
Korotkevitschia pelagica ( Korotkevitsch, 1961 )
|
publication ID |
https://doi.org/ 10.5281/zenodo.170850 |
|
DOI |
https://doi.org/10.5281/zenodo.5689964 |
|
persistent identifier |
https://treatment.plazi.org/id/03FD9E73-FFCB-1035-FEF3-141024D811C0 |
|
treatment provided by |
Plazi |
|
scientific name |
Korotkevitschia pelagica ( Korotkevitsch, 1961 ) |
| status |
|
Korotkevitschia pelagica ( Korotkevitsch, 1961) View in CoL ( Figs 1–20 View FIGURE 1 View FIGURES 2 – 7 View FIGURE 8 View FIGURES 9 – 14 View FIGURES 15 – 20 )
Cratenemertes pelagicus Korotkevitsch, 1961
Type specimen: Holotype: Zoological Institute of Russian Academy of Science, St.Petersburg, Russia, no 278, 28 slides with 778 oblique and transverse sections of complete immature male.
Type location: Indian Ocean, 61°33' S, 20°00' E, from a depth of 0–200 m collected February 25,1957 by K.V. Beklemishev and V.S. Korotkevitsch.
Description
Epidermis and dermis: The epidermis is 60–85 m thick in intestinal region and 80–100 m in cerebral region. It contains numerous weakly staining glands. The epidermal cells have a basalcup zone. The dermis in the intestinal region is 26–38 m thick but it reaches up to 46 m in the cerebral region. The basal part of the dermis is deeply stained.
Body wall musculature: The outer circular muscle layer is mostly 20–27 m thick. The inner longitudinal musculature varies from 37 m to 110 m thick. It is divided into wedges with typical "honeycomb" structure ( Crandall & Gibson, 1998). The pathways between adjacent wedges are narrow and occasionally contain 1–3 dorsoventral fibres. The precerebral septum is of the closed type. Welldeveloped cephalic retractor muscles derive from longitudinal body musculature and the precerebral septum ( Figs 1 View FIGURE 1 , 2 View FIGURES 2 – 7 ).
There is a thin layer of nonfasciated diagonal muscles between the circular and longitudinal layers ( Fig. 3 View FIGURES 2 – 7 ). It is clearly visible in the cerebral and intestinal regions but is completely absent in the rectal region. Diagonal fibres appear finer than the circular and longitudinal fibres and comprise two layers one fibre thick each. Fibres of the diagonal layers crisscross at an angle of 105–110 degrees but this condition may be the result of strong contraction of the specimen. Diagonal fibres in each layer are closely situated but some may be spaced about 2–6 fibres diameters apart.
There is a delicate layer of longitudinal fibres around the stomach wall ( Fig. 4 View FIGURES 2 – 7 ). Groups of radial fibres derived from stomach longitudinal muscles pass between the rear of the brain and the posterior part of the cerebral organ ( Fig. 5 View FIGURES 2 – 7 ). Dorsoventral musculature is weakly developed. Dorsoventral bundles are 3–6 fibres thick and pass lateral to the intestinal diverticula and the nerve cords. The thicker bundles lie between intestinal diverticula.
Parenchyma: Parenchyma is weakly developed around the rhynchocoel wall and intestine. The parenchymal layer around the intestine is mostly 2–4 m thick, but occasionally may reach up to 20 m thick.
Rhynchodaeum and rhynchocoel: Mouth and rhynchodaeum open separately ( Figs 1 View FIGURE 1 , 6 View FIGURES 2 – 7 ). The rhynchodaeum is lined by a ciliated epithelium 8–11 m thick. Anteriorly, longitudinal retractor muscles border the rhynchodaeum wall. The rhynchodaeal sphincter of circular muscle is up to 62 m thick. Immediately posterior to the sphincter the rhynchodaeum forms a large chamber lined by a glandular epithelium.
The rhynchocoel ends in the rectal region (section 715). The rhynchocoel wall musculature is of the interwoven type and 6–25 m thick ( Fig. 7 View FIGURES 2 – 7 ). The rhynchocoel endothelium is mostly very thin but can reach up to 8– 10 m.
Proboscis: The anterior chamber of proboscis is about 0.9–1.1 mm in diameter. Its wall is composed of a glandular epithelium 260–310 m tall with wide papillae, a layer of outer circular muscle 12–18 m across, a longitudinal muscle layer 85–100 m across, an inner circular layer only one fibre thick and flattened endothelium. The neural sheath, 6–9 m thick, divides the longitudinal muscle layer into inner and outer portions. Thirteen proboscis nerves pass through both anterior and posterior proboscis chambers. In its posterior third, the anterior chamber narrows to 0.6–0.8 mm in diameter and has more elongate epithelial papillae and a thicker neural sheath 12–16 m across. There is no bulb region between anterior and posterior chambers. The stylet pouch portion of the proboscis is not significantly differentiated from the anterior chamber ( Fig. 8 View FIGURE 8 ). It is 440–475 m in diameter, with two stylet pouches. The pouches are up to 170 m in maximum diameter, each containing 8–9 typical stylets 70–78 m long. There is a narrow opening in the posterior part of the pouches. Central stylet, basis and diaphragm are absent. Immediately anterior to the stylet portion a tubelike structure is formed in the proboscis lumen. This structure consists of cellular components in an extracellular substance and connects by filaments of the extracellular substance with the proboscis papillae. The anterior part of this structure is 80 m in diameter and opens into the lumen of the anterior proboscis chamber. Posteriorly, the "tube" widens up to 160 m in diameter and joins to glandular epithelium just behind the stylet pouches. It is probable that this structure is a tubelike clot of extracellular gel with cellular and tissue fragments ( Figs 9, 10 View FIGURES 9 – 14 ).
Immediately behind the stylet pouches the elongated dark staining acidophilous glands lie under the glandular epithelium and open into the proboscis lumen ( Fig. 10 View FIGURES 9 – 14 ). The position and texture of these glands are strongly resemble the stylet basis glands in other monostiliferous nemerteans.
The posterior portion of the proboscis is 250–350 m in diameter. Its wall is composed of a glandular epithelium 37–52 m thick, 13 bands of longitudinal muscle separated by 13 small nerves, a very thin layer of inner circular muscles and a flattened endothelial layer.
Frontal organ: The frontal organ opens slightly subterminally; it comprises a hemispherical chamber about 100 m in diameter ( Fig. 2 View FIGURES 2 – 7 ).
Precerebral glands: The "cephalic gland" complex is not well differentiated and represented by weakly staining basophilic frontal and cephalic glands. The frontal glands are associated with the frontal organ and seem to be situated lateral and dorsal to the anterior part of the rhynchodaeum. Diffuse basophilic cephalic glands are interspersed among retractor muscles in the precerebral region. There are a few granulated submuscular glands situated ventrolaterally in the anterior part of the precerebral region. The elongate, dark staining granulated glands anteriorly surround the rhynchodaeum and open subterminally around the rhynchopore and mouth ( Figs 6 View FIGURES 2 – 7 , 11 View FIGURES 9 – 14 ). Similar circumrhynchodaeal gland cells have been described in the cratenemertid Achoronemertes scoresbyi ( Crandall & Gibson, 1998) .
Cephalic grooves: Korotkevitsch (1961, p. 1417) reported that "… at the level of the brain, there is a shallow cutaneous ridge at the bottom of which the cerebral organ canals open". The shallow cephalic grooves lack secondary grooves found in most species of Nipponnemertes .
Nervous system: The well developed brain is enclosed by a thin outer neurilemma, but contains neither an inner neurilemma nor neurochord cells. Dorsal and ventral ganglia are of nearly equal size and clearly separated at the rear of brain. Dorsal and ventral commissures are respectively 73 m and 94 m thick. The lateral nerve cords continue posteriorly directly from the ventral ganglia. A very thin band of connective tissue adjoins the medial margin of the fibre core of the nerve cord. 50–60 myofibrils lie in the vacuolate space occupying 2/3–3/4 of the length of the band ( Fig. 12 View FIGURES 9 – 14 ). As the nerve cord extends posteriorly Jshaped or Ushaped arrangement of the myofibrils becomes clearly visible. In the posterior quarter of the body ventral myofibrils enter a ganglion cell layer. The posterior commissure of the nerve cords (section 766) is above the rectum on the level of the vascular anastomosis and way behind the end of rhynchocoel.
A few nerves lead forwards from the front of brain. The proboscis nerves originate from a pair of nerve trunks arising from the medial face of the brain below the dorsal commissure but above the ventral commissure.
The middorsal nerve is small and situated between the dermis and body circular musculature.
Cerebral organs: The cerebral organs are smaller than in species of Nipponnemertes . They open close behind the anterior brain border at about the 9–10 and 2–3 o'clock positions. The anterior elongate portion of the cerebral organ lies alongside the outer brain margin ( Figs 5 View FIGURES 2 – 7 , 13 View FIGURES 9 – 14 ). The posterior rounded portion of the cerebral organ contacts the rear of the brain ( Fig. 13 View FIGURES 9 – 14 ). The canal of the cerebral organ is bifurcated and leads to a saclike branch and a sensory canal ( Fig. 5 View FIGURES 2 – 7 ). The saclike branch continues posteriorly for about half the organ's length, while the sensory canal terminates at the rear of the organ ( Fig. 13 View FIGURES 9 – 14 ).
Blood vascular system: The blood system has a typical monostiliferous arrangement consisting of a cephalic loop and three postcerebral vessels. The cephalic loop is simple. The middorsal vessel arises just behind the ventral brain commissure but its origin is obscured. Behind the ventral commissure the middorsal vessel penetrates the rhynchocoel wall to form a short (total length approximately 140 m) vascular plug 20 m in diameter. There is a coiling of both the dorsal and lateral vessels in the sections 717–720. The posterior vascular anastomosis occurs above the rectum (sections 766–767).
Excretory system: The excretory tubules extend from just behind the brain to the midbody region. They consist of typical collecting ducts 24–30 m in diameter, running above the lateral nerve cords. A single slender efferent canal, 16–21 m wide, penetrates through all body wall layers in lateroventral direction ( Fig. 14 View FIGURES 9 – 14 ) and opens on each side of the body via a deep epidermal invagination up to 88 m in diameter (on the left at section 420 and on the right at section 353). The dermis is very thin under this epidermal chamber. The exterior openings are at about the 4.30 and 7.30 o'clock positions.
Alimentary canal: The mouth opens just under the rhynchopore ( Figs 1 View FIGURE 1 , 15 View FIGURES 15 – 20 ). There is no distinct esophagus in the anterior portion of the foregut. The stomach is long with a deeply folded epithelium 40–60 m thick. The anterior portion of the stomach is partly everted through the mouth. The large protrusions described for Achoronemertes scoresbyi ( Crandall & Gibson, 1998) are not found in K. pelagica , but its stomach has a few small protrusions in the precerebral region ( Fig. 4 View FIGURES 2 – 7 ). The pylorus is approximately 1.2 mm long, very flattened, with a ciliated epithelium 7–9 m thick ( Fig. 16 View FIGURES 15 – 20 ). The pyloric epithelium contains numerous granular glands. The intestinal caecum does not possess anterior pouches but is provided with three pairs of simple lateral pouches. The lateral intestinal pouches are large and possess shallow lobes ( Fig. 17 View FIGURES 15 – 20 ). There is a very long rectum, almost 10 % of the total body length, beginning in section 701–702. In section 695 a gut, about 420 m wide, has very small lateral pouch. Sections 696–700 are absent but in sections 701–703 the gut has small lateral pouch too. In section 704–706 the gut tube, 140–170 m wide, has one left and one ventral pouches (each about 160 m wide). Beginning in section 707 the gut has a typical rectal construction narrow tube without pouches ( Fig. 18 View FIGURES 15 – 20 ). Under and behind the caudal nerve commissure (sections 766–770) a sphincter of circular muscle 7–13 m thick surrounds the rectum ( Fig. 19 View FIGURES 15 – 20 ). Unfortunately, 30 sections of the rectal region of the body are missing and it is impossible to determine whether additional sphincters are present.
Reproductive system: The type specimen is an immature male. The narrow and elongated testes lie in the posterior quarter of the intestinal region, passing between the lateral nerve cords and the intestinal wall ( Fig. 20 View FIGURES 15 – 20 ) and opening via dorsolateral gonoducts. There is no gonadal muscle meshwork as found in many cratenemertid species ( Crandall, 1993), although scattered longitudinal muscle fibres are present.
Systematic discussion: A comparison between the present description and that originally given by Korotkevitsch (1961) reveals a number of discrepancies which require discussion. Neither the intestinal caecum nor cephalic glands were found by the original author. Although the intestinal caecum of K. pelagica is well developed, it is hardly distinguishable from the intestine on the deformed oblique sections. The precerebral gland complex of K. pelagica includes four types of the glands (frontal, diffuse cephalic, submuscular and circumrhynchodeal glands) but Korotkevitsch may have overlooked unstained frontal and cephalic glands. The excretory system extends from just behind the brain to the middle portion of the body, contrasting with Korotkevitsch's statement that nephridia are absent. I reexamined the transverse section (number 307, Fig. 17 View FIGURES 15 – 20 ), figured in the original description ( Korotkevitsch, 1961: fig. 1 B), and found that the structures Korotkevitsch labelled as gonads are, in fact, nephridia.
Hitherto only two species of the pelagic cratenemertids have been described, Achoronemertes scoresbyi and Korotkevitschia pelagica . In many respects K. pelagica and A. scoresbyi are very similar. Both species possess a suite of characters that are distinctly unusual among hoplonemerteans. These features include: modification of the proboscis bulb region, absence of the central armature and bulb duct, a remarkably long rectum, the extraordinary length of the excretory system with nephridiopores opening far back in the intestinal region. Norenburg (1986) noted that the structure of nephridial systems in Monostilifera is linked to habitat and therefore should be used as a diagnostic feature with caution. The apparent modification of the proboscis bulb region either represents a result of the reduction ( Crandall, 2001) or exhibits the initial stage of the formation of the monostiliferous armature ( Chernyshev, 2003).
Several other anatomical characteristics are shared by A. scoresbyi and K. pelagica , including pelagic habitat, the absence of eyes, poorly developed parenchyma, nonfasciated diagonal muscles, simple cephalic grooves, presence of frontal, cephalic, submuscular and circumrhynchodaeal glands. Three significant taxonomic characters, found in Korotkevitschia , distinguish it from Achoronemertes : the mouth and rhynchodaeum open separately, the intestinal caecum does not possess anterior pouches, stomach longitudinal muscles are present. Relationships between mouth and proboscis openings have long been used at the generic level. However a strongly contracted cephalic region or some eversion of an atrium can give the appearance of separate openings ( Crandall, 1993). Additional differences that can be used to separate both taxa are summarized in Table 1 View TABLE 1 .
One of the unusual features of K. pelagica is that the testes lie in the posterior quarter of the intestinal region. Since neither specimen of A. scoresbyi contained any evidence of gonads, no comments can be made regarding the reproductive system of other epipelagic cratenemertids. In the pelagic polystiliferous nemerteans the testes are limited to the cephalic or foregut region.
Crandall's phylogenetic analyses show that Korotkevitschia , Achoronemertes and a putative new genus of the epipelagic nemertean ("AtlanticH") form a strongly supported clade within the family Cratenemertidae . Two unique characters support this clade: paired nephridial ducts in the midbody region and a very long hindgut. Both Korotkevitschia and Achoronemertes differ from "AtlanticH" by the absence of a proboscis bulb duct. Crandall (2001, p. 104) considered that phylogenetic analysis "would support, at least on a provisional basis, treatment of the pelagic group, 'PacificP', and the group of remaining genera as subfamily grouping". Recently Thollesson and Norenburg (2003) have indicated that the Cratenemertidae is the sister group to the remaining monostiliferans and renamed the cratenemertid clade as Cratenemertea. However, they provided Cratenemertea neither a formal generic diagnosis nor even a single synapomorphy. Chernyshev (2003) regards Cratenemertea as an order and establishes a new family Korotkevitschiidae for epipelagic genera. I extend the diagnosis of the family as follows:
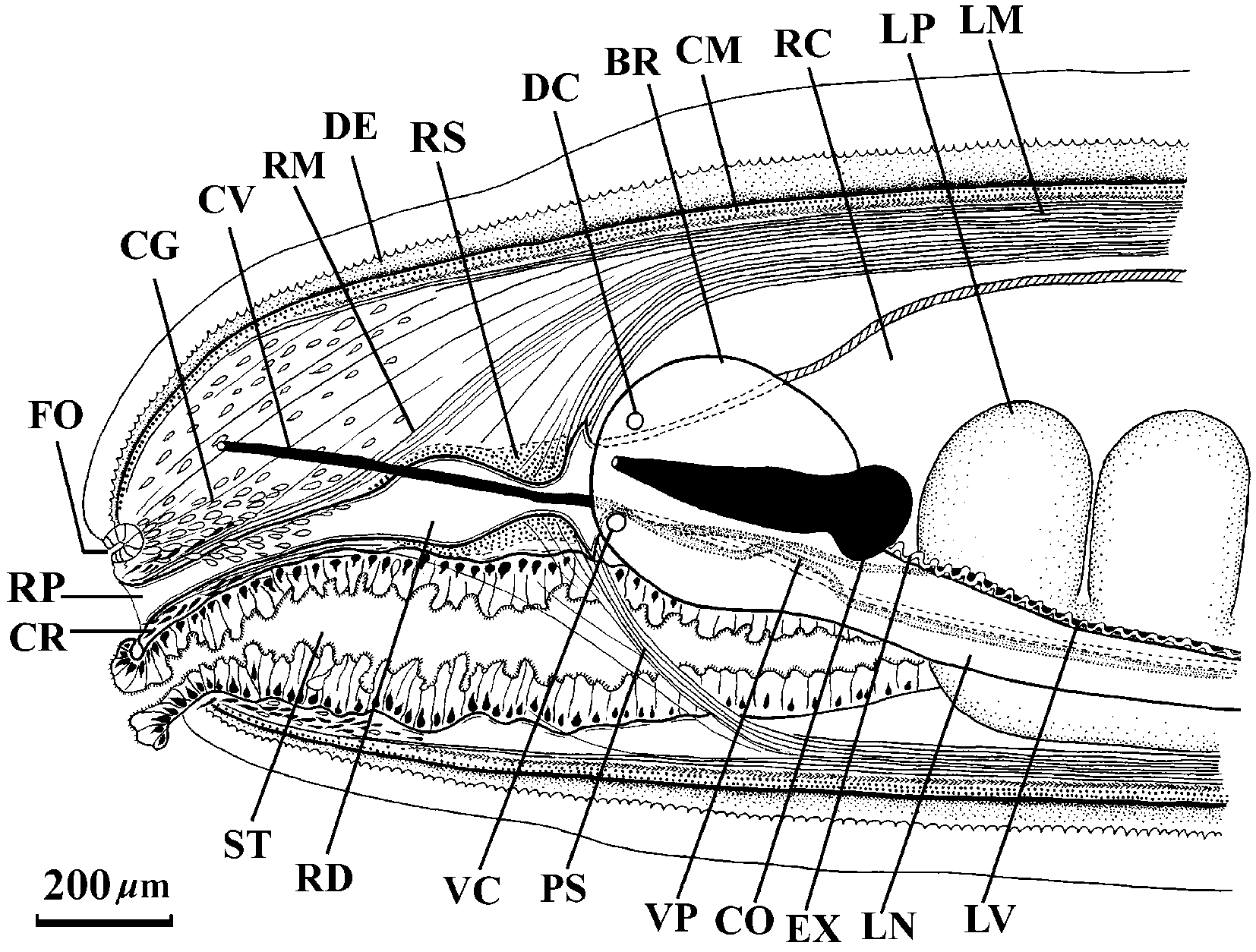
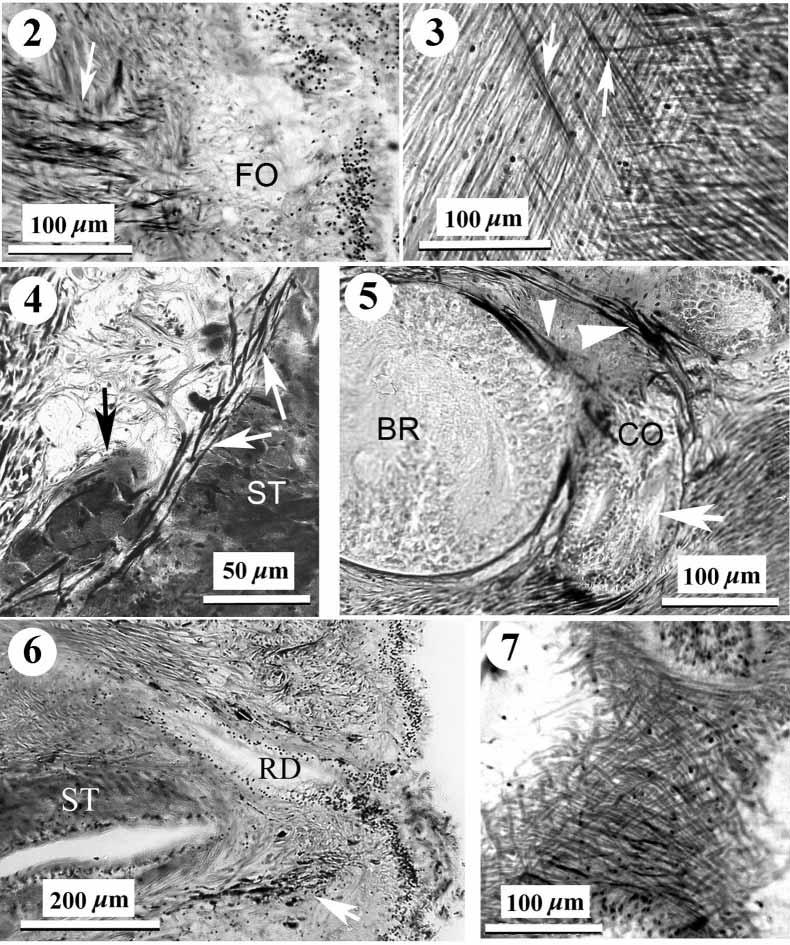
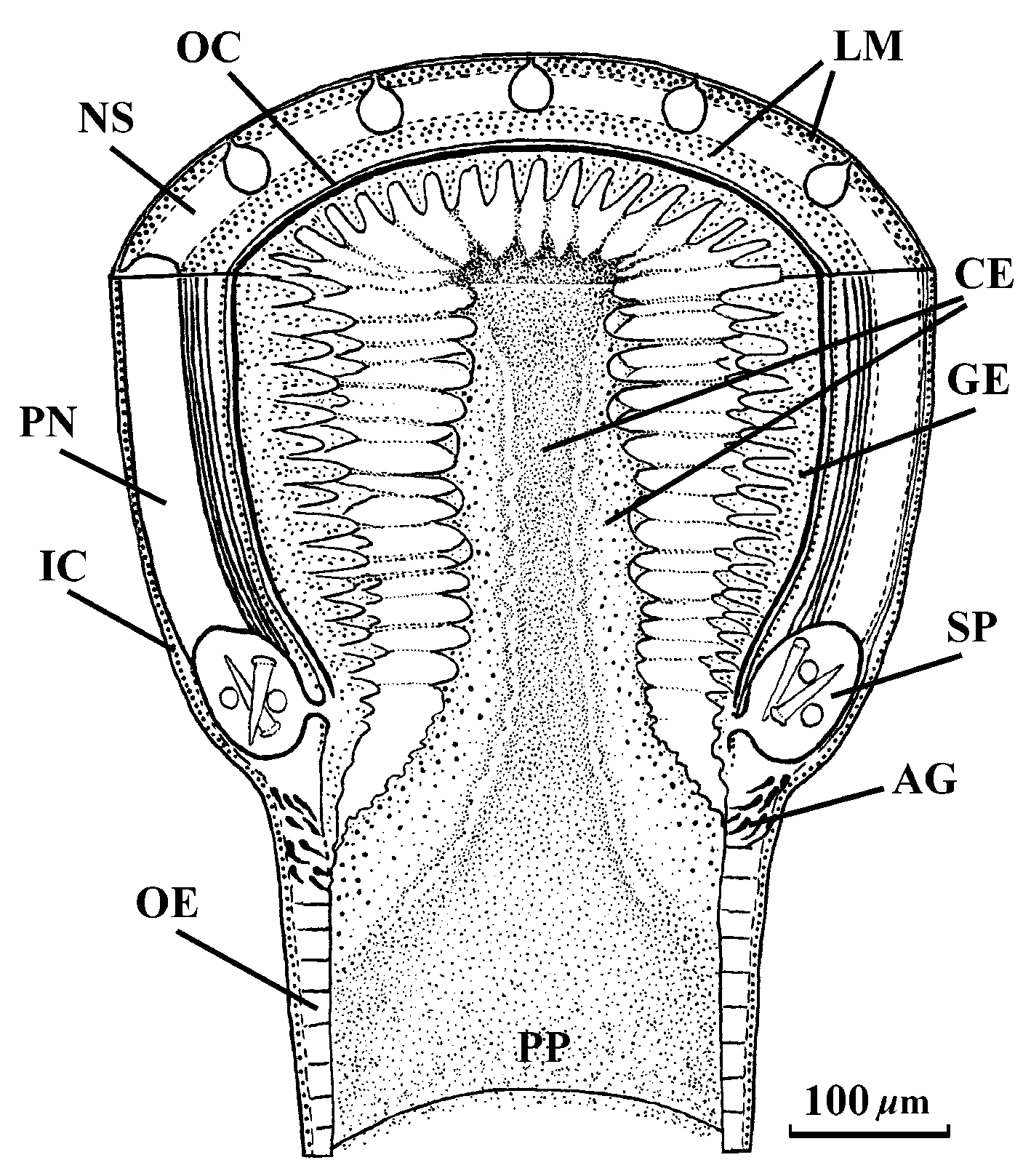
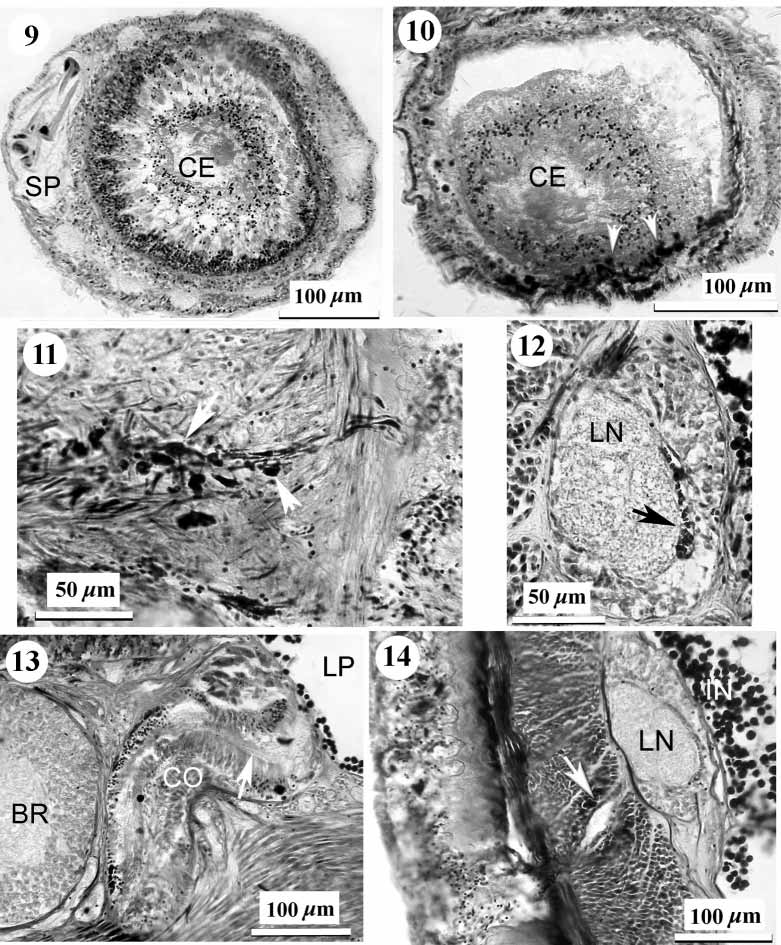
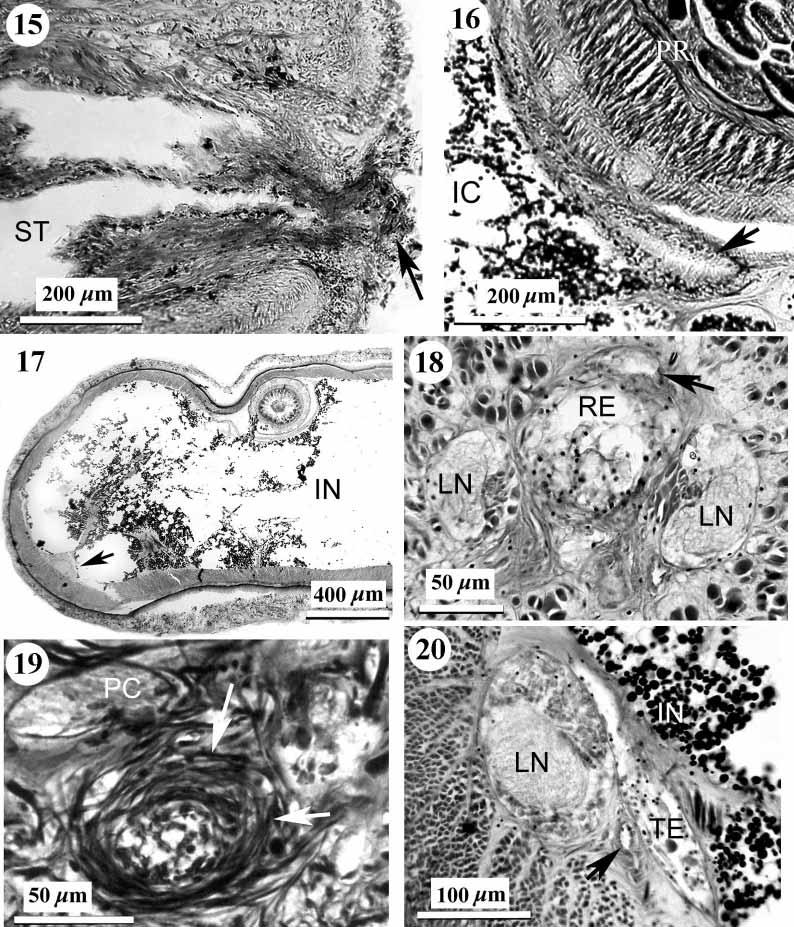
TABLE 1. Some of the principle characters distinguishing Korotkevitschia pelagica and Achoronemertes scoresbyi. Data taken from Wheeler (1934), Korotkevitsch (1961), Crandall & Gibson (1998) and the present paper.
| Features | Korotkevitschia pelagica | Achoronemertes scoresbyi |
|---|---|---|
| Body length (mm) | 9 | 5.5 |
| Body color in life | light orangepink | orangeyellow to orangered |
| Parenchyma in the intestinal region | poorly developed | virtually nonexistant |
| Rhynchostomemouth opening | separate | atrium |
| Number of proboscis nerves | 13 | 12 |
| Number of accessory stylets in each pouches | 8–9 | 2–4 |
| Oesophagus | absent (?) | present |
| Anterior pouches of intestinal caecum | absent | present |
| Length of rectum | 1/10 of total body length | 1/6 of total body length |
| Sphincter of circular muscle around rectum | 7–13 µm thick | very thin (onetwo fibres thick) |
| Stomach musculature | thin layer of longitudinal fibres | absent |
| Nephropore position (percent of body length) | 45–54 % | 55–60% |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Korotkevitschia pelagica ( Korotkevitsch, 1961 )
| Chernyshev, Alexei V. 2005 |
Cratenemertes pelagicus
| Korotkevitsch 1961 |