
Hemicyclopora, Norman, 1894
|
publication ID |
https://doi.org/ 10.5252/zoosystema2023v45a10 |
|
publication LSID |
urn:lsid:zoobank.org:pub:370E4D0A-FF10-4CAC-AF9F-A1A866FC1BEB |
|
DOI |
https://doi.org/10.5281/zenodo.8056983 |
|
persistent identifier |
https://treatment.plazi.org/id/03FBDF4F-9D03-FF97-D549-F29CCE5FBB75 |
|
treatment provided by |
Felipe |
|
scientific name |
Hemicyclopora |
| status |
|
“ Hemicyclopora ” pytheasi n. sp.
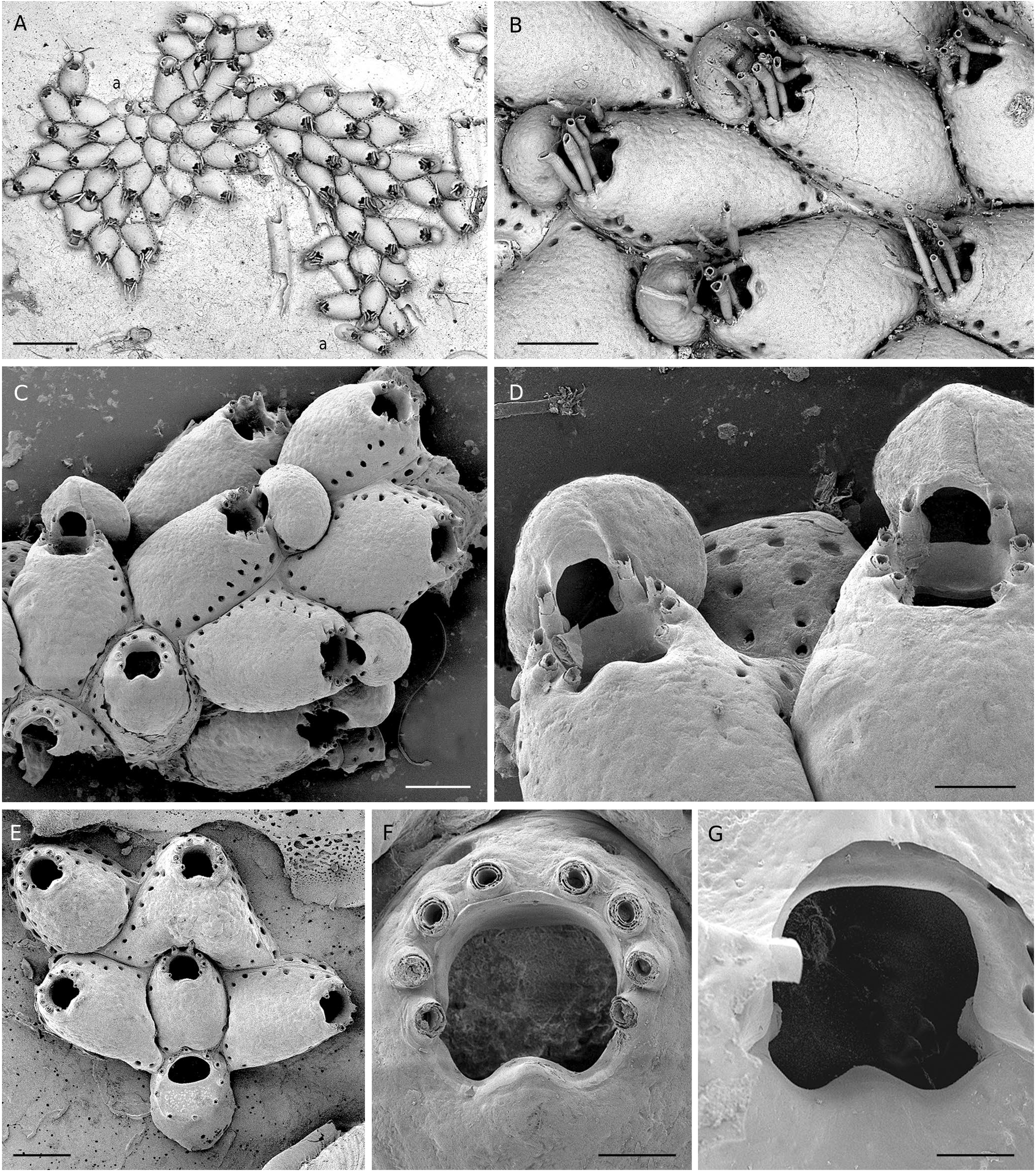
( Fig. 9 View FIG A-G; Tables 1 View TABLE ; 3 View TABLE ; 4 View TABLE )
urn:lsid:zoobank.org:act:5A983DF5-F216-42D8-A4EF-3721F1663EF7
? Hemicyclopora multispinata – Hayward & Ryland 1999, fig. 46D.
TYPE LOCALITY. — La Chapelle Bank, Armorican margin.
TYPE MATERIAL. — Holotype. NE Atlantic. France • 1 large colony, c. 80 zooids with a great proportion of ovicells (> 50%) and large interzoecial kenozooids; R/V Jean Charcot; Biaçores Stn 259; off Brittany, La Chapelle Bank ; 48°01.3’N, 7°51.5’W; 190 m depth; 19.XI.1971; on shell; Dre; H. Zibrowius leg.; MNHN-IB-2017-770 GoogleMaps .
Paratypes. NE Atlantic. France • 1 large colony, c. 60 autozooids (18 ovicells) + 2 ancestrulae budding 1 and 3 daughter zooids; R / V Jean Charcot; Biaçores Stn 259, off Brittany, La Chapelle Bank ; 48°01.3’N, 7°51.5’W; 190 m depth; 19.XI.1971; on shell; Dre; H. Zibrowius leg.; PMC. B33.5.5.2021 GoogleMaps • coated fragment of colony and detached orifices, same origin as holotype; MNHN-IB-2017-775 • 3 ovicellate colonies; JPL-Stn G61 (710), Bay of Biscay , 44°54’N, 02°11’W; 200 m depth; III.1967; on pebble; Dre; J. P. Lagardère leg.; MNHN-IB-2017-776 GoogleMaps • juvenile colony with ancestrula, coated, same origin as holotype; MNHN-IB-2017-777.
OTHER MATERIAL EXAMINED. — NE Atlantic. France • 1 colony; R / V Jean Charcot; Biaçores Stn 257, 47°57’N, 07°51.5’W; 335 m depth; 19.XI.1971; on Madrepora oculata ; Dre, H. Zibrowius leg.; MNHN GoogleMaps • 1 colony; R / V Jean Charcot; Biaçores Stn 259; same data as holotype and paratype PMC. B33.5.5.2021; on bivalve shell GoogleMaps .
ETYMOLOGY. — In honour of Pytheas, famous antique astronomer and sailor, citizen of Massalia (ancient Greek name of Marseille), who explored the northern seas during the 4th century B.C., and in reference to the geographical distribution of this species.
DIAGNOSIS. — Autozooids separated by deep grooves, frontal shield convex with mamillate texture, marginal pores large. Distal wall subvertical. Orifice terminal, condyles indistinct and relatively square, without lyrula-like protuberance; proximal edge of secondary orifice clearly convex with round top. Oral spines eight in both ovicellate and non-ovicellate zooids; spines of the proximalmost pair clearly converging; in ovicellate zooids, bases of the distal pair adjoining the ovicell. Ovicell apparently acleithral, attached to distal wall of maternal zooid, terminal and produced by a small, basal kenozooid; endooecium smooth, without proximal thickening. Large, interzooidal, porous kenozooids occasionally present. Ancestrula with opesia, cryptocyst and gymnocyst equally extended, five spines around the cryptocyst, 4-6 spines around the opesia.
DESCRIPTION
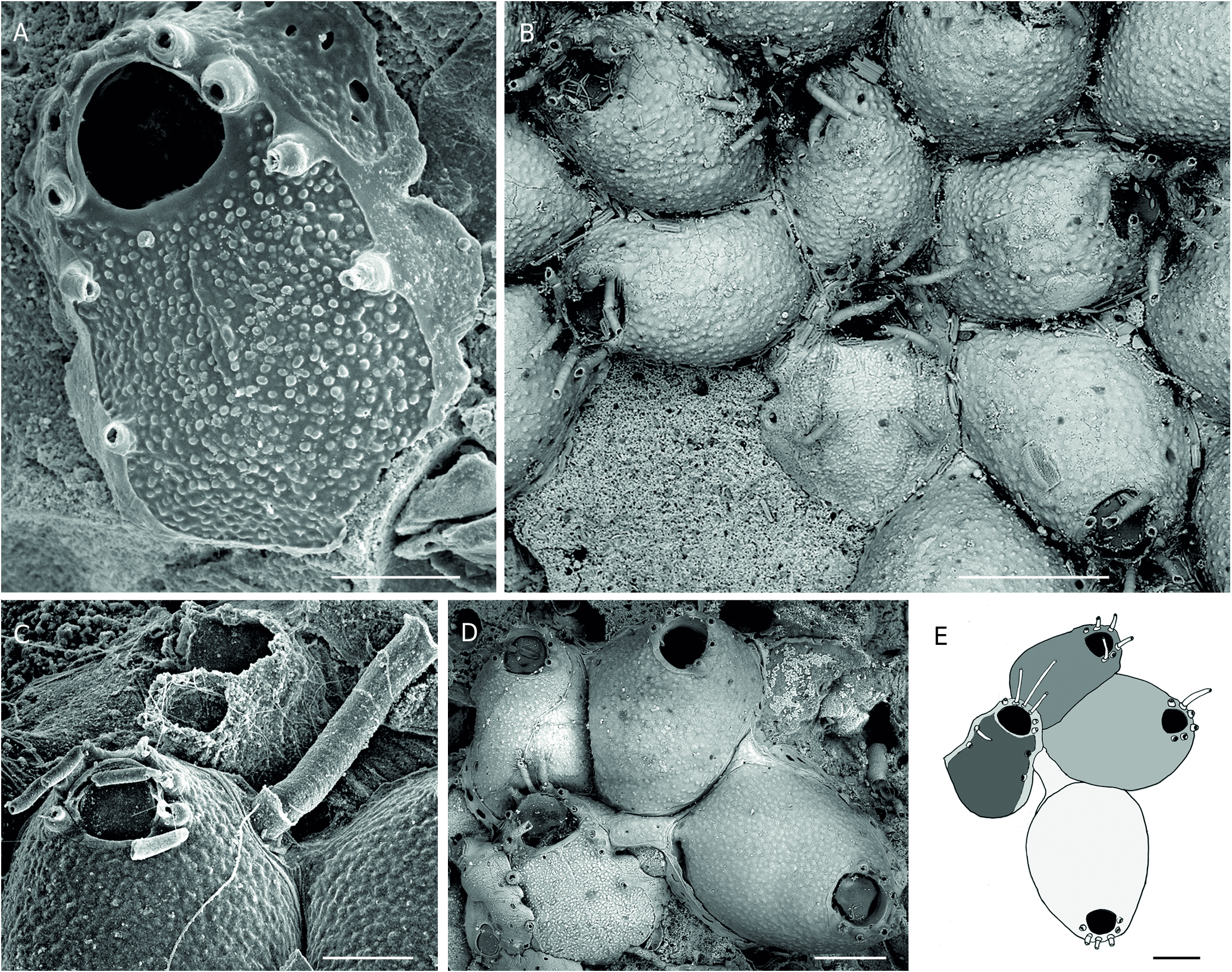
Colony encrusting, unilaminar, small to medium-sized. Autozooids large, elongated (L/W ratio = 1.48), distinctly separated by deep grooves, laid out in quincunx; frontal shield convex, smooth, slightly mamillated; marginal pores large (15-30 µm), distally arranged in a double or triple row and proximally in a single row ( Fig. 9 View FIG A-C). Distal wall subvertical ( Fig. 9B, C, E View FIG ), its inner side visible in ovicellate zooids as a broad, vertical frame below the ovicell opening and between the distalmost pair of spines ( Fig. 9D View FIG ). Orifice distal; primary orifice as long as wide or slightly longer, its calcified ring with step-shaped proximal ends, i.e., without prominent condyles; inner proximal side smooth and without any protuberance ( Fig. 9G View FIG ); proximal edge of secondary orifice clearly convex, with round top and without umbo ( Fig.9 View FIG B-D, F). Oral spines eight in ovicellate and non-ovicellate zooids, long (200-250 µm), huddled together, with thick, prominent bases, the proximalmost pair clearly convergent ( Fig. 9D, F View FIG ). Ovicells frequent, globular, sometimes remarkably wide, with surface structure of endooecium similar to that of frontal shield, without proximal protuberance, seemingly not closed by the operculum, attached to the vertical distal wall of the maternal zooid, apparently terminal and free ( Ostrovsky 2013: 125, 129), but produced by a small, poorly visible, basal kenozooid at the colony margin ( Fig. 9B, C View FIG ). Pore chambers numerous. Large interzooidal kenozooids occasional, with porous frontal shield ( Fig. 9D View FIG ). Ancestrula with proximal rim of opesia concave, proximal gymnocyst broad, five spines at the cryptocyst periphery, and four, five or six spines bordering the opesia ( Fig. 9E View FIG ).
REMARKS
Morphological features
The morphological features of “ H.” pytheasi n. sp. are very similar to those of E. similis and “ E.” massiliana n. sp., except for the structure of the primary orifice. Unlike other species of the same species complex, the internal side of the convex proximal edge of the orifice of “ H.” pytheasi n. sp. is smooth, without any bulge that could be identified as a lyrula ( Fig. 9G View FIG ). In the absence of such structure, does the convex edge of the orifice play the function of a lyrula and can stop the open operculum in a vertical position? The respective position of the maternal orifice and the ovicell suggests that the latter is acleithral ( Fig. 9C, D View FIG ), as in typical Escharella species. However, this feature remains incertain without specific anatomical investigation. Therefore, pending a molecular approach, this species was placed in the genus Hemicyclopora in quotation marks. Colonies of “ H”. pytheasi n. sp. often have a great proportion of ovicellate zooids, e.g.> 50% in the holotype specimen, and even more in two other colonies (Stn JPL-G61: 59%; Biaçores Stn 259: 74%). This high fertility was observed in most colonies (mean: 39 ± 24%) and likely has a specific significance. The occurrence of large interzoecial kenozooids ( Fig. 12D View FIG ) has been only observed in specimens from the Armorican Margin (Biaçores st. 259).
Taxonomic issues
“ Hemicyclopora ” pytheasi n. sp. shows obvious morphological similarities with “ H.” celtica n. sp. with which it can coexist along the Armorican margin. These species share the same shape of the secondary orifice and the presence of eight spines in non-ovicellate zooids. However, “ H.” pytheasi n. sp. differs in the external aspect of the frontal shield and endooecium (mamillated vs granular), the size of the marginal pores (large vs tiny), the condyles (step-shaped vs protuberant), and the ovicellate zooids. The latter differ in the number of spines in ovicellate zooids (eight in “ H.” pytheasi n. sp. vs six in “ H.” celtica n. sp.), and the building of ovicells, which involves a basal kenozooid (“ H.” pytheasi n. sp.), instead of a distal autozooid (“ H.” celtica n. sp.) ( Table 3 View TABLE ). The co-occurrence of these two species in the same depth zone of the same geographical area (Armorican margin) confirms the validity of these observed differences for discriminating them at species level. The specimen figured without indication of origin by Hayward & Ryland (1999: fig. 46D) and attributed to H. multispinata shows some typical features of “ H.” pytheasi n. sp.: proximal edge of the orifice typically convex, ovicellate zooid with eight spines, and large marginal pores. On the other hand, the drawing illustrating H. multispinata by the same authors ( Hayward & Ryland 1999: fig. 49) might be attributed as well to “ H.” pytheasi n. sp. or to “ H.” celtica n. sp. because of the lack of information concerning the ovicellate zooids and the shape of condyles.
HABITAT DISTRIBUTION
Colonies of “ H.” pytheasi n. sp. encrusted small substrates (empty shells, fragmented skeletons of Madrepora oculata , pebbles) at the edge of the continental shelf (190-335 m).
GEOGRAPHICAL DISTRIBUTION
The examined material came from the Armorican margin and the Bay of Biscay.
| R |
Departamento de Geologia, Universidad de Chile |
| V |
Royal British Columbia Museum - Herbarium |
| MNHN |
Museum National d'Histoire Naturelle |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SuperFamily |
Lepralielloidea |
|
Family |