
Periclimenes laevimanus, Ďuriš, 2010
|
publication ID |
https://doi.org/10.11646/zootaxa.2372.1.12 |
|
persistent identifier |
https://treatment.plazi.org/id/03FC7D16-FFBE-FF8C-61AB-B4187D582B4F |
|
treatment provided by |
Felipe |
|
scientific name |
Periclimenes laevimanus |
| status |
sp. nov. |
Periclimenes laevimanus View in CoL sp. nov.
( Figs. 1–6 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 )
Material. Vietnam, Nhatrang Bay , 12°10’06.0”N, 109°17’45.1”E, 23.IX.2008, sandy-mud bottom, 14 m, on hydroid, cf. Lytocarpia sp GoogleMaps ., coll. I. Marin: holotype ov. female cl 2.4 mm, RMNH D.53129; allotype male cl 1.8 mm, RMNH D.53130; paratypes: ov. female cl 2.1 mm (preserved in 96% ethanol), juvenile male cl 1.2 mm, juvenile female cl 1.3 mm ( RMNH D.53131); ov. female, cl 2.3 mm (dissected), juvenile male cl 1.2 mm, juvenile female cl 1.2 mm ( ION); male cl 1.7 mm, juvenile female cl 1.4 mm ( ZMMSU). One specimen kept in the author’s collection, ov. female cl 2.3 mm (preserved in 96% ethanol) .
Etymology. The specific name is combined from laevis (Latin: smooth, beardless) and manus (Latin, hand) referring to the smooth, not granulated, surface of the second pereiopods, unlike the granulated second pereiopods in the most similar species, P. granulimanus .
Description. Based on holotype, ovigerous female. Small sized shrimp with slender body ( Fig. 1 View FIGURE 1 ). Carapace smooth, with well developed antennal, hepatic and epigastric spines; hepatic spine subequal and situated posterior to antennal spine; epigastric spine articulated with carapace, subequal but clearly separated by wider space from posterior dorsal rostral tooth. Rostrum long and slender, overreaching intermediate segment of antennular peduncle, straight, with developed dorsal lamina bearing eight teeth, proximal tooth postorbital; dorsal teeth subequal, distal two slightly smaller; ventral lamina lacking, margin setose, with minute subterminal tooth. Inferior orbital angle well produced, obtusely subquadrate.
Thoracic sternites narrow, without special features.
Abdominal segments smooth, tergites non-carinate or posteriorly produced (third tergite with transverse malformation dorsally in holotype, Fig.1 View FIGURE 1 ); pleura of first to fifth abdominal segments posteroventrally rounded. Telson slender, four times as long as wide, tapering distally, with two pairs of small dorsal spines at 0.5 and 0.7 of length, three pairs of posterior marginal spines, small lateral spines subequal to dorsal spines, long and slender intermediate spines 8–9 times lateral spine’s length and about 0.3 of telson length; slender, setulose submedian spines about half of intermediate spines in length.
Eyes ( Fig. 2B View FIGURE 2 ) well developed, with cornea rounded, diameter of cornea subequal to eyestalk, latter about twice corneal length, additional pigment spot minute, situated dorsolaterally on eyestalk close to cornea.
Antennula ( Fig. 2C View FIGURE 2 ) well developed; basal segment about 2.5 times as long as wide; stylocerite narrow, acute, reaching almost to middle of basal segment, disto-lateral tooth slender, acute, overreaching produced rounded anterior margin, ventral tooth situated on middle of medial margin; intermediate and distal segment length combined about half of basal segment length; intermediate segment robust, slightly longer than wide, laterally flattened, setose; distal segment narrower but slightly longer than preceding segment; upper flagellum biramous, fused part with 5 segments, shorter free ramus consisting of 5 segments, with 8 groups of aesthetascs, longer free ramus and lower flagellum slender, filiform.
Antenna ( Fig. 2D View FIGURE 2 ) well developed, basicerite with distolateral tooth; carpocerite about twice longer than wide, not reaching middle of scaphocerite; flagellum well developed; scaphocerite exceeding antennular peduncle and rostrum, about 3.5 times as long as maximum width at its proximal third, distolateral tooth strong but not exceeding lamina.
First pereiopod ( Figs. 3A–E View FIGURE 3 ) slender, unarmed, overreaching scaphocerite by fingers; chela with fingers equal to palm length; fingers ( Figs. 3B–D View FIGURE 3 ) slender, about 5 times as long as wide, tapering distally to tiny rounded tips, feebly curved and medially, with narrow space when closed, cutting edges simple, 2–4 dense tufts of long setae on sides of both fingers; palm subcylindrical, about twice longer than deep; carpus subequal to chela length and about 4 times longer than distal width, tapering proximally; merus about 1.2 times longer than carpus and about 6 times longer than deep, ischium about 0.6 of merus length and about 3 times as long as deep, several long setae medially; basis and coxa short, with group of setae distomedially ( Fig. 3E View FIGURE 3 ).
Second pereiopods smooth, long and slender, generally similar in shape but very unequal in size. Minor pereiopod ( Figs. 3F–G View FIGURE 3 ) overreaching scaphocerite by distal three quarters of carpus, chela subequal to cl; fingers slender, about 5.5 times as long as deep proximally, slightly curved, cutting edges entire, tips sharp, hooked; palm about twice longer than fingers, slightly compressed, about 4.5 times as long as maximal depth and 6 times of width; carpus 0.6 of palm length and 3.5 times as long as proximal width, tapering proximally; merus 1.5 times longer than carpus and about 8 times longer than deep; ischium subequal to palm length, about 1.2 times longer than merus and about 10 times as long as maximal depth; basis and coxa short, without special features.
Major pereiopod ( Figs. 3H–J View FIGURE 3 ) subequal to total body length (see Fig. 1 View FIGURE 1 ), overreaching scaphocerite by distal three quarters of merus; chela ( Fig. 3I View FIGURE 3 ) about 1.7 times cl; fingers short and stout, occupying less than 0.2 of chela length, dactylus strongly hooked and tapering distally, tip pointed, cutting edge with low lamella distally, proximal half bearing three large obtuse teeth, lowering proximally, fixed finger with ventral margin straight, tip slightly upturned, cutting lamina convex distally, with three obtuse teeth on proximal half; palm about 4.5 times longer than fingers and about 8 times longer than deep, depressed, smooth; carpus about 0.4 of palm length and about 4 times as long as deep distally, tapering proximally; merus and ischium slender, uniformly deep, merus about 1.6 times longer than carpus and about 10 times longer than deep, ischium 1.1 times longer than merus; basis and coxa short, without special structures.
Third pereiopod ( Figs. 4A–B View FIGURE 4 ) slender, exceeding scaphocerite by dactylus; dactylus about 6 times longer than deep basally, slender and curved, together with long propodal spines forming grasping structure, unguis well demarcated from corpus, slender, with sharp, curved tip, with shallow proximoventral emargination ( Fig.4D View FIGURE 4 ), occupying distal third of dactylus length, rudimentary additional unguis in form of inconspicuous distoventral pointed angle on corpus, ventral margin of corpus concave; propodus about 0.6 of cl, 4 times longer than dactylus and about 11 times as long as maximal depth at distal quarter, slightly upturned dorsally, 3 pairs of long spines on distoventral quarter of palm, increasing in size proximally, 0.45–0.6 of dactylus length, 3 single ventral spines posteriorly, situated at 0.4, 0.65 and 0.8 of propodus length, decreasing in size proximally, scattered setae dorsally, longer setae dorsodistally; carpus 0.4 of propodus length and about 4 times as long as distal depth, tapering proximally; merus slightly longer than propodus, about twice longer than carpus and 12 times as long as wide; ischium as deep as and about half length of merus, basis and coxa short.
Fourth and fifth pereiopods generally similar to third leg, but increasing in length; fourth leg ( Fig. 4C–D View FIGURE 4 ) 1.15 times longer, and fifth about 1.35 times longer, than third pereiopod; propodus of fourth pereiopod with only two single ventral spines proximally to 3 distal prehensile pairs; propodus of fifth pereiopod ( Figs. 4E–F View FIGURE 4 ) with 4 pairs of grasping spines, each of 3 proximal pairs with long serrulate seta between spines, and 1 single short spine at half of propodus length.
Uropods ( Figs. 2E, H View FIGURE 2 ) slender, exceeding telson; lateral border of uropodal endopod straight on distal 2/3, with small distal tooth and slender mobile spine, about 3–4 times longer than tooth.
Mouthparts from paratype ovigerous female (cl 2.3 mm) examined ( Fig. 5 View FIGURE 5 ), left side dissected (excl. third maxilliped shown from right side). Mandible ( Fig. 5A View FIGURE 5 ) without palp, incisor process ( Fig. 5B View FIGURE 5 ) stout, with 4 terminal teeth (3 on right mandible), molar process well developed. Maxillula ( Fig. 5C View FIGURE 5 ) normal, palp feebly bilobed, lower lobe with small hooked subterminal seta, upper lacinia larger than lower lacinia, broadly rounded distally, with about 9 strong distal spines; lower lacinia strongly tapering distally to subacute apex bearing slender spines, latter also distributed along ventral margin. Maxilla ( Fig. 5D View FIGURE 5 ) with well developed simple palp without distal setae, basal endite well developed, with apex distinctly bilobed, lobes with groups of slender spiniform serrulate setae; coxal endite lacking; scaphognathite narrow, about 3 times longer than wide, with plumose marginal setae. First maxilliped ( Fig. 5E View FIGURE 5 ) with slender recurved palp bearing single preterminal seta; basal endite well developed, broad and rounded anteriorly, fused with coxal endite, endites with numerous marginal and submarginal slender spiniform setae; exopod robust, with 4 terminal plumose setae, caridean lobe well developed, with plumose marginal setae; epipod triangular in shape, feebly bilobed distally. Second maxilliped ( Fig. 5F View FIGURE 5 ) normal, dactylar segment moderately broad, 3 times longer than broad, with numerous stout serrulate setae along medial margin; distomedial angle of propodus broadly rounded, with slender setae, several simple setae on distal margin; ischium and merus fused, suture feebly demarcated; epipod simple, broadly ovate, without podobranch; exopod long, with 4 plumose setae terminally. Third maxilliped ( Fig. 5G View FIGURE 5 ) with slender segments; coxa with rounded lateral plate, and long plumose seta basally, reduced finger-like arthrobranch present; basis and ischiomerus fused, sutures feebly marked, compound segment about 6 times as long as wide basally and with serrulate setae medially, very long, mostly 0.4–0.6 of ischiomerus length, ischium with longitudinal series of about 8 short submarginal setae, merus with 3 short spinules distally in longitudinal row; penultimate segment about 5 times as long as wide and about 0.85 of ischiomerus length, with several long serrulate setae medially and distally; terminal segment about 5 times as long as wide and about 0.85 of penultimate segment, with numerous slender serrulate setae, terminal ones longer than half of segment; exopod well developed, reaching to about 0.8 of ischiomerus, with 4 long plumose setae terminally, one shorter subterminal plumose seta.
Endopod of first male pleopod ( Figs. 4 G–H View FIGURE 4 ) almost half of exopod length and twice as long as wide, broadening distally; apex angulate; medial margin with obtuse, hooked, distally directed lobe at distal third, single setulose seta and two short simple setae at proximal third; distolateral margin broadly rounded, with three setulose marginal setae, lateral margin with two simple submarginal setae at middle of segment. Endopod of second male pleopod ( Figs. 4 I–J View FIGURE 4 ) with well developed appendices; appendix interna overreaching half of endopod length, slender, about 10 times longer than distal width, with group of about 7– 8 cincinnuli distomedially; appendix masculina slightly stouter, overreaching half of appendix interna length, with three apical stout setae, 1 long, subequal to appendix length, and 2 about half length, 2 widely spaced short setae, laterally turned, situated on distal half of appendix masculina corpus; three submarginal simple curved setae situated along medial margin of endopod proximally off appendices.
Remarks. The series of 11 specimens examined shows only limited morphological variability. The rostrum formula is 2 + 7–8/1 for adults, with the maximum number of dorsal rostral teeth in the three ovigerous females only, and 1–2 + 5/0 in juveniles (one of those bearing an indistinct subterminal ventral tooth). The epigastric spine is absent in some of the smallest juvenile specimens (cl 1.2–1.3 mm); a minute one is present in the largest juvenile (cl 1.4 mm) ( Table 1), and is regularly present in the adults.
Males are generally similar to the holotype female; they differ by their smaller size, a more slender body, longer antennular and antennal flagella, and stouter pleopodal protopods. The antennules bear distinctly more aesthetasc groups, 13 in the allotype, 18 in the paratype (cl 1.7 mm, ZMMSU) .
The length ratios of the second pereiopods are also consistent in adults. The major second pereiopods of males are slightly stouter but otherwise similar to those of adult females, the minor ones have shorter chelae, about 1.5 times longer than the carpus (versus 2.5 times in the ovigerous female holotype), and with the palm about 1.5 times longer than the fingers (versus twice the finger length in the holotype). The minor second pereiopod chela, however, is extremely elongate in the holotype. The smaller ovigerous female paratype (cl 2.3 mm, ION), has segment proportions comparable to those of fully-grown males (see above). The juveniles (cl 1.2–1.4) have the second pereiopods distinctly unequal, but proportionally much shorter and stouter, reaching the end of the scaphocerites by the distal end of the major pereiopod carpus. The major cheliped of the juvenile male (cl 1.2 mm, RMNH) has the palm about twice as long as the fingers, and the carpus about 0.4 of the palm length. The minor second pereiopods have segment length ratios subequal to those described for males .
The new species is morphologically most related to P. granulimanus sharing with it a general appearance and most morphological characters, with some exceptions (see Remarks for P. granulimanus ). Shared characters include the well developed prehensile (grasping) structure on the distal propodi and dactyli of the walking legs. The dorsal border of the propodi is slightly upturned distally, and the ventral margin is distinctly swollen in their distal quarter to accommodate a closely set series of 3 to 4 pairs of long, distal spines. Thus, the distoventral spines are directed against the dactylus when flexed. The dactyli are simple, without the microscopic distoventral tooth on the corpus present in P. laevimanus sp. nov. Possibly, the prehensile legs have evolved to aid the shrimp’s movement on and attachment to slender branching cnidarian hosts such as hydroids, from which both these species were described. P. granulimanus is also known from an unidentified antipatharian ( Bruce 1978), equally a cnidarian with slenderly-built branches.
A different type of prehensile structure is developed in the species of two other hydroid-associated shrimp genera, Rapipontonia Marin, 2007 (Pontoniinae) , and Chlorotocella Balss, 1914 (Pandalidae) . The former possesses a similar dorsal curvature and distal swelling of the propodi, but the spination is formed by a pair of long spines situated some distance from the distal end and by 2 single median spines situated a little posterior to the distal pair ( Marin 2007; I. Marin pers. comm.). The posterior two-thirds of the propodus are spineless in Rapipontonia species , similarly to Periclimenes brucei , an antipatharian associate ( Ď uriš 1990). Chlorotocella gracilis specimens, collected together with the new species from the same hydroid colonies, has similarly formed walking propodi (but slightly upturned and deeper distally) and slender, simple and curved dactyli flexing against a series of some closely set and paired long spines distoventrally on the propodus, and with a dense series of unpaired spines along the remaining part of the ventral margin of the propodus (vs. 1–2 single posterior spines in both P. laevimanus sp. nov. or P. granulimanus ). These grasping structures provide an excellent example of a morphological convergence in related, as well as distinctly unrelated, shrimp species associated with similar invertebrate hosts.
Measurements. The holotype has the minor second pereiopod segment lengths (fingers: palm: carpus: merus: ischium) 0.7: 1.9: 1.1: 1.6: 1.7 mm. The two ovigerous female paratypes (cl 2.3 mm, UO; 2.1 mm, RMNH) have: 0.7: 1.2 (1.0 in the smaller female): 1.0: 1.2: 1.4 mm. Other proportions are listed in Table 1.
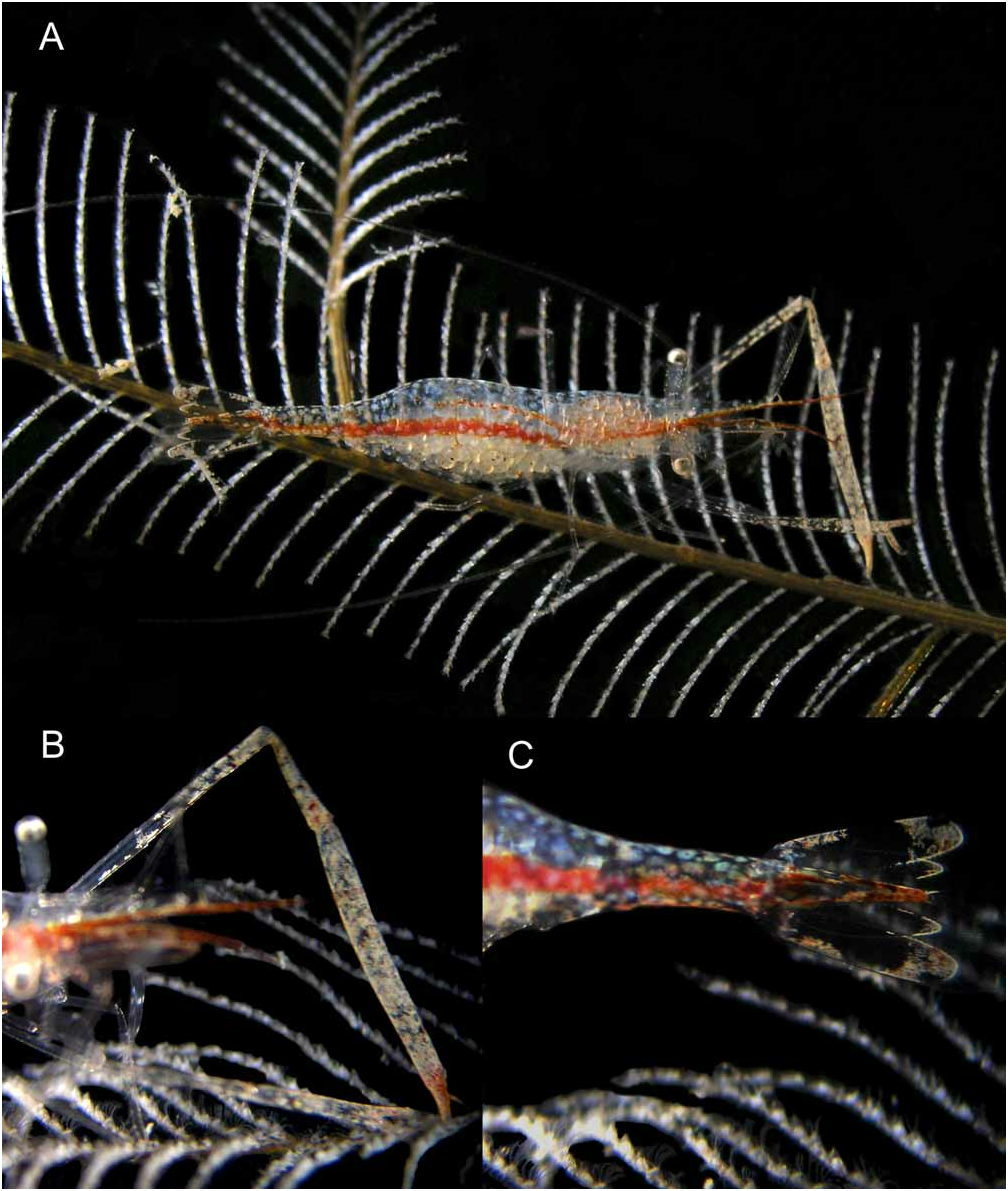
Color. Two distinct brownish-red median stripes run through the generally transparent body, covered by scattered light bluish chromatophores ( Fig. 6 View FIGURE 6 ). The anterior, narrower, median stripe originates from a pair of reddish longitudinal stripes running anteriorly through the medial portion of each antennular peduncle to their lower flagellum, through the ventral part of the cephalothorax, going up through the upper abdominal region, where it covers the digestive tube up to the third segment, and then continues posteriorly as a line of diffuse, inconspicuous chromatophores. A much wider, very distinct, red longitudinal swath is situated on the ventral portion of the abdomen; its anterior end is situated below the upper stripe, and continues to the base of the telson; the lateral margins of the telson are irregularly outlined by brown swaths connecting posteriorly. Scattered brownish-cream chromatophores are present on the uropodal exopods, the carpus and palm of the second pereiopods, and also as part of the background, reddish, coloration of the antennules. Fingers of the second pereiopod chelae are brown-cream coloured; distal segments of these appendages, are covered by a dense network of white-cream chromatophores, less markedly developed on the minor cheliped. Whitish patches are also present on anterolateral and distal parts of the uropodal exopods, and along the distal margin of the uropodal endopod. The intermediate pair of posterior telson spines is white.
Host. All individuals from the type series were collected from three grouped, branching colonies of the hydroid Lytocarpia sp. ( Hydrozoa: Leptothecata : Plumulariidae ), up to 15 cm high.
Habitat. Sandy-mud flat, gently sloping bottom off a sandy beach with alternating coastal fringing coral reef, 14 m depth.
Associated fauna. Specimens of the new species were collected together with two subadult specimens of Dasycaris sp. (Pontoniinae) and three specimens of Chlorotocella gracilis (Pandalidae) .
Distribution. Known only from the type locality: Nhatrang Bay, Vietnam, South China Sea (this report).
Periclimenes brucei Ď uriš, 1990
( Figs. 7–8 View FIGURE 7 View FIGURE 8 )
Periclimenes brucei View in CoL Ď uriš, 1990: 1, Figs. 1–2 View FIGURE 1 View FIGURE 2 .
Nec Periclimenes View in CoL ? brucei View in CoL . — Fransen 1994: 119, Figs. 85–86.
Material. Holotype ovigerous female cl 2.5 mm, ZMMU Ma 2522.
Supplementary description. The original figure of the total view of Periclimenes brucei was erroneously reproduced in a mirrored view ( Ď uriš 1990, Fig.1 View FIGURE 1 ), thus, it is presented here ( Fig.7 View FIGURE 7 ) in its correct orientation. The original description also lacks details on mouth parts; these were subsequently examined and the description and figures ( Fig. 8 View FIGURE 8 ) are also provided here.
Mandible ( Fig. 8A View FIGURE 8 ) without palp, incisor process stout, with four terminal teeth, molar process well developed. Maxillula ( Fig. 8B View FIGURE 8 ) normal, palp feebly bilobed, lower lobe with distinct hooked subterminal seta, upper lacinia larger than lower lacinia, broadly rounded distally, with about 10 strong serrulate distal spines and several simple spines ventrally; lower lacinia tapering distally with group of slender apical spines, similar ones also distributed along ventral and dorsal margins. Maxilla ( Fig. 8C View FIGURE 8 ) with well developed simple palp with two basal setae, basal endite well developed, with apex distinctly bilobed, upper lobe larger, lobes with groups of slender spiniform setae, coxal endite lacking; scaphognathite narrow, about 3 times longer than wide, with plumose marginal setae. First maxilliped ( Fig. 8D View FIGURE 8 ) with slender, distally recurved palp (no preterminal seta seen); basal endite well developed, broad and rounded anteriorly, completely fused with coxal endite, endites with numerous marginal and submarginal slender spiniform setae; exopod robust, with 4 terminal plumose setae, caridean lobe well developed, with plumose marginal setae; epipod simple, slightly longer than wide, distally rounded. Second maxilliped ( Fig. 8E View FIGURE 8 ) normal, dactylar segment moderately broad, 4 times longer than broad, with numerous serrulate setae along mesial margin; propodus with several simple setae on distal margin; ischium and merus well demarcated; epipod simple, broadly subquadrate, without podobranch; exopod long, with 4 plumose setae terminally. Third maxilliped ( Fig. 8F View FIGURE 8 ) slender; coxa with rounded lateral plate and small unilamellate arthrobranch; basis and ischiomerus fused, with suture demarcated, compound segment about 5.5 times as long as wide, with serrulate setae medially, most of setae long, 0.25–0.35 of ischiomerus length; penultimate segment about 5 times as long as wide and about 0.7 of ischiomerus length, with several long serrulate setae medially and distally; terminal segment about 4 times as long as wide and about 0.7 of penultimate segment, with numerous slender serrulate setae, terminal ones longer than half of segment; exopod well developed, reaching to about 0.7 of ischiomerus, with 4 long plumose setae terminally.
Remarks. P. brucei occupies an isolated position within the genus. Among its specific morphological characters, the most conspicuous is the extremely slender and long major second pereiopod. In the only known specimen, the adult ovigerous female holotype, this appendage overreaches the scaphocerite by almost the whole length of the merus, while in the related species, P. laevimanus sp. nov. and P. granulimanus , only the distal part of the merus reaches beyond the distal end of the antennal scale. Periclimenes brucei may be also easily distinguished from other congeners by its extremely long carpi of both second pereiopods, which are longer than the chelae and meri. In all species discussed here, the carpi are usually 2–3 times, shorter than the chelae, at least in the major second pereiopod. The ambulatory legs are very prehensile in P. brucei , with distoventrally broadened propodi bearing about 4 pairs of long spines set against the curved dactylus bearing a minute, almost lacking, additional unguis. Such a grasping mechanism is also well developed in P. laevimanus sp. nov., and P. granulimanus , but their propodi also have 1 to 3 single, shorter proximal spines, which are lacking in P. brucei .
During this study, we had a possibility to examine a series of specimens of Periclimenes ? brucei (RMNH D.42886, D.42889), collected from unidentified white gorgonians in the Seychelles ( Fransen 1994). All these specimens, mostly juveniles or subadults, have the rostrum with a rather concave ventral margin, and the epigastric spine, lacking in some specimens, is placed more closely to the posterior tooth of the rostral series, equally to a distance of the last 2 teeth, in comparison with the holotype of P. brucei . Carpi of the first pereiopods are about 0.75 of the chelae length. The second pereiopods are subequal and short, rather than distinctly unequal and long, reaching beyond the scaphocerite only by the distal part of the palm, not by the merus. The chela of the major second cheliped is substantially longer than the carpus, not distinctly shorter.
Nothing is known on growth changes in proportions of these segments in P. brucei . The examined specimens from the Seychelles, however, seem to represent a different, possibly undescribed, species. Some of the males have well developed gonopods, with the endopod of the first pleopod bearing a distinct medial lobe, and the second pleopod with an appendix masculina. These just adult or subadult specimens do not differ noticeably from more juvenile ones. The leg segment length proportions are relatively stable in the series examined, and, thus, different from those described for P. brucei . The first pereiopod coxa in the Seychelles specimens bears a well produced distoventral setose process, which is not present in species of the P. granulimanus group (see Discussion), excluding P. tonga , but is present in the Periclimenes obscurus species group. We suspect that the Seychelles specimens belong, in all likelihood, to one of the species of the P. obscurus species group.
Color. No data; the species is known from only a single preserved specimen.
Host. Antipathes sp. (Cnidaria: Anthozoa: Antipatharia ), 52 m.
Distribution. Western Indian Ocean: Genego Is., North Nilandu Atoll, Maldives ( type locality).







No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Periclimenes laevimanus
| Ďuriš, Zdeněk 2010 |
Periclimenes
| Fransen, C. H. J. M. 1994: 119 |
Periclimenes brucei
| D uris, Z. 1990: 1 |