
Cephaloleia orchideivora, Sekerka & Windsor & Staines, 2013
|
publication ID |
https://doi.org/ 10.5281/zenodo.5740509 |
|
persistent identifier |
https://treatment.plazi.org/id/03FDA67E-FFBC-0123-A99E-FEA7FC3178C3 |
|
treatment provided by |
Marcus |
|
scientific name |
Cephaloleia orchideivora |
| status |
sp. nov. |
Cephaloleia orchideivora sp. nov.
Type material. HOLOTYPE, J, glued: ‘ PANAMA: Chiriqui | LaFortuna ; 1200m | 8°45’N; 82°15’W | 12 - V [handwritten] -199[printed]7[handwritten] | D.M. Windsor [white, printed and cardboard label]’ ( USNM) GoogleMaps . PARATYPES: PANAMA: CHIRIQUÍ: 2 paratypes, glued: with same label as holotype, one with following additional label ‘ex. Elleanthus | Orchidac. [white, handwritten and cardboard label]’ ( DWCP) ; paratype, glued: ‘ PANAMA: Chiriqui | LaFortuna ; 1200m | 8°45’N ; 82°15’W | 20 - I [handwritten] -199[printed]7[handwritten] | D.M. Windsor [white, printed and cardboard label]’ ( DWCP) ; paratype, glued: ‘ PANAMA: Chiriqui | La Fortuna, 9-1100m | Rio Hornito Trail | 8°45’N, 82°14’W | 15 - II [handwritten] -199[printed]8[handwritten] | D.M. Windsor [white, printed and cardboard label] || Orchid [white, handwritten and soft label]’ ( DWCP) GoogleMaps ; 2 paratypes, glued: ‘ PANAMA, Chiriqui | Fortuna, Rio Hornito | Tr, N 8 47’, W 82 13’ | 1100m, 21-vii-2009 | D Windsor ; L Sekerka [white, printed and cardboard label] || ex Elleanthus | Orchidac. [white, handwritten and soft label]’ ( DWCP) ; paratype, in ethanol: ‘ Epidendrum werkli | 21 VII 09 CRH [= Camino Río Hornito ] [white, handwritten and cardboard label]’ ( DWCP) ; paratype, glued: ‘ PANAMA Chiriquí prov. | La Fortuna - Rio Hornito trail | 8°47’N, 82°13’W, 1100 m | cloud forest 21.vii.2009 | ORCH: Epidendrum sp. | L. Sekerka & D. Windsor lgt. [green, printed and cardboard label]’ ( LSCL) GoogleMaps ; 10 paratypes, glued and one in ethanol: ‘ PANAMA: Chiriqui | La Fortuna, 9-1100m | Rio Hornito Trail | 8°45’N, 82°14’W | 12 - II [handwritten] -199[printed]2010[handwritten] | D.M. Windsor [white, printed and cardboard label]’ ( DWCP, UPPC) GoogleMaps ; 5 paratypes, glued: ‘ PANAMA Chiriquí prov. | La Fortuna: Rio Hornito | 8°42.58’N, 82°13.48’W, 1180 m | montane forest | ORCH: Epidendrum wercklei | L. Sekerka lgt. 3.-4.x.2010 [green, printed and cardboard label]’ ( BMNH, LSCL, NMPC) GoogleMaps ; paratype, in ethanol:‘ PANAMA, CHIRIQ | Quijada, Fortuna | 1200 m., | Windsor-200[printed]1-AG-5 | ex. orchid [white, handwritten and cardboard label with black frame]’ ( DWCP) ; 3 paratypes, glued: ‘ PANAMA Chiriquí prov. | La Fortuna: Quijada del Diablo | 8°42.07’N, 82°13.36’W, 1200 m | montane forest | ORCH: Epidendrum wercklei | L. Sekerka lgt. 5.ix.2010 [green, printed and cardboard label]’ ( LSCL) GoogleMaps ; 3 paratypes, glued: ‘ PANAMA Chiriquí prov. | La Fortuna: Quijada del Diablo | 8°42.07’N, 82°13.36’W, 1200 m | montane forest | ORCH: Epidendrum wercklei | L. Sekerka lgt. 8.ix.2010 [green, printed and cardboard label]’ ( LSCL) GoogleMaps ; paratype, glued: ‘ PANAMA Chiriquí prov. | La Fortuna: Quijada del Diablo | 8°42.07’N, 82°13.36’W, 1200 m | montane forest | ORCH: Epidendrum wercklei | L. Sekerka lgt. 7.x.2010 [green, printed and cardboard label]’ ( LSCL). BOCAS DEL TORO: paratype, in ethanol: ‘ PANAMA, Bocas d T | Palo Seco Tr. Km 63 | 8 48’ N GoogleMaps ; 82 13’W | 18 ix 2008 | D Windsor; L Sekerka [white, printed and cardboard label]’ ( DWCP) ; paratype, in ethanol: ‘ Panama | B.d. T. | 900 m | 3 II 2007 | D.W. | Cephaloleia | irregularis [white, handwritten and cardboard label]’ ( DWCP). COCLÉ: paratype, glued: ‘ Panamá: Coclé Prv. | 9-1000m, Caracoral | 8°37’N, 80°7’W GoogleMaps ; | 27 March 1993 | D.M. Windsor [white, printed and cardboard label]’ ( LSCL) ; paratype, glued: ‘ PANAMA: Cocle Prv. | La Mesa ab. El Valle | Cerro Caracoral | 15 Nov 92 ; el. 850 m | col. H. Stockwell [white, printed and cardboard label]’ ( DWCP). PANAMÁ: two paratypes, glued: ‘PANAMA Panamá prov. | PN Chagres: Cerro Jefe | 9°13.7’N, 79°23.0’W, 950 m | montane forest | ORCH: Epidendrum sp. | L. Sekerka lgt. 22.x.2010 [green, printed and cardboard label]’ ( LSCL) GoogleMaps ; paratype, glued: ‘ Cerro Campana, 800 m | Panamá Prov. , R. P. | 29 Apr. ’70 [handwritten] | H. P. Stockwell [white, printed and cardboard label] || ♀ [white, printed and cardboard label] ( USNM) .
Larval material. Larvae were collected from three different sites (Quijada del Diablo, El Vivero and Samudio Trail), all between 1100 and 1200 m elevation in the La Fortuna Reserve. One mature larva (21.vii.2009, DWCP) and two mature and four younger instars (5.ix.2010, LSCL and DBET) were collected from these sites and preserved in ethanol. One mature larva collected on Cerro Jefe (22.x.2010, DWCP) and one recently emerged, first instar larva from Quijada del Diablo (23.xii.2012, DWCP) were cleared in KOH and slide mounted.
Adult diagnosis. The new species is most similar to Cephaloleia irregularis Uhmann, 1930 described from a mountainous locality, Coronado (1400–1500 m), Limón province, Costa Rica. Cephaloleia orchideivora sp. nov. differs by the mostly regularly punctate elytra (irregularly in C. irregularis ), the completely yellow and broad explanate margin of elytra (narrow and dark metallic brownish-green in C. irregularis ), the anterior margin of pronotum emarginate (straight in C. irregularis ), and the pronotum with triangular metallic spot (reverse V-shaped in C. irregularis ).
Description of adult. Measurements (n = 16): length of body 5.48–6.89 mm (mean 6.20 mm), width of body 2.41–3.11 mm (mean 2.69 mm), length/width of body ratio 2.14–2.42 (mean 2.31), length of pronotum 1.06–1.37 mm (mean 1.23 mm), width of pronotum 1.55–2.08 mm (mean 1.82 mm), width/length of pronotum ratio 1.43–1.53 (mean 1.48). Body elongate-oval and subconvex ( Figs 1, 3 View Figs 1–3 ).
Head metallic olive-green sometimes with bluish or violet reflection. Mouthparts, antennal insertions and first antennomere rust-colored, remaining antennomeres black. Pronotum yellow (dry specimens) to pink (live and ethanol preserved specimens) with large triangular olive-green spot along midlength reaching to basal and apical margins. The spot is usually with more or less distinct lateral emargination. Scutellum and elytral disc olive-green, explanate margin yellow to penultimate interspace. Thorax mostly black with episterna brownish or yellowish. Abdomen brown-black variegated with distinct broad yellow outer margin. Coxa rust-colored, trochanters pitchy-brown, tibia and femur internally yellow and externally black, tarsi black.
Pronotum transverse, 1.5 times broader than long, anterior margin deeply emarginate behind head, emargination in the middle convex. Anterior angles of pronotum broadly rounded, lateral margins almost parallel-sided, slightly widened in the basal third in some specimens. Tubercle with sensila situated in corners of anterior emargination. Disc of pronotum coarsely and sparsely punctate with large shiny impunctate areas. Punctures deeply impressed, foveolate. Lateral margin only slightly explanate, distinctly separated from disc by deep impression and extreme outer margin swollen. Surface smooth and shiny, area with metallic spot distinctly micro-reticulate.
Scutellum subpentagonal, smooth, impunctate and micro-reticulate.
Elytra feebly convex, flat. Elytral base slightly sinuate, broader than base of pronotum. Humeral angles broadly rounded. Punctation coarse and sparse, mostly regular. Punctures deeply impressed and foveolate. Scutellar row very long, extending to basal fourth. Scutellar row and rows 1–5 and 8–10 regularly arranged in rows, 7 and 8 with numerous confused punctures on lateral slope. Between rows 7 and 8 there is an additional, more or less regular, row of punctures extending from about midlength of elytra to the apex. Interspaces between puncture rows narrow, 0.5–1.0 times as wide as puncture diameter. Intervals broad, 1–2 times wider than puncture diameter, second interval widest, about 4 times wider than puncture diameter. Surface of intervals and interspaces smooth, shiny and micro-reticulate. Punctation gradually coarser from suture to lateral rows. Row 10 twice as coarse as remaining lateral rows. Lateral margin of elytra distinctly explanate, as broad as 1/6 of elytron, slightly declivous. Its surface smooth, shiny and micro-reticulate, sometimes with 1–3 isolated punctures in basal fourth, otherwise impunctate. Extreme outer margin slightly swollen, distinctly serrate, particularly in apical fifth. Denticles obsolete, with short seta on tip. Apex of epipleura bare.
Vertex of head densely and coarsely punctate, punctures along the middle have tendency to form short striae. Frons impunctate but distinctly micro-reticulate. Clypeus very short, typical for Cephaloleia . Eyes normal, gena well visible. Antennae, long, filiform, moderately thick and reaching beyond humerus. Pedicel and scapus cylindrical, smooth and shiny, only sparsely pubescent; remaining antennomeres elongate, densely setose and micro-reticulate. Third antennomere 2.3 times longer than second and 1.5 times than fourth. Terminal antennomere 1.6 times longer than 10th with rounded apex. Length ratio of antennomeres: 100: 111: 257: 175: 167: 150: 155: 147: 152: 133: 217. Prosternal process broad, between coxae smooth and shiny with deep sulcus along midlength, apex broadly explanate, micro-reticulate and with 7–9 sulci. Mesothorax smooth, micro-reticulate. Metathorax mostly smooth and shiny, its surface appears polished and with several punctures situated basolaterally, each puncture with moderately long golden seta. Metepisterna coarsely punctate. Abdomen finely and densely punctate and covered with adherent setae. Suture between abdominal sterna 1 and 2 complete. Last visible sternite slightly emarginate in male and deeply emarginate in female.
Legs short, robust, claws divergent and simple.
Variability of adults. Individuals from the Chiriquí population ( Fig. 1 View Figs 1–3 ) are relatively uniform with minor variability present only in the shape and size of the pronotal spot and elytral serration. Some specimens have denticles very obsolete (visible only under higher magnification) thus the margin appears nearly smooth. On the other hand some specimens, particularly those from central Panama, have distinctly serrate outer margin of elytra. Specimens from central Panama ( Fig. 3 View Figs 1–3 ) (Cerro Campana, Cerro Caracoral and Cerro Jefe) also differ in slightly coarser punctation overall, pronotal spot always triangular and large, lateral sides of elytra yellow to 8 th row, slightly narrower pronotum and second antennomere slightly shorter. However, we consider these characters as intraspecific variability. Particularly, punctation and surface of elytra is quite often finer in west Panamanian cassidine populations than in central and eastern.
Etymology. The specific epithet arises from the plant family Orchidaceae and Latin “ voro ” = feed.
Egg. The egg of C. orcheidivora sp. nov. is light yellow, membranous, flat, semitransparent, 2.5 mm in length and 1.5 mm in width (n = 5) bearing no maternal adornments ( Figs 4, 8 View Figs 4–9 ).
Larval diagnosis. The larva of C. orchideivora sp. nov. can be distinguished from the four species described by GARCÍA- ROBLEDO et al. (2010) by the following character combination: dorsum medially ridged 2), lacking setae, prothorax with central and lateral areas shallowly rugose, prothorax carinate and venter shallowly rugose. One or two pairs of elongate, sclerotized plates, located medially on the prothorax ( Fig. 12 View Figs 10–15 ) distinguish C. orchideivora larvae from most if not all other Cephaloleia species. Degree of pigmentation in these structures is variable, darkening noticeably with age of first instar larva and reaching greatest density in the second instar. An additional pair of narrower, elongate sclerotized plates are present on the terminal tergite (fused abdominal segments 8 and 9) of most second-instar larvae. Larva of C. mauliki Uhmann, 1930 described by MAULIK (1932) lacks these plates and differs in stemmata arranged in one line of four and one line of two (two lines of three in C. orchideivora , Fig. 13 View Figs 10–15 ) and mandibles sharply tridentate (shallowly quadridentate in C. orchideivora , Figs 14–15 View Figs 10–15 ). Description of larva. Measurements. Upon emergence from the egg and before feeding first instar larva measured 2.3 by 1.3 mm (length by width) (n = 1); older first instar larvae 4.1 by 2.2 mm (n = 3), and second instar larvae 7.9 by 4.4 mm (n = 3). Body of greatest width at abdominal segment 3.
Color and size. Recently emerged first instar largely translucent and without pigmentation, while older first instar larvae have two dull, cream-colored, longitudinal stripes flanking a transparent region (gut) extending longitudinally along the midline. Margins are largely translucent, venter entirely transparent. Second instar differs from first by having one or two elongate, darkly-pigmented areas on the dorsal surface of the prothorax and one on the final abdominal segment. The body of both live and fresh ethanol preserved specimens have two conspicuous pink longitudinal stripes extending from mid-prothorax to abdominal segment 6, and groups of darker, greenish cells near spiracles, yellowing toward the margin. Body color in older specimens white bordered by transparent fringe, dorsum shallowly rugose, with an elevated medial ridge. Older ethanol preserved larvae turns to white or creamy color.
2) This character is, however, possibly an artifact caused by dehydration. We have observed many Cephaloleia larvae and they would have such dorsal ridge only in case of low humidity. It is also caused sometimes by quick dehydration in absolute alcohol.
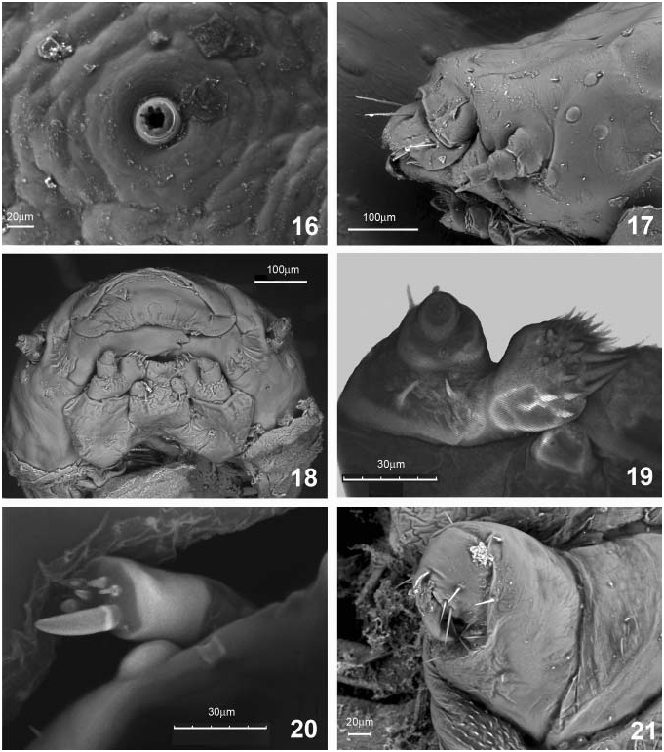
Dorsum. Basal prothorax elevated medially, sloping evenly toward anterior margin, surface shallowly rugose and medially bearing fields of sclerotized nonpigmented or pigmented cells, divided by diagonal anterior carina, sparsely placed microsetae; lateral margins rugose, terminating in a membranous fringe composed of chitinous rods of uniform breadth, surface populated by numerous minute spinules and a smaller number of longer setae (cf. MAULIK 1932, for a more detailed description of the fringe). Meso- and metathoracic segments elevated medially, shallowly rugose with sparse microsetae. Abdominal tergites 1–8 without carinae, shallowly rugose, bearing sparsely placed microsetae. Spiracles ( Fig. 16 View Figs 16–21 ) annular with crenulate peritreme, located equidistant between median and margin. Eighth abdominal spiracle located near margin between tergites 8–9. Separation of tergites 8 and 9 indistinct, margin rounded apically.
Venter. Surface of expanded segments smooth to shallowly tuberculate. Prothoracic spiracle opening to ventral surface between legs 1 and 2. Head ( Figs 17, 18 View Figs 16–21 ) surface smooth with 5 large and one small stemmata on each side; clypeus with surface smooth, basally with 4 long-stout setae and apically with margin densely fringed with short robust setae unequal in length; maxillary palp ( Fig. 19 View Figs 16–21 ) with two palpomeres, basal palpomere with 2 isolated setae, apical palpomere with one lateral seta, terminating with 11 short sensillae; maxilla robust, clavate, with two stout setae at base of palpomere and apically fringed with setae unequal in length and stoutness; labium densely setose; labial palp with one palpomere, terminated by approximately eight short sensillae; antenna ( Fig. 20 View Figs 16–21 ) with three antennomeres, apical antennomere bearing one long, stout conical sensillum, seven short sensillae and one short seta. All thoracic and abdominal segments shorter than wide, surface smooth, concave and joined to adjacent segments by transverse sutures. Femur of leg ( Fig. 21 View Figs 16–21 ) wider and shorter than subconical tibiotarsus, the latter bearing one stout claw and 10 setae at apex.
Comments on larval morphology. The larva of Cephaloleia orchideivora ( Figs 11, 12 View Figs 10–15 ) conforms closely to the basic “onisciform” Cephaloleiini ground plan described first by MAULIK (1932) and later for four species by GARCÍA- ROBLEDO et al. (2010). The larva cuts a transverse slit in one end of the egg and exits without consuming any part of the chorion. Larval thoracic and abdominal segments are well defined, extended, laterally flattened and terminated by a continuous membranous fringe (laterally-fused setae) which appears to form a hermetic seal maintained between the larva, both active and resting, and its glabrous substrate. The prognathous head is strongly retracted and attached to the ventral surface of the prothorax and thereby is concealed from direct view at all times and from all angles. Three to four stemmata are visible in dorsal view through the cuticle of the prothorax of the first instar larva ( Fig. 11 View Figs 10–15 ). Due to increased thickness and pigmentation of the epidermis and more lateral presentation on the head, stemmata are usually not visible in dorsal view of the second instar larva ( Fig. 12 View Figs 10–15 ). However, by manipulating or dissecting the head from its ventral position, a cluster of 6 stemmata (one most ventral far smaller) is readily apparent on each side of the vertex ( Fig. 13 View Figs 10–15 ) just posterior to the base of the antenna in both first and second instar larvae. Mandibles ( Figs 14, 15 View Figs 10–15 ) are shallowly quadri-dentate with a widened concave mesal surface.
Host plants. Orchideaceae: Elleanthus cf. robustus (Rchb. f.) Rchb. f., Elleanthus sp. , Epidendrum werklei Schltr. , Oerstedella exasperata (Rchb. f.) Hágsater, Oerstedella wallisii (Rchb. f.) Hágsater.
Ecological observations. The species lives in montane cloud forests from 800 to 1200 m a.s.l. Most specimens were collected in the windy pass called ‘Quijada del Diablo’ where the Rio Hornito trail begins in the La Fortuna Reserve in Chiriquí Province, western Panama. The locality is characterized by sparse and shrubby forest with stands of terrestrial orchids including the three known host plants, Elleanthus cf. robustus , Epidendrum werklei and Oerstedella exasperata . Other terrestrial orchids present include Epidendrum radicans Lindl. , Oncidium spp. , and Sobralia spp. , however, these were uniformly without feeding traces. The other localities where the new species was collected are Cerro Campana, Cerro Caracoral and Cerro Jefe, isolated mountains in central Panama where the vegetation is sparse, open and rather low. The label data from the specimen from Cerro Campana does not record any host plant or habitat.
Specimens from Cerro Jefe were feeding on a small Epidendrum -like epiphytic orchid growing about 4 m above ground level. While the plant was not flowering, it was clear from vegetative characters that it was distinct from either host plant species in Chiriquí. It had distinct warts on leaf sheaths and reddish scales on stem, characters found in the genus Oerstedella Rchb. f. The stem was similar to O. wallisii which grows abundantly on Cerro Jefe, however, we cannot assign it with certainty as our plant was without flowers. Nearby, on another occasion we found larvae on an epiphytic Elleanthus species.
Larval feeding damage is similar to that of the adult and is mostly present as narrow strips, often but not always paralleling leaf venation in smooth-leaved orchids ( Figs 4–7 View Figs 4–9 ) or as broader strips in species with pleated leaves ( Figs 8, 9 View Figs 4–9 ). Adult beetles are mostly hidden in the terminal leaf or between the terminal leaf and the base of last fully exposed leaf ( Fig. 7 View Figs 4–9 ). During the night adults also feed on the open leaf surface and sometimes the damage can be massive ( Fig. 6 View Figs 4–9 ). Adults lay eggs singly along the mid-vein of the terminal semi-closed leaf. The larva feeds on the top surface of the second or third newest leaves and shelters at the semi-closed leaf base ( Fig. 9 View Figs 4–9 ) in all host plant species. As the leaf expands and opens the larva moves to feed on the next newer leaf.
Distribution. Panama (Bocas del Toro, Chiriquí, Coclé and Panamá provinces).



| V |
Royal British Columbia Museum - Herbarium |
| USNM |
Smithsonian Institution, National Museum of Natural History |
| CRH |
Centre de Recherche en Hydrobiologie |
| UPPC |
University of the Philippines |
| NMPC |
National Museum Prague |
| T |
Tavera, Department of Geology and Geophysics |
| R |
Departamento de Geologia, Universidad de Chile |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |