
Monstrillopsis hastata, Suárez-Morales & Mckinnon, 2014
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3779.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:096F0F73-2CA0-4759-9DF6-C8B4654EDB46 |
|
DOI |
https://doi.org/10.5281/zenodo.5060971 |
|
persistent identifier |
https://treatment.plazi.org/id/03FC87D5-713B-FFA0-34E7-FCDE3B3820EE |
|
treatment provided by |
Felipe |
|
scientific name |
Monstrillopsis hastata |
| status |
sp. nov. |
Monstrillopsis hastata sp. nov.
( Figs 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 )
Material examined: Holotype: adult male from Western Port Bay (Station G 2VA of Kimmerer & McKinnon 1985), Victoria, Australia (38°35.344’S 144°59.687’E), partially dissected, slide-mounted in glycerine, sealed with Entellan®. Date of collection: 15 May 1983. Slide deposited in MTQ, Australia (cat. MTQW34268 ). GoogleMaps
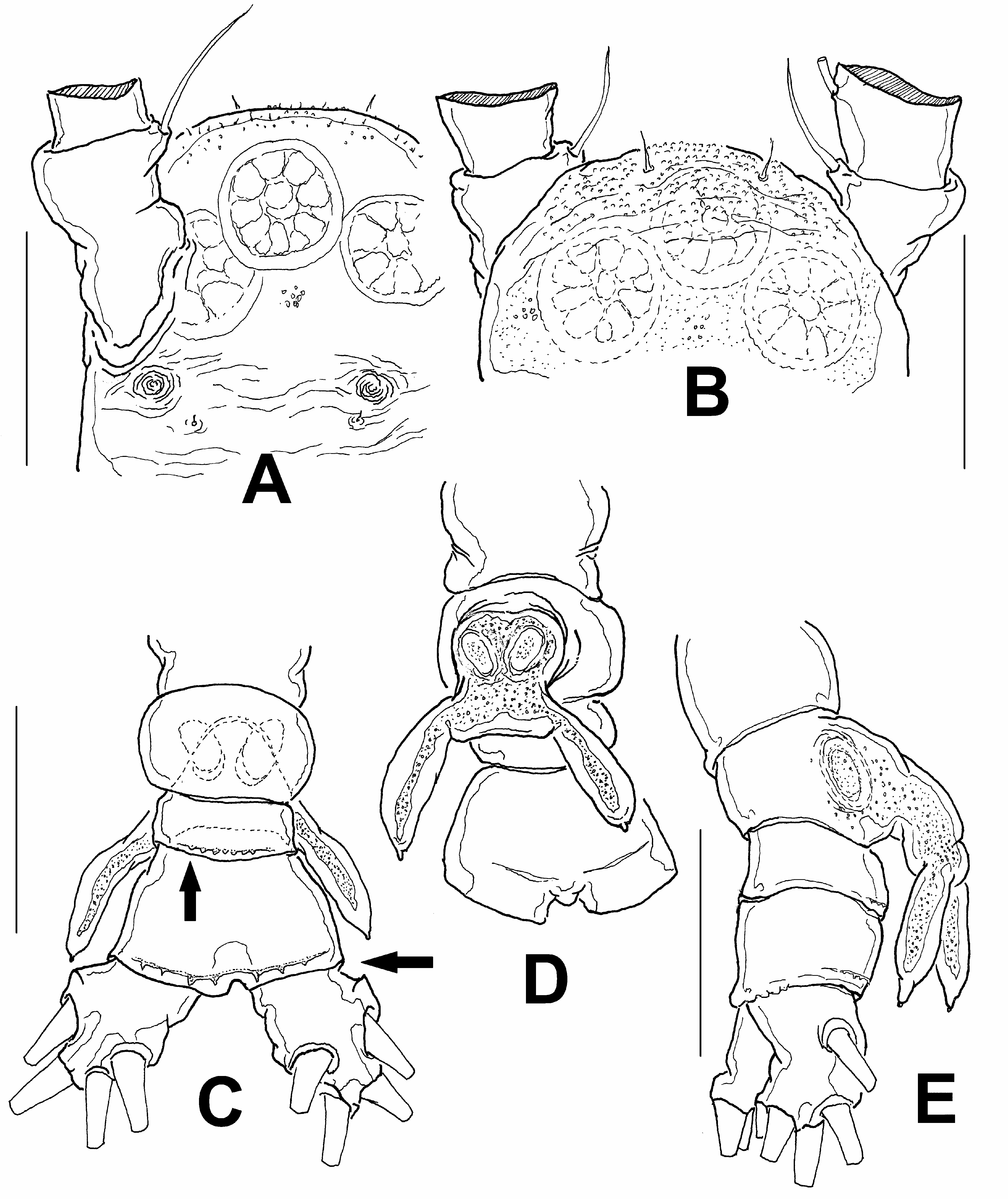
Description. Male: Total body length of adult male holotype: 0.81 mm. Cephalothorax 0.42 mm long, representing 51% of total body length. Antennule 0.38 mm long, representing 47% of total body length and 85% of cephalothorax length ( Fig. 1A, B View FIGURE 1 ). Oral papilla small, located anteriorly, about 27% of way back along ventral surface of cephalothorax ( Fig. 1A View FIGURE 1 ). Pair of relatively large ocelli present, pigment cups moderately developed, separated by less than half eye diameter, weakly pigmented; ventral cup and lateral cups equally sized. Forehead widely rounded, with two large sensilla and field of small cuticular papillae extending to most of cephalic area, including ventral and dorsal surfaces ( Fig. 2A, B View FIGURE 2 ). Ventral surface of cephalic area bearing two pairs of nipple-like cuticular processes between antennule bases and oral papilla, one pair being small ( Fig. 2A View FIGURE 2 ). Light pattern of cuticular striation present surrounding these structures, transverse striae reaching also middle ventral surface between nipple-like processes ( Fig. 2A View FIGURE 2 ).
As usual in male monstrilloids, antennules five-segmented, geniculate ( Fig. 1C View FIGURE 1 ). In terms of pattern described by Grygier and Ohtsuka (1995), element 1 present on first segment; elements 2d 1, 2d 2, 2v 1, 2v 2, 2v 3, and IId present on second segment. Third segment with elements 3, IIId, and IIIv, the latter being unusually stiff and slender ( Fig. 1C View FIGURE 1 ). Fourth segment bearing elements 4d 1,2 and 4v 1-3 as well as IVd. Inner middle margin of segment with pair of subtriangular protuberances near insertion of element 4v ( Fig. 1D View FIGURE 1 ). Terminal segment with elements 1–6, unbranched elements A–D (on posterior margin), and short medial aesthetasc ( Fig. 1D View FIGURE 1 ). As usual in male Monstrillopsis , terminal antennular segment modified, with inner rounded expansion and elongate, sabre-like distal half representing 58% of segment length; inner expansion ornamented with four spinules (arrowed in Fig. 1C View FIGURE 1 ).
First pedigerous somite incorporated into cephalothorax; this and succeeding three free pedigerous somites each bearing pair of biramous swimming legs. Pedigerous somites 2–4 together accounting for 33% of total body length in dorsal view. Intercoxal sclerites of legs 1–4 sub-rectangular, without ornamentation on surface or along distal margin. Basis of legs articulating with rectangular coxa along diagonal line. Basis with thin, simple lateral seta on legs 1, 2, and 4; on leg 3, this seta thicker, lightly setulate and four times longer than on other legs. Endopodites and exopodites of swimming legs 1–4 triarticulate ( Fig. 3A–C View FIGURE 3 ). Ramus setae all lightly and biserially plumose except for spiniform outer setae on exopodal segments 1 and 3, and inner seta of first exopodal segment, these all being short and slender. Also, outer apical exopodal seta of swimming legs 1–4 with outer margin smooth, inner margin lightly setose.
| Armature formula of swimming legs: | |||
|---|---|---|---|
| basis | endopodite | exopodite | |
| leg 1 | 1-0 | 0-1;0-1;1,2,2 | I-1;0-1;I,2,2 |
| legs 2–4 | 1-0 | 0-1;0-1;1,2,2 | I-1;0-1;I,1,2,2 |
Fifth legs absent. Urosome consisting of four somites: fifth pedigerous somite, genital somite with genital apparatus, one free postgenital somite, and large anal somite possibly incorporating fused preanal somite ( Fig. 2E View FIGURE 2 ). Ventral surface of genital somite forming enlarged base of cylindrical shaft with distal genital lappets. Lappets represented by divergent pair of posteriorly directed cylindrical processes, each tapering distally into nipple-like process and reaching distal margin of anal somite ( Fig. 2C–E View FIGURE 2 ). Anal+preanal somite large, representing 37% of urosome, ornamented with posterior hyaline fringe of short spines on dorsal surface; similar ornamentation also present on dorsal surface of succeeding free postgenital somite (arrows in Fig. 2C View FIGURE 2 ). Anal+preanal somite with irregular posterior margin in dorsal view ( Fig. 2E View FIGURE 2 ). Caudal rami subrectangular, symmetrical, divergent, approximately 1.3 times longer than wide, each ramus bearing four setae ( Fig. 2C,E View FIGURE 2 ).
Female: unknown.
Type locality. Western Port Bay, Victoria, Australia (38°35.344’S 144°59.687’E).
Etymology. An adjective derived from the Latin noun hasta, meaning spear, the specific epithet makes reference to the unusually stiff, spear-like setal element IIIv of the fourth antennular segment.
Diagnosis. Third antennular segment bearing stiff seta IIIv, fourth segment with two nipple-like protuberances. Terminal antennular segment modified, with inner rounded expansion and elongate distal half, latter sabre-like and representing 58% of length of segment; inner expansion of this segment ornamented with four spinules. One free postgenital somite; preanal and anal somites apparently fused with no trace of intersegmental suture. Genital complex of type II (defined below), with mammiliform distal tips of lappets. Spinous hyaline fringe posteriorly on dorsal surface of fused preanal+anal somites and free postgenital somite.
Remarks. This male specimen is easily assignable to the genus Monstrillopsis by its possession of a distinctively modified fifth antennular segment, with an inner rounded protuberance and an attenuated, sabre-like distal half (see Huys & Boxshall 1991; Suárez-Morales et al. 2006). The position of the oral papilla far anteriorly on the cephalothorax, the fully developed eyes, and the presence of four setal elements on the caudal rami are also regarded as diagnostic of this genus ( Sars 1921; Suárez-Morales et al. 2006). This specimen has a reduced number of free postgenital somites compared to its congeners, one instead of two: as mentioned in the descriptive section, the preanal and anal somites appear to be completely fused, without traces of an intersomite suture. This condition is usually found in males of Cymbasoma and has not been known to occur in Monstrillopsis except for incomplete fusion marked by a ventral suture and lateral constrictions in M. chathamensis Suárez-Morales & Morales- Ramírez, 2009 ( Suárez-Morales & Morales-Ramírez 2009) and also in Monstrillopsis boonwurrungorum , described in the present work. In other species of the genus in which the division of the preanal and anal somites is fully expressed, as in Mon. sarsi ( Isaac, 1974) , Mon. chilensis Suárez-Morales, Ramírez & Derisio, 2008 (Suárez- Morales et al. 2008), and Mon. cahuitae Suárez-Morales & Carrillo, 2013 ( Suárez-Morales et al. 2013), the anal somite is short.
Among the males of Monstrilla and Monstrillopsis there are two main types of genital complex, one of which is represented by a long, cylindrical apparatus with paired distal globose processes (type I). This kind of genital structure has been recorded mainly in species of Monstrilla ( Suárez-Morales et al. 2013) , but also in Monstrillopsis fosshageni ( Suárez-Morales & Dias 2001) and its congener Mon. cahuitae ( Suárez-Morales et al. 2013). The type II apparatus is similar to that present in our specimen, with a short shaft and relatively long, basally separated lappets. This kind of genital complex was earlier reported in two other species of Monstrillopsis, Mon. chilensis Suárez-Morales, Bello-Smith and Palma, 2006 from the Beagle Channel ( Suárez-Morales et al. 2008) and Mon. chathamensis from the Eastern Tropical Pacific ( Suárez-Morales & Morales-Ramírez 2009). The present new species has a type II genital complex but differs from its above-mentioned congeners in having mammiliform, not rounded, lappet tips. The new species also differs in characters of the antennule structure; it has a longer terminal sabre-like process of the fifth segment than in Mon. chilensis and Mon. chathamensis and it also has a set of 4–5 spinules on the proximal rounded process of the segment, whereas this process is naked in Mon. chathamensis and in two of the four Australian species described herein. This process is ornamented with a patch of minute spinules in Mon. chilensis ( Suárez-Morales et al. 2008). A set of spiniform elements is present in another Australian species described herein, Mon. nanus sp. nov., but the genital lappets have a different structure.
The present new species can be distinguished from all other species of Monstrillopsis on the basis of: 1) the stiff seta IIIv, which is usually flexible, not so rigid or spiniform as here, in both Monstrillopsis and other known monstrilloids; 2) the presence of two nipple-like protuberances on the fourth antennular segment ( Fig. 1D View FIGURE 1 ), a feature not hitherto reported in any monstrilloid; 3) the mammiliform distal tips of the genital lappets; and 4) the spinous hyaline fringes on the dorsal surface of the free postgenital (preanal) and anal somites. A similar fringe was described in both Mon. chathamensis and Mon. chilensis, but only on the anal somite and at a more posterior position ( Suárez-Morales & Morales-Ramírez 2009; Suárez-Morales et al. 2008).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |