
Oxynaspis ryukyuensis, Chan, Benny K. K. & Hayashi, Ryota, 2012
|
publication ID |
https://doi.org/ 10.5281/zenodo.281652 |
|
DOI |
https://doi.org/10.5281/zenodo.6174491 |
|
persistent identifier |
https://treatment.plazi.org/id/03FC87D4-FFD3-8A17-C7D7-22E753F9F81F |
|
treatment provided by |
Plazi |
|
scientific name |
Oxynaspis ryukyuensis |
| status |
sp. nov. |
Oxynaspis ryukyuensis n. sp.
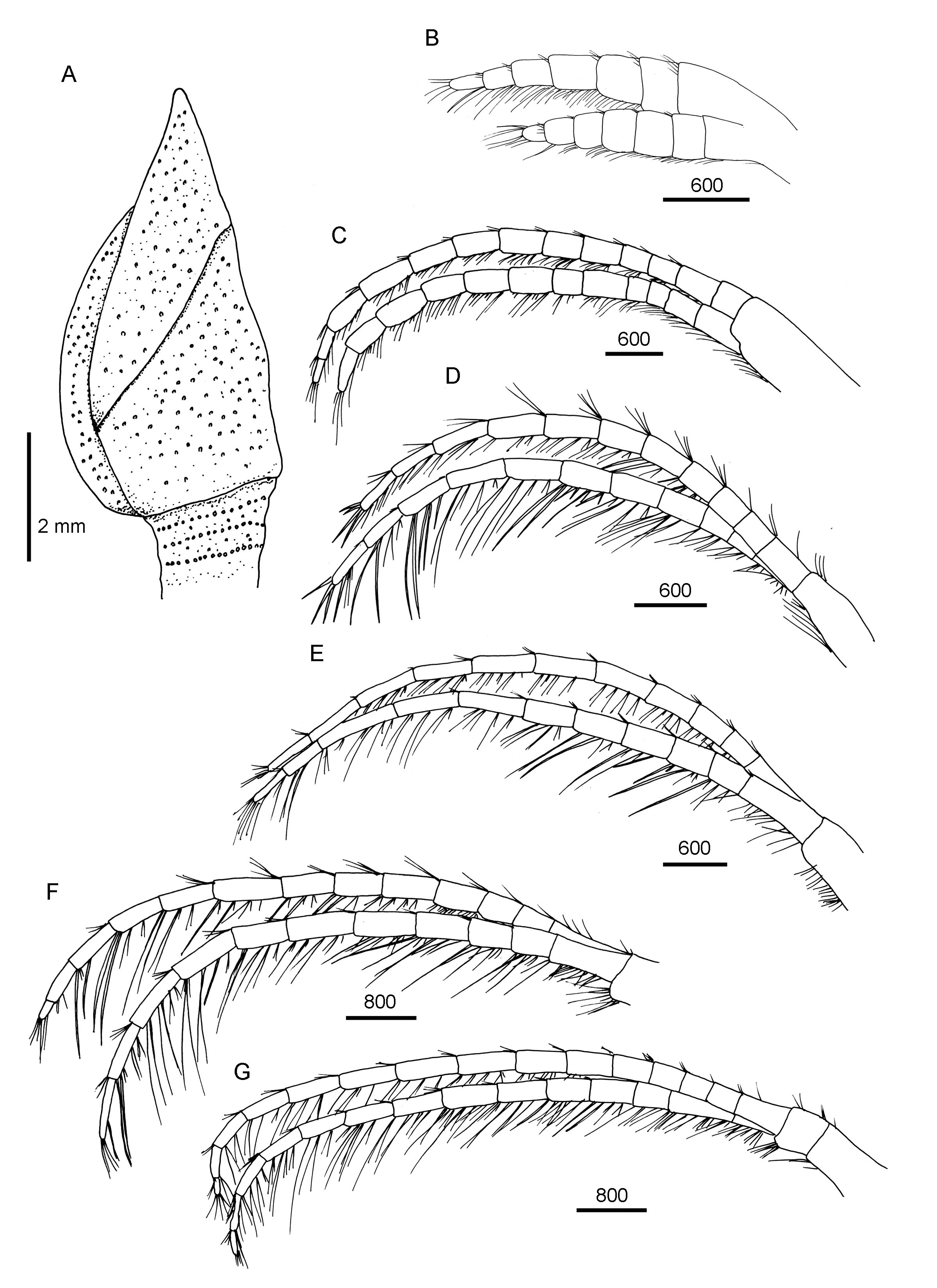
( Figs 2 View FIGURE 2 D, 9–11)
Material examined. Holotype: RUMF-ZC-1558, one specimen (CL 5.4 mm) on an antipatharian coral; trawl 75, 26°19.586´N, 126°42.453´E, 19 Nov. 2009, depth 137– 156 m. Paratypes: RUMF-ZC-1559, one specimen (CL 5 mm), same data as holotype. NHMS-6690-002, one specimen (CL 5.8 mm), same data as holotype.
Diagnosis. Capitulum elongated, with 5 completely calcified, thin plates. Tergum triangular, apex acute, high. Scutum quadrangular, occludent margin almost parallel with carinal margin. Carina slightly concave, umbo of carina located very close to basal region of carina, length of distal arm of carina about 7.5 times length of basal arm. Two filamentary appendages on dorsal side of somatic body.
Description. Hermaphrodite, capitulum narrow, surface covered by antipatharian; 5 fully calcified’ thin plates, covering whole capitulum ( Figs 2 View FIGURE 2 D, 9A). Carina slightly concave, apex reaching middle of carinal margin of tergum, umbo located very close to basal region of carinal margin, base non-forked ( Fig. 11 View FIGURE 11 C, D), length of distal arm 7.5 times length of basal arm ( Figs 2 View FIGURE 2 D, 9A). Tergum triangular, narrow, apex acute, umbo apical, occludent margin shortest, umbo apical ( Figs 2 View FIGURE 2 D, 9A, 11B). Scutum trapezoid, umbo at middle portion of occludent margin, basal margin straight, perpendicular to occludent margin, carinal margin shortest ( Figs 2 View FIGURE 2 D, 9A, 11A).
Maxilla rounded, simple setae on all margin ( Fig. 10 View FIGURE 10 A, B). Maxillule strongly notched ( Fig. 10 View FIGURE 10 C), 2 large spines above notch, region below notch protuberant, several short, simple setae in notch, approximately 8 setae below notch ( Fig. 10 View FIGURE 10 D). Mandible with4 teeth, first separated from remainder, fourth smallest, lower margin short, with 3 short pectinations, inferior angle with 1 sharp pectination ( Fig. 10 View FIGURE 10 E, F). Mandibular palps elongated, simple setae distally and on exterior margin ( Fig. 10 View FIGURE 10 G, H).
Cirrus I with anterior and posterior rami 6-segmented, anterior ramus slightly longer ( Fig. 9 View FIGURE 9 B); setae on both rami simple ( Fig. 11 View FIGURE 11 E). Cirrus II with anterior and posterior rami 13- and 12-segmented, respectively ( Fig. 9 View FIGURE 9 C); setae on both rami simple ( Fig. 11 View FIGURE 11 F, G). Cirri III and IV with anterior and posterior rami 11-segmented ( Fig. 9 View FIGURE 9 D, E). Cirrus V with anterior and posterior rami 12- and 11-segmented ( Fig. 9 View FIGURE 9 F). Cirrus VI with anterior and posterior rami 15- and 14-segmented, respectively ( Fig. 9 View FIGURE 9 G). Intermediate segments of posterior rami of cirri III to VI with 3 pairs of long simple setae and 1 pair of short simple setae ( Fig. 11 View FIGURE 11 H). Caudal appendages absent. Penis half length of cirrus VI ( Fig. 11 View FIGURE 11 J). Dorsal margin of somatic body with 2 narrow, long filamentary appendages ( Fig. 11 View FIGURE 11 I).
Distribution. Currently only known from Kumejima Island, Ryukyu Islands.
Etymology. The name ryukyuensis denotes the type locality, the Ryukyu Islands.
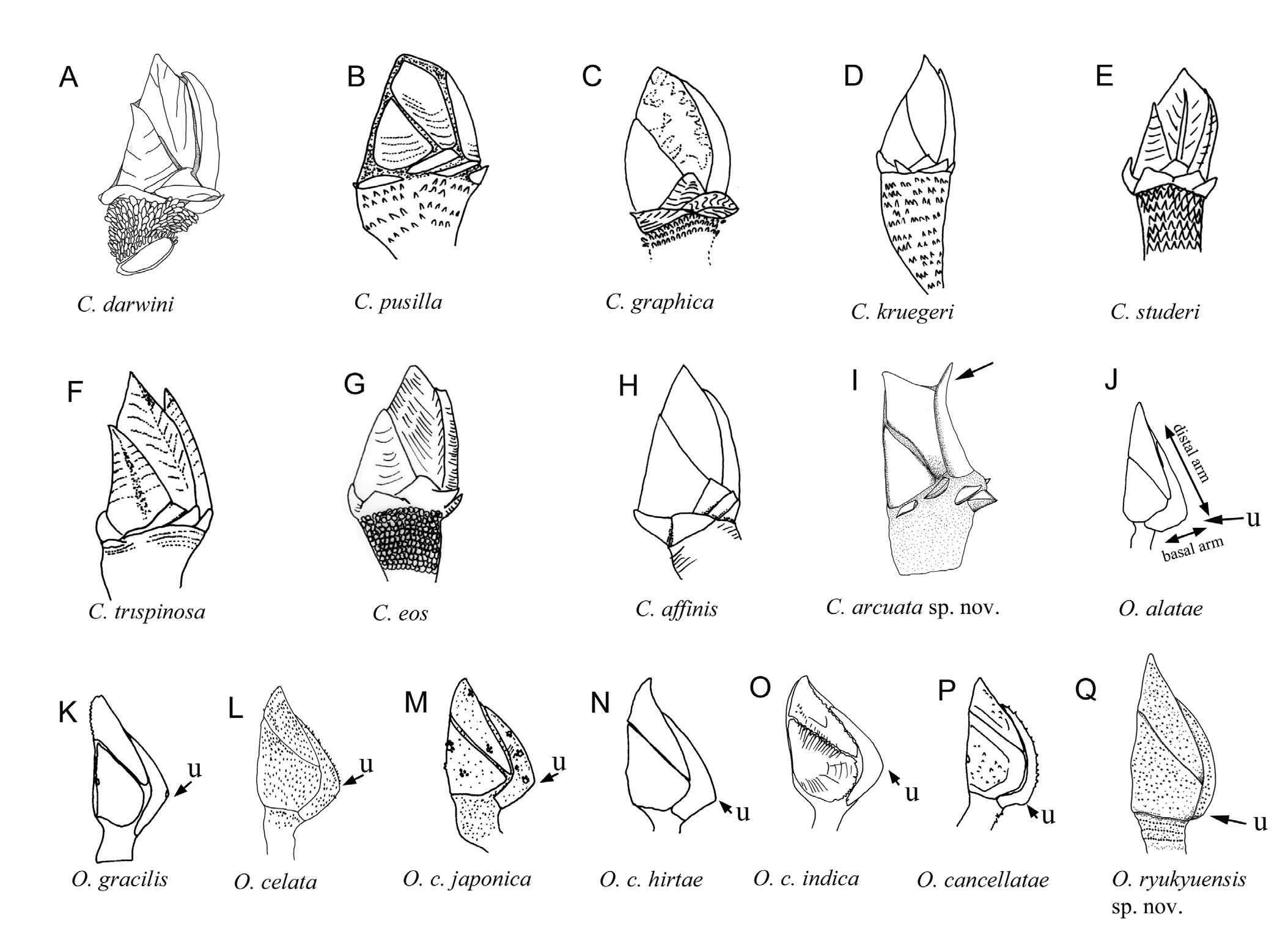
Remarks. Previous studies on the diversity of Oxynaspis have received very scant attention. A recent revision of the Oxynaspididae ( Van Syoc & Dekelboum 2011) erected two new genera, Pycnaspis Van Syoc & Dekelboum, 2011 and Minyaspis Van Syoc & Dekelboum, 2011 , in addition to Oxynaspis . Pycnaspis is a monotypic genus, containing the thick and strong plated P. connectens ( Broch, 1931, = ex Oxynaspis connectens ). Miyaspis contains 15 species, all of which have a capitulum partially covered by plates and a basally forked carina ( Van Syoc & Dekelboum 2011). Oxynaspis presently contains nine species ( Oxynaspis celata Darwin, 1852 , O. gracilis Totton, 1940 , O. alatae Totton, 1940 , Oxynaspis cancellatae Totton, 1940 , Oxynaspis pacifica Hiro, 1931 , Oxynaspis rossi Newman, 1972 , O. perekrestenkoi Van Syoc & Dekelboum, 2011 , O. joankovennae Van Syoc & Dekelboum, 2011 and O. joandianeae Van Syoc & Dekelboum, 2011 ) and four sub-species of O. celata (see remarks on O. celata above; Van Syoc & Dekelboum 2011). In Zevina’s (1982) revision of the poecilasmatid barnacles, Oxynaspis cancellatae Totton, 1940 was considered a junior synonym to O. connectens Broch, 1931 , while Newman (1972) considered them two separate species. Even Van Syoc & Dekelboum (2011) erected a new genus Pycnaspis to accommodate O. connectens , due to its distinctly strong and thick capitular plates. In the present study, we adopt the classification of Van Syoc & Dekelboum (2011). No other Oxynaspis species are reported to be junior synonyms of O. gracilis and O. alatae (see Zevina, 1982).
Oxnaspis pacifica , O. rossi , O. perekrestenkoi , O. joankovennae and O. joandianeae have large uncalcified spaces between the scutum and carina, differing from O. ryukyuensis sp. n., which has the capitulum completely covered by the capitular plates.
The location of the umbo on an opercular plate is an important taxonomic character in identifying stalked barnacles (see Zevina, 1982). In Oxynaspis , the umbo of the scutum is located at the occludent margin; the location of the umbo on the carina, located at the middle or below the middle portion of the carinal margin, is a morphological characteristic used to identify Oxynaspis species ( Newman, 1972; Van Syoc & Dekelboum 2011). Annandale (1909) and Totton (1940) measured the length ratio of the distal arm (carinal length above the umbo) and basal arm (carinal length below the umbo) of the carina of Oxynaspis for species comparisons. Comparing the present specimen with Oxynaspis celata collected in the present study (see O. celata in the previous section), the location of the umbo on the carina exhibited great differences between O. celata and O. ryukyuensis n. sp. The umbo of the carina of O. celata was located in the proximal one third of the carina and the length of the distal arm was about twice the length of the basal arm. In O. ryukyuensis n. sp., the location of the carinal umbo was located very close to the basal region, with the length of the distal arm 7.5 times greater than the basal arm (Table 1). Both specimens of O. celata and O. ryukyunesis n. sp. were adults with mature egg masses and thus such differences in the locations of the umbones should not be due to age variation.. In addition, the dorsal side of the somatic body of O. ryukyuensis n. sp. has two filamentary appendages, whereas filamentary appendages were absent from O. celata , suggesting that O. ryukyuensis n. sp. differs from O. celata (Table 1).
The location of the carinal umbo has been described and clearly illustrated for O. celata Darwin, 1852 , O. celata japonica Broch, 1922 , O. celata hirtae Totton, 1940 , O. celata indica Annandale, 1902 (illustrated in figs 12–14 in Totton 1940; redrawn in Fig. 16 View FIGURE 16 ), O. cancellatae Totton, 1940 , O. gracilis Totton, 1940 , and O. alatae Totton, 1940 . The umbo of the carina in O. celata (see pl. 3 in Darwin 1852; Fig. 16 View FIGURE 16 L), O. celata hirtae (see figs 10, 11 in Totton 1940; Fig. 16 View FIGURE 16 N), O. celata japonica (see fig. 32 in Broch 1922; Fig. 16 View FIGURE 16 M), O. celata indica (see figs 12–14 in Totton 1940; Fig. 16 View FIGURE 16 O), O. gracilis (see fig. 8 in Totton 1940; Fig. 16 View FIGURE 16 K) and O. alatae (see fig. 3 in Totton 1940; Fig. 16 View FIGURE 16 J) are expanded, forming a distinct angle and are located at the middle or proximal one third of the carina, differing from the specimens of O. ryukyuensis n. sp., which have the umbo located very close to the basal region of the carina ( Fig. 16 View FIGURE 16 Q; Table 1). Measuring the type illustration of Oxynapsis species mentioned above (except O. indica illustrated in Totton 1940), the length ratio of the distal arm to basal arm was 1.9 in O. alatae , 1.5 in O. gracilis , 1.9 in O. celata , 2.1 in O. celata japonica , 2.4 in O. c. hirtae , 1.7 in O. c. indica , obviously differing from 7.5 in O. ryukyuensis n. sp. (Table 1). The basal location of the carinal umbo of O. ryukyuensis n. sp. is consistent in the holotype and the two paratypes. The length ratio of the distal arm to the basal arm is 7.9 and 8.0 for the two paratypes. There is no detailed description of O. c. novazelandica in Broch (1922) but O. c. novazelandica was described as morphologically very close to O. c. indica , with only the maxillule differing between the two species. In the present study, we believe the external morphology of O. c. novazelandica is close to that of O. c. indica and thus morphologically differs from O. ryukyuensis n. sp., therefore confirming that O. ryukyuensis n. sp. does not belong to the O. celata cryptic species group.
Oxynaspis cancellatae (see fig. 1 in Totton 1940; Fig. 16 View FIGURE 16 P) and O. ryukyuensis n. sp. have the carinal umbo located close to the basal region (see Fig. 16 View FIGURE 16 Q). The carinal umbo of O. cancellatae is located higher than in O. ryukyuensis n. sp., resulting in the length ratio of the distal arm to the basal arm in O. cancellatae as 5.7, shorter than that of O. ryukyuensis n. sp., which is 7.5. The base of cirrus I of O. cancellatae has a single filamentary appendage (see fig. 13b in Broch 1931) but a filamentary appendage is absent from the base of cirrus I of O. ryukyuensis n. sp. ( Fig. 9 View FIGURE 9 B). The capitulum with completely calcified plates, the location of the carinal umbo and the absence of a filamentary appendage at the base of cirrus I define O. ryukyuensis n. sp..



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |