
Monstrillopsis nanus, Suárez-Morales & Mckinnon, 2014
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3779.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:096F0F73-2CA0-4759-9DF6-C8B4654EDB46 |
|
DOI |
https://doi.org/10.5281/zenodo.4910346 |
|
persistent identifier |
https://treatment.plazi.org/id/03FC87D5-7133-FFA9-34E7-FADB3C862005 |
|
treatment provided by |
Felipe |
|
scientific name |
Monstrillopsis nanus |
| status |
sp. nov. |
Monstrillopsis nanus sp. nov.
( Figs 6–8 View FIGURE 6 View FIGURE 7 View FIGURE 8 )
Material examined: Holotype: adult male from Giralia Bay , Exmouth Gulf , Western Australia (22°28’S; 114°20.75’E; McKinnon & Ayukai 1996), undissected, ethanol-preserved, slide mounted in glycerine, sealed with Entellan ®. Date of collection: 25 September, 1994. Slide deposited in WAMC, Australia (cat. WAMC55071 ). GoogleMaps
Description. Male: Total body length of adult holotype 0.50 mm. Cephalothorax 0.25 mm long, representing 50% of total body length. Antennule 0.23 mm, relatively long, 1.15 times as long as cephalothorax and representing 48% of total body length ( Fig. 6A, B View FIGURE 6 ). Oral papilla small, located anteriorly, about 35% of way back along ventral surface of cephalothorax ( Fig. 6A View FIGURE 6 ). Pair of relatively small ocelli present, pigment cups moderately developed, separated by less than half eye diameter, weakly pigmented on outer rim; ventral cup slightly larger than lateral cups. Forehead nubbly, with medial rounded protrusion ( Fig. 7C, D View FIGURE 7 ). Ventral surface of cephalic area with single pair of nipple-like cuticular processes and adjacent field of transverse cuticular striations between antennule bases and oral papilla ( Fig. 7D View FIGURE 7 ).
As usual in male monstrilloids, antennules five-segmented, geniculate ( Fig. 7A View FIGURE 7 ). In terms of pattern described by Grygier and Ohtsuka (1995), element 1 present on first segment; elements 2d 1, 2d 2, 2v 1, 2v 2, 2v 3, and IId present on second segment. Third segment with elements 3, IIId, and IIIv. Segment four bearing elements 4d 1,2 and 4v 1,2 ( Fig. 7B View FIGURE 7 ) (element 4v 3 not observed), as well as elements IVd and IVv. A spiniform setal element observed between elements IVd and IVv, not identifiable by position following Grygier and Ohtsuka (1995),thus provisionally designated as element 4v 1 ( Fig. 7A View FIGURE 7 ). Following Huys et al. ’s (2007) nomenclature, armature of terminal segment including elements 1, 2, 5, and 6, plus unbranched elements A–D on posterior margin. Terminal antennular segment modified, inner rounded expansion ornamented with row of five short, spiniform elements; elongate distal half sabre-like, curved distally, comprising 50% of segment length.
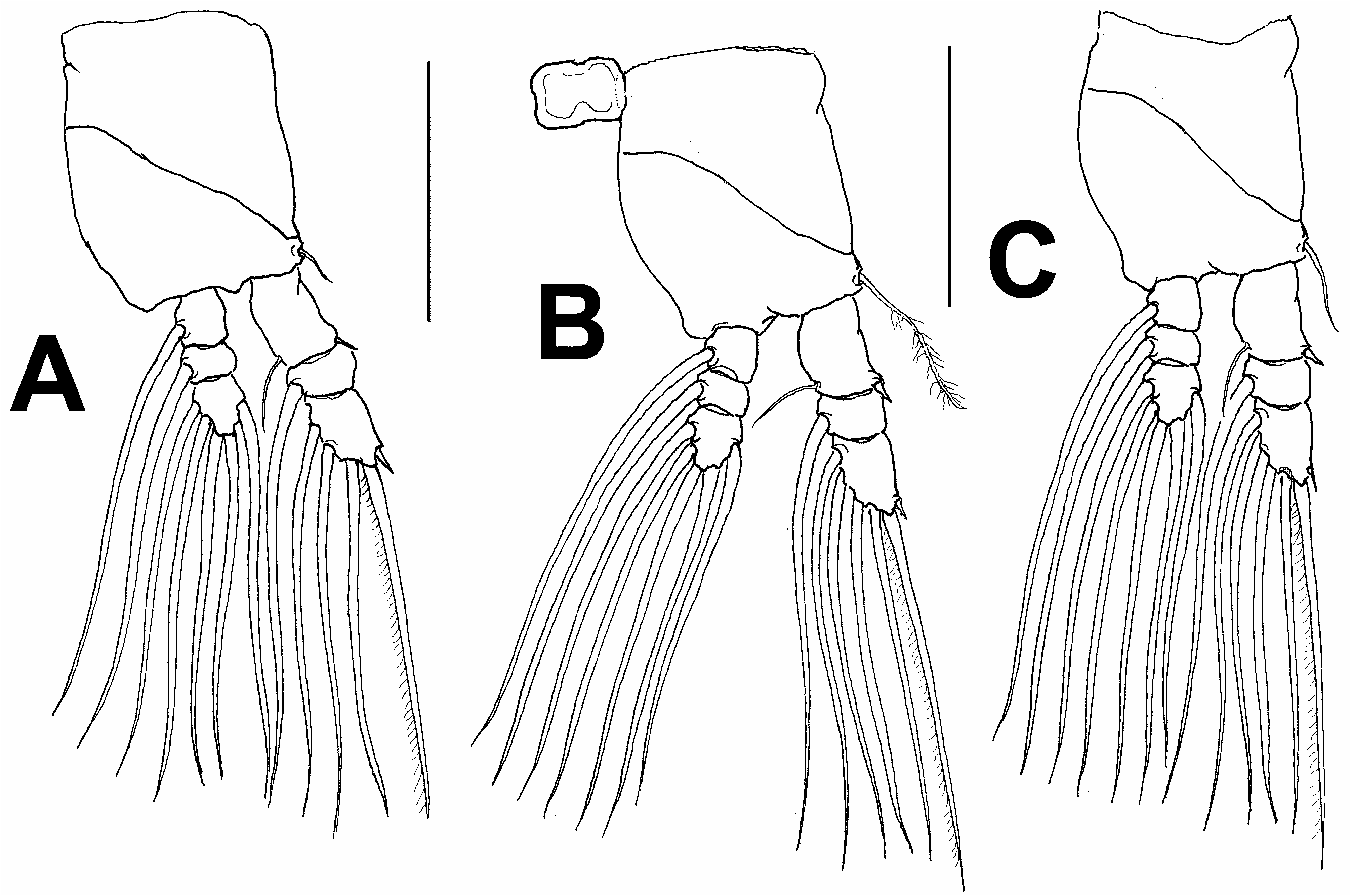
First pedigerous somite incorporated into cephalothorax; this and succeeding three free pedigerous somites each bearing pair of biramous swimming legs. Pedigerous somites 2–4 together accounting for 33% of total body length in dorsal view. Intercoxal sclerites of legs 1–4 sub-rectangular, without ornamentation on surface or along distal margin. Basis of legs articulating with rectangular coxa along diagonal line. Basis with thin, naked outer basipodal seta on legs 1, 2, and 4; on leg 3, this seta thicker, lightly setulate, and 3.5 times longer than that on leg 1, 2 times longer than those on legs 2 and 4, reaching well beyond distal margin of second exopodal segment ( Fig. 8B View FIGURE 8 ). Endopodites and exopodites of swimming legs 1–4 triarticulate ( Fig. 8A–C View FIGURE 8 ). Ramus setae all lightly and biserially plumose except for spiniform outer seta on exopodal segments 1 and 3 and inner seta of first exopodal segment, these all being short and slender. Outer apical exopodal seta of swimming legs 1–4 with outer margin smooth, inner margin lightly setulate.
Armature formula of swimming legs as in Monstrillopsis hastata sp. nov.
Fifth legs absent. Urosome consisting of four somites: fifth pedigerous somite, genital somite with genital apparatus, one free postgenital somite, and large anal somite possibly incorporating fused preanal somite as described above in Mon. hastata . Anal somite comprising 35% of length of urosome. Dorsal surface of genital somite with row of hyaline papillae along distal margin. Ventral surface of genital somite forming enlarged base of short, cylindrical shaft with distal genital lappets. Genital complex of type II. Lappets represented by pair of short, posteriorly directed but moderately divergent bulbous processes ( Fig. 6 C, D View FIGURE 6 ); lappets reaching to about midlength of anal somite. Caudal rami subrectangular, weakly divergent, approximately 1.8 times longer than wide, each ramus bearing five setae.
Female: unknown.
Type locality. Giralia Bay in Exmouth Gulf , Western Australia (22°28’S; 114°20.75’E) GoogleMaps .
Etymology. The specific epithet, a noun in apposition, is from the Latin noun nanus , meaning dwarf, to emphasize the size of this species, the smallest among its known congeners. The noun is used in masculine gender to match that of the specimen examined.
Diagnosis. Remarkably small species, total body length 0.50 mm. Terminal sabre-like process of fifth antennule relatively short, representing 50% of segment length. Genital complex of type II, with short shaft and globose terminal lappets. Anteriormost section of prosome, between antennules, with distinctive nubbly surface. Oral papilla situated at 35% of length of cephalothorax. Caudal rami with five caudal setae.
Remarks. With a total body length of 0.50 mm, Mon. nanus sp. nov. is the smallest Monstrillopsis known, followed by Mon. cahuitae from Central America ( Suárez-Morales et al. 2013) that measures 0.55 mm. Among the Monstrilloida , only Monstrilla pygmaea Suárez-Morales, 2000 (0.43 mm) and Mon. minuta Isaac, 1975 (0.49 mm) are smaller. In addition, the proximal rounded process of the fifth antennular somite is clearly weaker than in the other three Australian congeners and the terminal sabre-like process is relatively shorter than in the other Australian species (50% vs. 58–64% of fifth segment length). The genital apparatus of Mon. nanus , with a short shaft and globose terminal lappets, also differs from that found in its Australian congeners. This combination of characters has not been found in any other species of Monstrillopsis .


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |