
Opisthotropis voquyi, Ziegler & David & Ziegler & Pham & Nguyen & Le, 2018
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4374.4.2 |
|
publication LSID |
lsid:zoobank.org:pub:3B7D44F9-45E7-46BA-BE29-AA568199B205 |
|
DOI |
https://doi.org/10.5281/zenodo.5962213 |
|
persistent identifier |
https://treatment.plazi.org/id/03FC87B5-A227-F51D-1CD9-F9A7FBDEFE2E |
|
treatment provided by |
Plazi |
|
scientific name |
Opisthotropis voquyi |
| status |
sp. nov. |
Opisthotropis voquyi sp. nov.
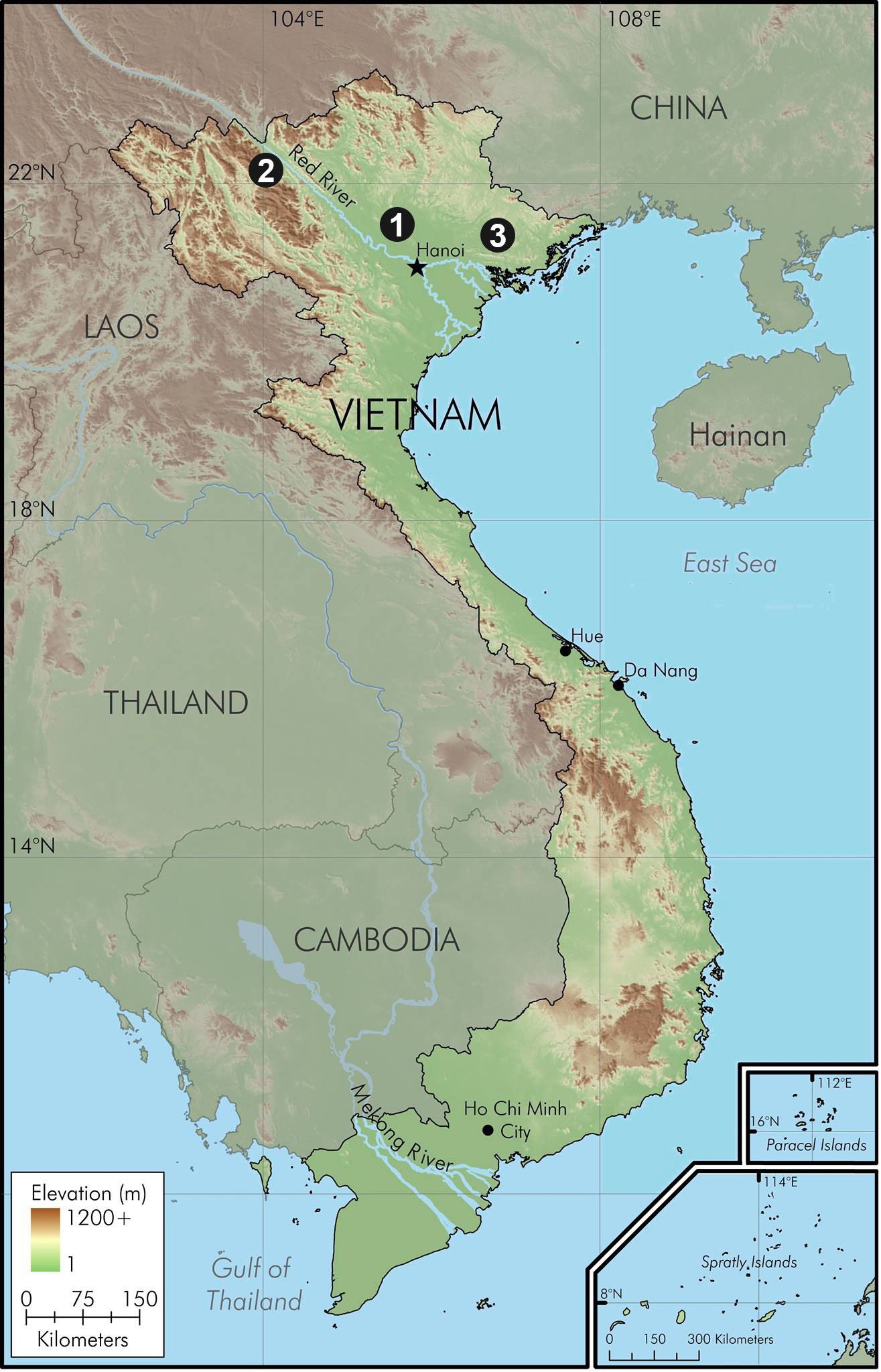
Holotype. IEBR 4326 [Field No. TYT.2013.4], an adult male, from Tay Yen Tu Nature Reserve, Bac Giang Province, 437 m asl., Vietnam, collected by Cuong The Pham et al. on 14 June 2013 ( Figs. 6–8 View FIGURE 6 View FIGURE 7 View FIGURE 8 ).
Paratypes. VNMN 0 6315 [Field No. TYT 2013.1], male, collected by Cuong The Pham et al. in June 2013; VNMN 0 6316 [Field No. BG 2015.13], female, collected by Cuong The Pham et al. on 17 May 2015; IEBR 4327 [Field No. BG 2015.33], female, collected by Cuong The Pham et al. on 22 May 2015; IEBR 4328 [Field No. BG 2015.34], female, collected by Cuong The Pham et al. on 22 May 2015; ZFMK 100819 [Field No. BG 2015.35], female, collected by Cuong The Pham et al. on 22 May 2015; ZFMK 100820 [Field No. TYT 2014.11], female, collected by Hang An Thi in May 2014, all paratypes same locality data as the holotype ( Figs. 8–10 View FIGURE 8 View FIGURE 9 View FIGURE 10 ).
Bourret & Pa) Sa Bourret & Pa) Sa Chapa () 1934 a? Tamdao) b Chapa) (a 1935 Chapa (a 1935) Chapa a (1935) Tam Dao (1935) b Dao Tam (b) 1935 NganSon c 1935 ()) (1936 Angel 1933 (syntype Angel (1933 syntype. 259 M Bourret A. 57 1934 (. M 518 Bourret 953 X. Bourret. 954 X Bourret 451 M. Bourret 794. X Bourret M. 498 Bourret Bourret ...Continued on next page TABLE 2b. (Continued) Diagnosis. A species of the genus Opisthotropis , characterized by a combination of the following characters: (1) internasal not in contact with loreal; (2) one preocular; (3) usually two postoculars; (4) one anterior temporal; (5) one posterior temporal; (6) 7 or 8, rarely 9 supralabials; (7) 25 maxillary teeth; (8) subcaudals 74–86; (9) 15 dorsal scale rows at neck, at midbody and before vent; (10) body scales smooth or only with few faint keels; (11) dorsal scales being grey, greyish-brown or brown in preservative, posteriorly more or less edged with pale greyishbrown.
Description of holotype. Body stout, oval in cross section; head short, slightly broader than neck, depressed, dorsally covered with large shields; nostrils in dorsal position and oblique, narrow, piercing the middle of nasal, which is divided beneath; eye small, pupil round.
Size. SVL: 337 mm; TaL: 106 mm; TL: 443 mm; ratio TaL/TL: 0.239.
Dentition. Maxillary teeth: right upper maxilla with 25 subequal teeth or sockets, without diastema.
Body scalation. Dorsal scale rows 15–15–15, smooth, with few faint keels.
181 ventrals (+ 1 preventral); 85 subcaudals, all paired; precloacal divided.
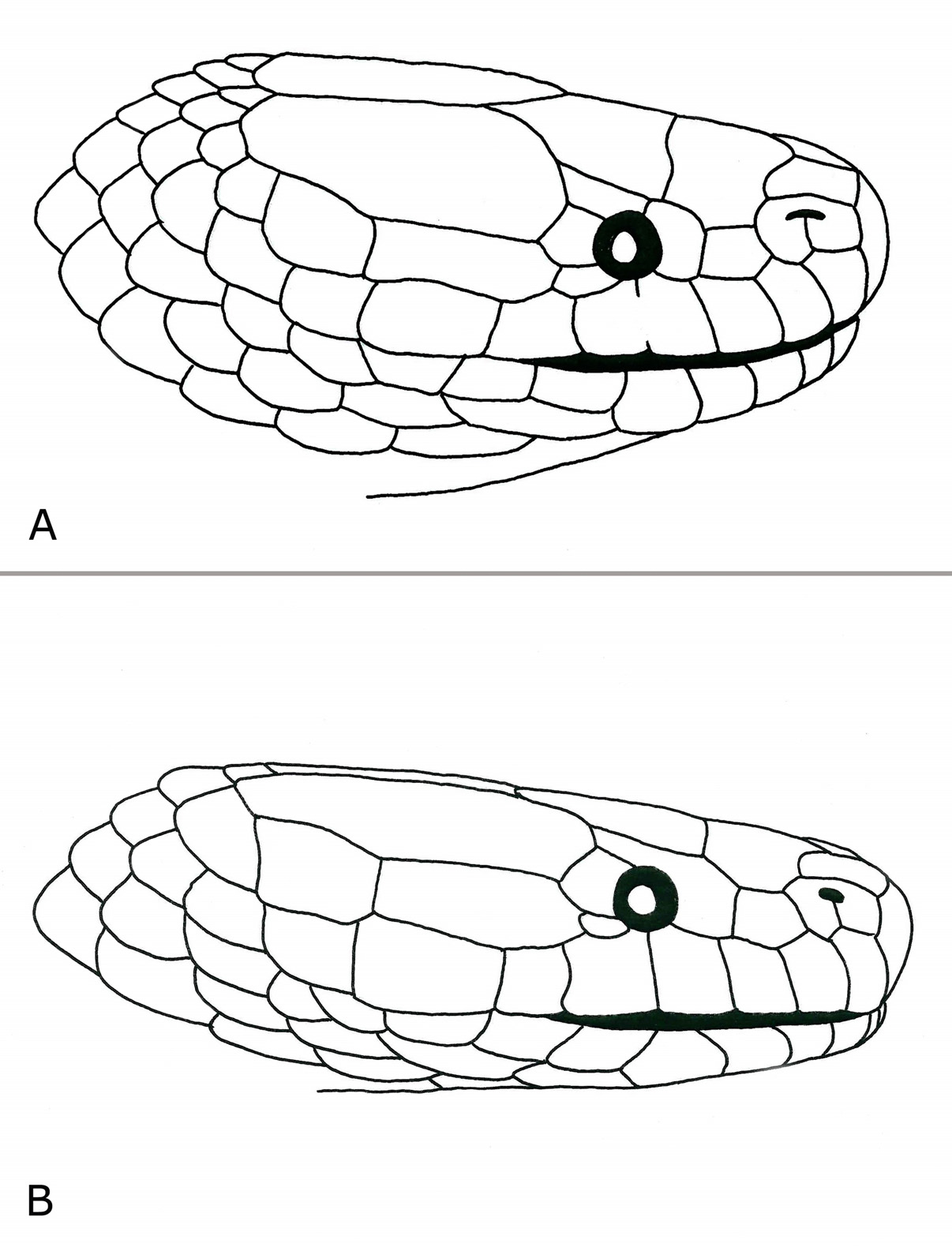
Head scalation. Rostral heptagonal, wider than high, readily visible from above; nasals large, much longer than high, divided below nostril by a distinct furrow; internasals two, strongly curved, longer than wide, in contact with rostral, nasals, and prefrontal; prefrontal single, anteriorly pointed, much broader than long, in contact with internasals, nasals, loreals, preoculars, and frontal; frontal pentagonal, slightly wider than long, apex directed posteriorly, 1.3 times longer than prefrontal; parietals longer than wide, in contact approximately the length of frontal; 1 / 1 supraocular, distinctly wider than high; 1 / 1 loreal, pentagonal, slightly wider than high, not in contact with internasal; 1 / 1 preocular, large, hexagonal, distinctly higher than wide, reaching frontal, in broad contact with prefrontal; 2 / 2 postoculars, upper largest, curved, higher than wide, lower distinctly wider than high; 8 / 7 supralabials, anterior ones distinctly higher than long, fourth entering orbit (the fourth supralabial on the right side fused with the subsequent supralabial), SL 7 / 6 largest; 1+1 / 1+1 temporals, anterior one very long and narrow, in broad contact with SL 6–7 / 5–6 and parietals, posterior one more strongly developed; infralabials 9 / 9, first pair in contact behind small mental, IL 1–4 in contact with anterior chin shields, IL 5 largest, apex directed posteriorly; posterior chin shields about the same length than anterior ones, separated from each other by 2 + 4 scales.
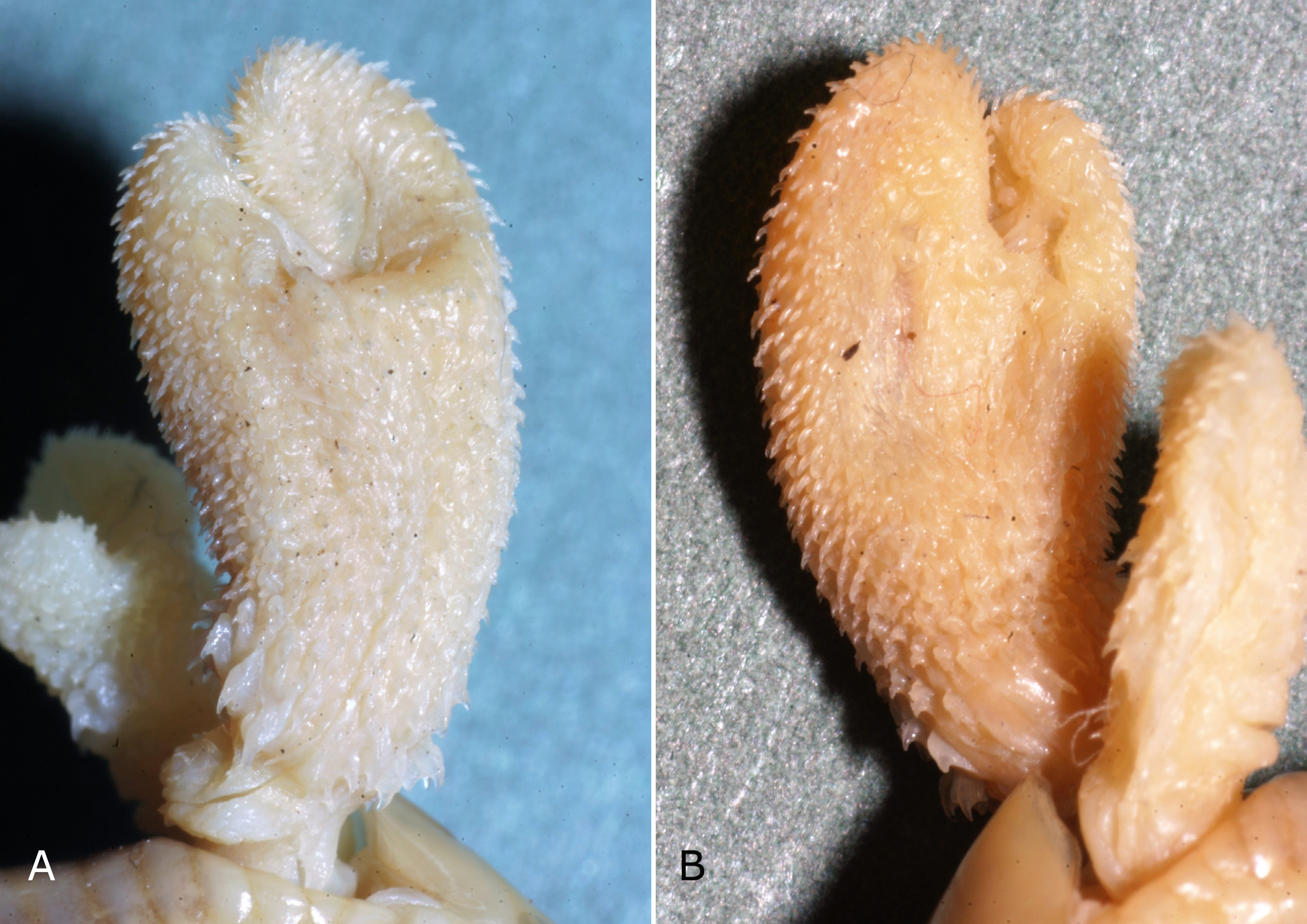
Hemipenes. Only the left hemipenis fully everted, ending in two short, inwards curved lobes ( Fig. 7 View FIGURE 7 ). Sulcus spermaticus undivided, extending only to the slightly smaller lobe. Hemipenes covered with small spines, curved backwards. Only the terminal area of the hemipenes between the lobes without spines. Upper truncus of hemipenes with ring of enlarged spines. Largest spines extending from sulcus spermaticus to hemipenis sides, with the lowermost one being the most distinct. Lower truncus of hemipenes covered with tiny spines.
Coloration (in preservative). Upper head and body surface brown, the venter yellowish-beige. Dorsal scales posteriorly more or less edged with pale brown. Dorsal head surface paler around nostrils and in the middle of parietals. Infralabials same colour as dorsal scales, adjoining scales becoming gradually paler towards mental groove. Outermost edges of ventrals and subcaudals the same colour as dorsal scales. Subcaudals speckled with small dark spots, becoming more numerous towards the tail tip.
Variation of paratypes. An overview of the meristic and scalation data of the type series of Opisthotropis voqyui sp. nov. is given in Table 3. The dorsal ground coloration varies between greyish, greyish brown, and brown. Concerning a potential sexual dimorphism, males seem to be brown, females grey or greyish brown. Females also seem to grow larger. The dorsal tail scales in the males had few faint keels, which was only the case for one female; the remaining four females had smooth dorsal tail scales. The paratype series generally accorded well with the holotype description. Of the type series six individuals had two postoculars, only one individual had 2 postoculars on the left side and one postocular on the right side.
Comparisons. The comparison of Opisthotropis voquyi sp. nov. with other species of the genus Opisthotropis and the genera Parahelicops and Paratapinophis was based on the following references: Stuart & Chuaynkern (2007), David et al. (2011), Teynié et al. (2013), Nguyen et al. (in press), and Wang et al. (2017).
There are only few species of Opisthotropis and other genera with 15 midbody dorsal scale rows (see Table 4).
15 15 or 17 17 19 23
Opisthotropis alcalai Brown & Leviton, 1961 x O. andersonii (Boulenger, 1888) x
O. balteata (Cope, 1895) x O. cheni Zhao, 1999 x
O. cucae David, Pham, Nguyen & Ziegler, 2011 x O. daovantieni Orlov, Darevsky & Murphy, 1998 x
O. durandi Teynié, Lottier, David, Nguyen & Vogel, 2013 x
O. guangxiensis Zhao, Jiang & Huang, 1978 x
O. jacobi Angel & Bourret, 1933 x
O. kikuzatoi Okada & Takara, 1958 x
O. kuatunensis Pope, 1928 x O. lateralis Boulenger, 1903 x
O. latouchii (Boulenger, 1899) x
O. laui Yang, Sung & Chan, 2013 x ...Continued on next page Main additional differences between Opisthotropis voquyi sp. nov. and Parahelicops annamensis are: one preocular in the new species versus two in P. annamensis ; internasal not in contact with loreal in the new species versus in contact in P. annamensis ; body scales smooth or only with few faint keels in the new species versus keeled throughout in P. annamensis and especially strongly keeled on the posterior part of the body and the base of the tail; body dorsum grey to greyish-brown or brown, with dorsals posteriorly more or less edged with pale greyish-brown in the new species versus dorsum with orange bars and spots in P. annamensis .
Opisthotropis voquyi sp. nov. differs from O. guangxiensis by 7–9 versus 9–10 supralabials, one posterior temporal (versus two), 74–86 versus 51–58 subcaudals, dorsal scales arranged in 15–15–15 versus 17–15–15 rows, and a greyish to greyish–brown or brown dorsum, dorsal scales posteriorly more or less edged with pale greyishbrown in the new species versus dark body with pale crossbars in O. guangxiensis .
Opisthotropis voquyi sp. nov. differs from O. kikuzatoi by 7–9 supralabials (versus 6), one preocular (versus two), one anterior temporal (versus two), and the greyish to greyish-brown or brown dorsal pattern, with the dorsals posteriorly being more or less edged with pale greyish-brown in the new species (versus dark dorsum with dorsolateral orange spots).
Opisthotropis voquyi sp. nov. differs from O. maculosa by a grey to greyish-brown or brown dorsum, with dorsals being posteriorly more or less edged with pale greyish-brown in the new species versus with yellow dots on a dark background in O. maculosa .
In external morphological characters, O. voquyi sp. nov. is particularly similar to O. jacobi , from which it differs in usually having two postoculars versus only one in O. jacobi , in the number of maxillary teeth: 25 in the new species versus 19–23 in O. jacobi , and in the dorsal scales being grey to greyish–brown or brown versus blackish–brown in O. jacobi .
Etymology. The specific epithet is a noun in the genitive case. This species is named in honour of Professor Doctor Vo Quy of Vietnam National University, Hanoi, who passed away on 10 January 2017, and who played a pioneering role in nature conservation in Vietnam. He inspired later generations of conservation biologists, including authors of this study.
Suggested common names. Vo Quy’s Mountain Stream Keelback (English) and Rắn trán võ quý (Vietnamese).
Distribution. Opisthotropis voquyi is currently known only from the type locality in Bac Giang Province ( Fig. 11 View FIGURE 11 ).
Natural history. Specimens were found at night between 19:30 and 23:00 h in and around small rocky streams, where the snakes were observed in the water or on the rocks close to the water. The surrounding habitat was secondary evergreen forest of medium and small hardwoods mixed with bamboo, shrubs, and vines ( Fig. 12 View FIGURE 12 ). The air temperature was 29.2–36.5 o C and the relative humidity was 55–73%. Other species co-occurring with Opisthotropis voquyi include Cyclophiops multicinctus , Opisthotropis lateralis , Sinonatrix percarinata , Gekko palmatus , Sphenomorphus cryptotis , and Shinisaurus crocodilurus vietnamensis .




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Opisthotropis voquyi
| Ziegler, Thomas, David, Patrick, Ziegler, Tim N., Pham, Cuong T., Nguyen, Truong Q. & Le, Minh D. 2018 |
Opisthotropis voquyi
| Ziegler & David & Ziegler & Pham & Nguyen & Le 2018 |
Opisthotropis voquyi
| Ziegler & David & Ziegler & Pham & Nguyen & Le 2018 |
Opisthotropis voquyi
| Ziegler & David & Ziegler & Pham & Nguyen & Le 2018 |
Opisthotropis voquyi
| Ziegler & David & Ziegler & Pham & Nguyen & Le 2018 |
Opisthotropis voquyi
| Ziegler & David & Ziegler & Pham & Nguyen & Le 2018 |
O. durandi Teynié, Lottier, David, Nguyen & Vogel, 2013 x
| Teynie, Lottier, David, Nguyen & Vogel 2013 |
O. laui
| Yang, Sung & Chan 2013 |
O. cucae
| David, Pham, Nguyen & Ziegler 2011 |
O. tamdaoensis
| Ziegler, David & Vu 2008 |
O. maculosa
| Stuart & Chuaynkern 2007 |
O. daovantieni
| Orlov, Darevsky & Murphy 1998 |
O. guangxiensis
| Zhao, Jiang & Huang 1978 |
O. guangxiensis
| Zhao, Jiang & Huang 1978 |
O. guangxiensis
| Zhao, Jiang & Huang 1978 |
O. kikuzatoi
| Okada & Takara 1958 |
Parahelicops annamensis
| Bourret 1934 |
P. annamensis
| Bourret 1934 |
P. annamensis
| Bourret 1934 |
P. annamensis
| Bourret 1934 |
P. annamensis
| Bourret 1934 |
Parahelicops annamensis
| Bourret 1934 |
O. jacobi
| Angel & Bourret 1933 |
Paratapinophis praemaxillaris
| Angel 1929 |
O. spenceri
| Smith 1918 |
O. maxwelli
| Boulenger 1914 |
O. rugosa
| Lidth de Jeude 1890 |
O. typica
| Mocquard 1890 |
O. shenzhenensis
| Wang, Guo, Liu, Lyu, Wang, Luo, Sun & Zhang 1872 |