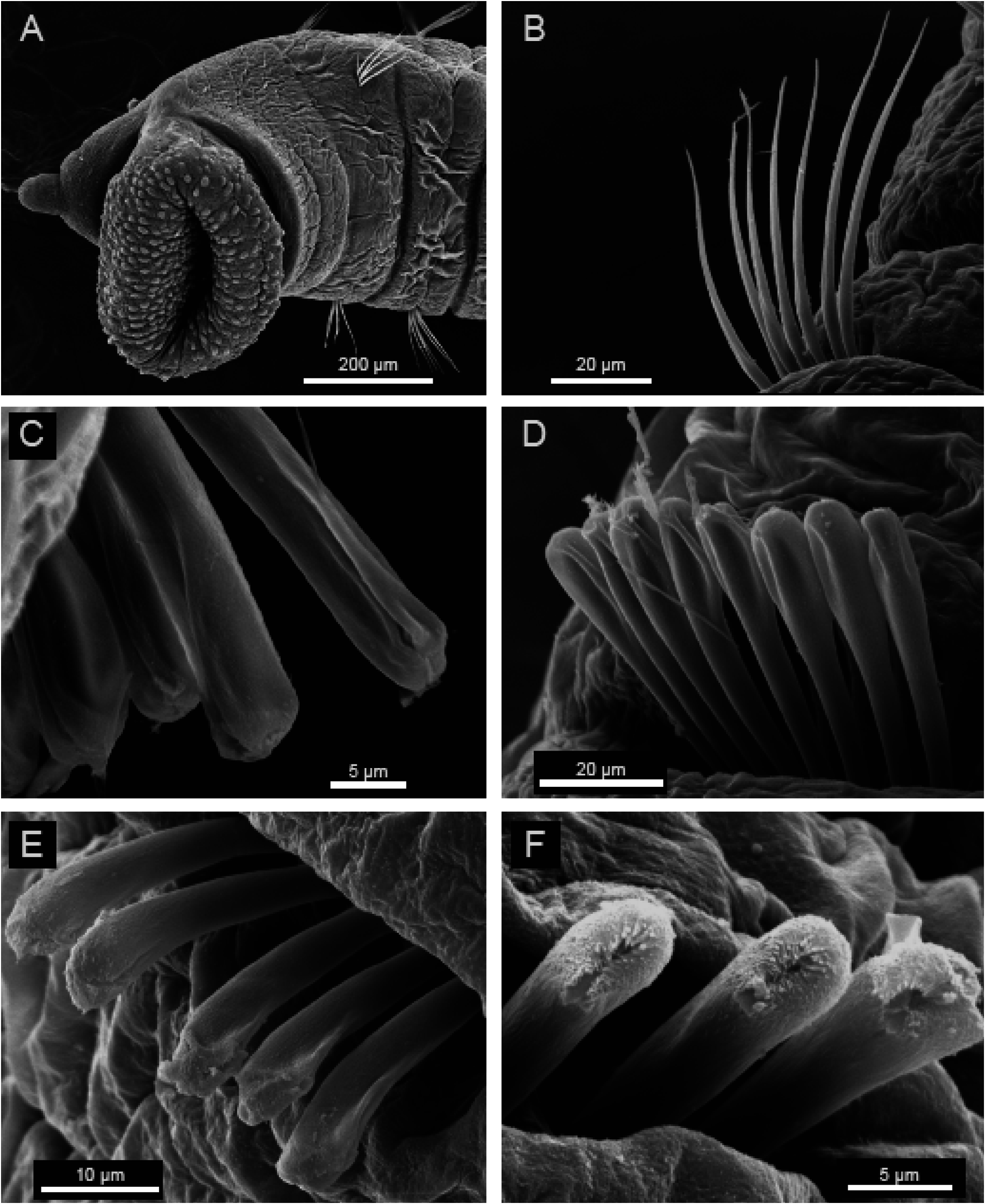
Prostomium ( Fig. 1A View Fig ) conical, with short palpode; eversible proboscis with numerous minute papillae ( Fig. 2A View Fig ). Peristomium 1.7 times as wide as long, slightly longer than chaetiger 1, without eye spots, achaetigerous.
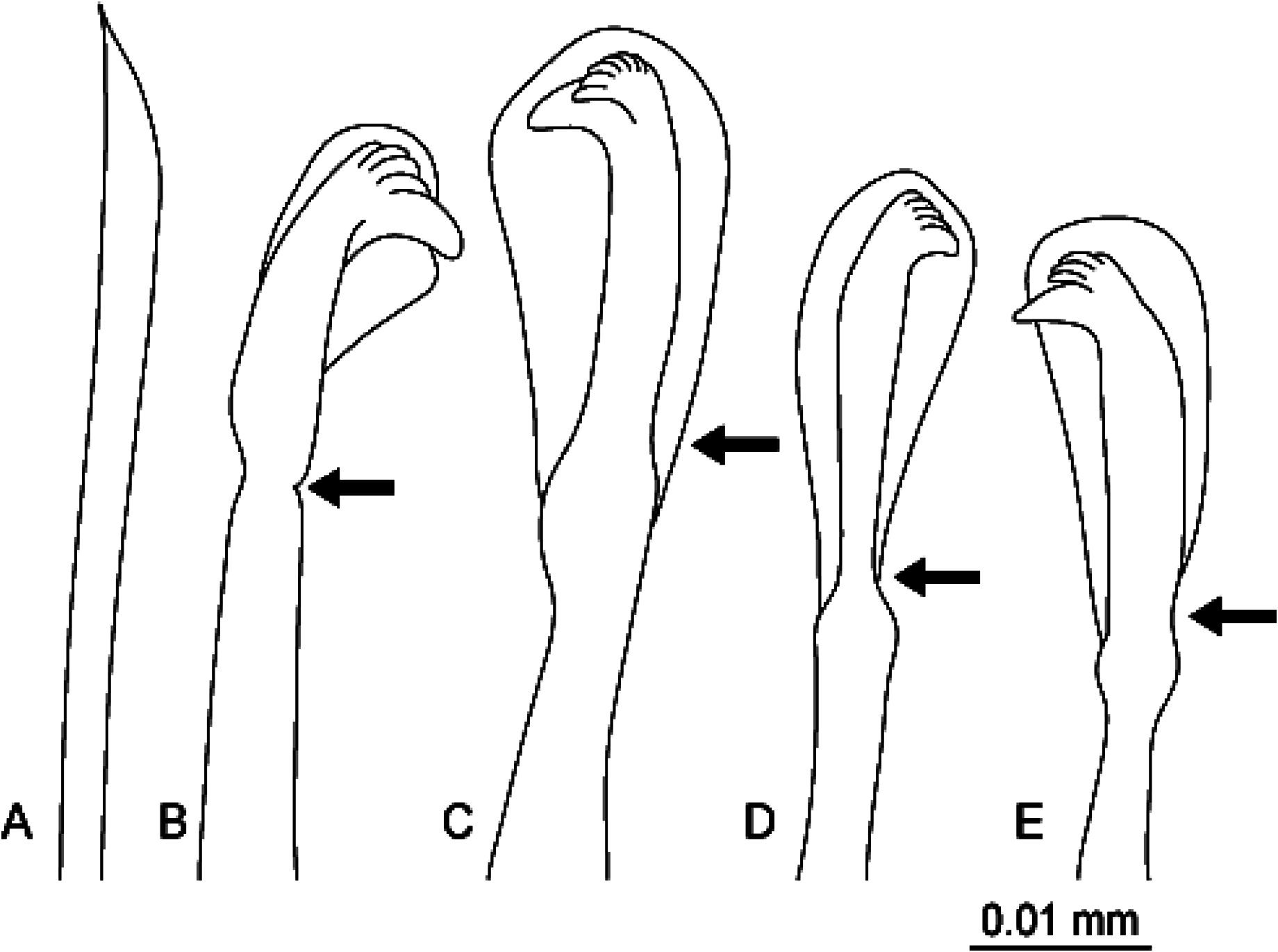
Thoracic chaetigers biannulate. Whip-like capillary chaetae without wing ( Fig. 3A View Fig ) present on chaetigers 1–4; noto- and neurochaetae each 5–10 in number per fascicle ( Fig. 2B View Fig ). Chaetigers 5–10 with hooded hooks, but without paddle-like chaetae. Notopodial hooded hooks with obvious fang ( Fig. 3B View Fig ); hood with small opening; fang with 4 teeth; constriction distinct ( Fig. 3B View Fig , arrowed); 5–8 hooks per fascicle ( Fig. 2C View Fig ). Neuropodial hooded hooks without opening ( Fig. 3C View Fig ); fang with 7 teeth; constriction slight ( Fig. 3C View Fig , arrowed); 5–8 hooks per fascicle ( Fig. 2D View Fig ).
Abdominal segments 1.4 times as wide as long ( Fig. 1B View Fig ); posterior segments becoming narrower and longer than anterior chaetigers; with hooded hooks only. Notopodial hooded hooks with obvious fang ( Fig. 3D View Fig ); hood with opening wider than that of thoracic notopodial hooded hooks; fang with 4 teeth; with distinct constriction ( Fig. 3D View Fig , arrowed); 2–5 hooks per fascicle ( Fig. 2E View Fig ). Neuropodial hood- ed hooks with pointed fang ( Fig. 3E View Fig ); hood with opening wider than that of thoracic notopodial hooded hooks; fang with 4 teeth; with distinct constriction ( Fig. 3E View Fig , arrowed); 2–5 hooks per fascicle ( Fig. 2F View Fig ).
Transition from thorax to abdomen marked by alteration of shape of hooded hooks; neuropodial hooded hooks in thorax without opening on hoods, while those in abdomen with opening on hoods.
Pygidium with short, rounded-cylindrical caudal cirrus ( Fig. 1C View Fig ).
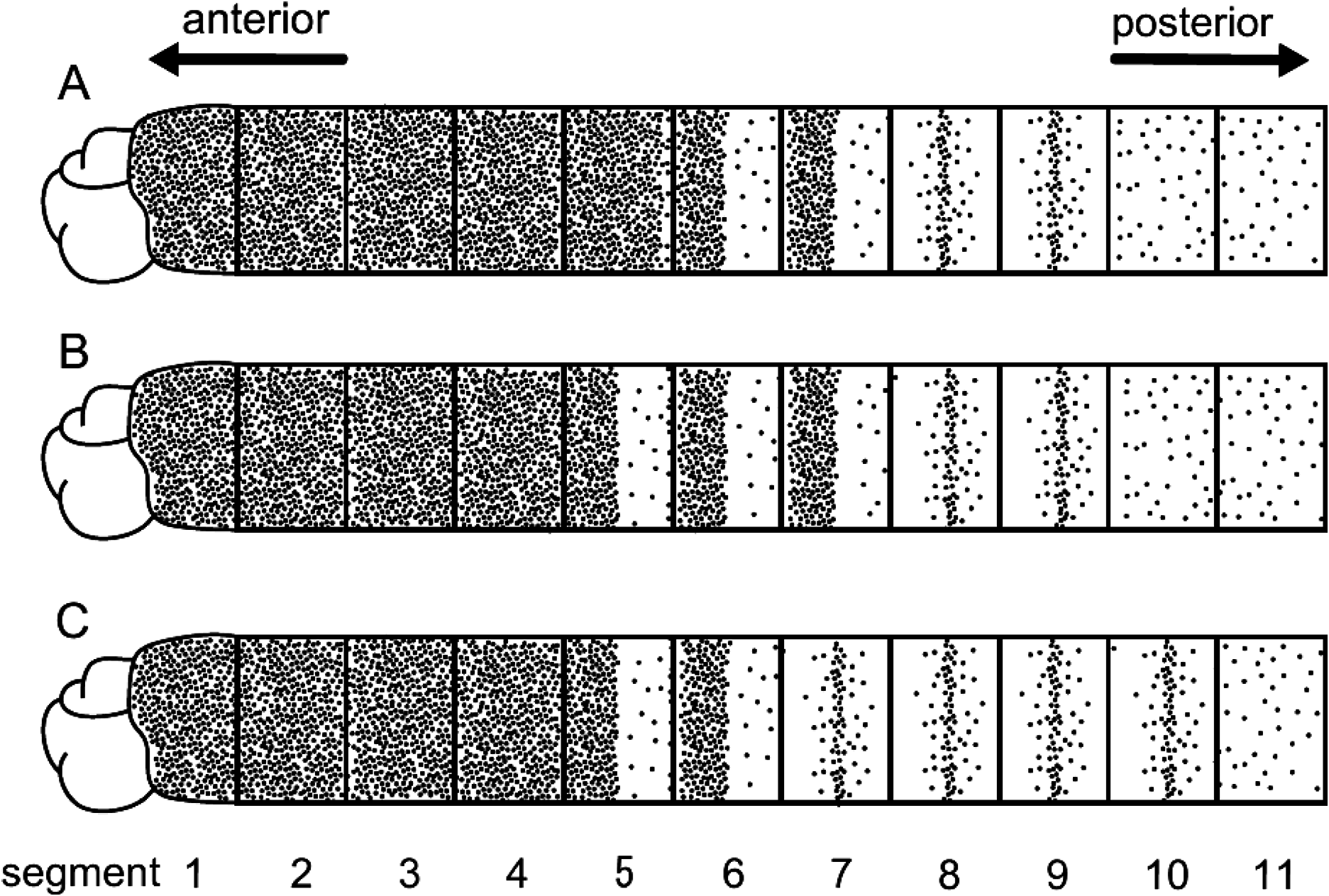
Methyl-green staining. Among nine specimens observed, methyl-green staining resulted in several patterns, three of which are diagrammed in Fig. 4 View Fig . All patterns showed numerous minute spots densely and uniformly covering segments 1–4, though in some individuals, spots completely covered segment 5 as well ( Fig. 4A View Fig ). All patterns showed, immediately posterior to the completely spotted segments, at least two ( Fig. 4A, C View Fig ) and sometimes three ( Fig. 4B View Fig ) segments that had spots densely and uniformly distributed only in the anterior half of each segment. Finally, all patterns showed, immediately posterior to the half-spotted segments, at least two ( Fig. 4A, B View Fig ) but up to four ( Fig. 4C View Fig ) segments having a narrow band of dense spots in the middle of each segment. Fig. 5 View Fig shows a stained worm with a pattern most similar to that illustrated in Fig. 4B View Fig , except that only segment 8 appears to have a middle band of deeply stained spots.
COI sequences. The four sequences obtained from the paratypes differed from each other at 1–4 sites within the aligned 624 bases, giving Kimura (1980) 2-parameter (K2P) distances of 0.002 –0.006. The four paratypes were identical in the translated amino-acid sequences (208 residues).
As of writing, GenBank contained only one COI sequence ( EU835672 View Materials ) for any species in Mediomastus , an unidentified species from Darwin Harbor, Australia (M. J. Neave et al. unpublished data). The sequences from M. opertaculeus differed from that of the Australian Mediomastus sp. at 116–117 sites, giving K2P distances of 0.216 –0.218, which are far greater than 0.0043, an average intraspecific K2P distances in 11 species of Hydroides Gunnerus, 1768 (Serpulidae) from the Pacific ( Sun et al. 2012), indicating that M. opertaculeus and the Australian Mediomastus sp. are not conspecific. Sun et al. (2012) reported the average interspecific K2P distance between 11 species of Hydroides was 0.262, a value slightly greater than that observed between M. opertaculeus and the Australian Mediomastus sp. , possibly strengthening the adequacy of the generic placement of our new species. The amino acid sequence differed at five sites between M. opertaculeus and the Australian Mediomastus sp.
Etymology. The specific name is a noun, from the Latin participle opertus (“hidden”) and the Latin noun aculeus (“thorn”), referring to the species’ diagnostic hooded hooks, in which the hood completely covers the fang.
Remarks. Among 13 species of Mediomastus , nine have been reported to lack both paddle-like chaetae and abdominal capillary chaetae as in M. opertaculeus sp. nov.; these are M. australiensis Warren, Hutchings and Doyle, 1994 ; M. californiensis ; M. cirripes Ben-Eliahu, 1976 ; M. calliopensis Warren, Hutchings and Doyle, 1994 ; M. capensis Day, 1961 ; M. deductus (Pillai, 1961) ; M. fragilis Rasmussen, 1973 ; M. thomassini Warren, Hutchings and Doyle, 1994 ; and M. warrenae ( Warren et al. 1994; Green 2002). Mediomastus opertaculeus differs from these nine species in the staining pattern with methyl green; Warren et al. (1994) presumed that this method would indicate the presence of glandular cells. In M. opertaculeus , segments 1–4 stain with numerous spots, while in the latter nine species the corresponding segments do not stain, except in M. warrenae (only known from the type locality, the Andaman Sea), in which segment 1 remains unstained and segments 2–4 stain variably with numerous spots, or remain unstained. Mediomastus opertaculeus also differs from M. warrenae in the staining pattern in segments 6–11. In M. opertaculeus , the overall staining pattern is variable across segments 6–11, with dense, deeply stained spots occupying the anterior half or middle of each segment; in M. warrenae , each of segments 6–11 stains with numerous, uniformly distributed spots ( Warren et al. 1994; Green 2002).
Mediomastus opertaculeus differs from subsets of congeners in other characters: from M. australiensis , M. californiensis , M. capensis in the absence of nephridiopores ( Hartman 1944; Day 1961; Warren et al. 1994); from M. ambiseta and M. cirripes in the shape of the capillary chaetae [the capillary chaetae in M. opertaculeus lack a wing, whereas those in M. ambiseta and M. cirripes possess a broad wing ( Warren et al. 1994)]; from M. acutus , M. ambiseta , M. branchiferus , M. calliopensis , M. californiensis , and M. fragilis in the lack of an opening in the thoracic neuropodial hooded hooks ( Warren et al. 1994); and from M. warrenae in the absence of constriction of hooded hooks on thorax ( Green 2002).
| ZIHU |
Zoological Institute, Hokkaido University |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.