
Arganthomyza socculata ( Zetterstedt, 1847 )
|
publication ID |
https://doi.org/ 10.5281/zenodo.4272829 |
|
publication LSID |
urn:lsid:zoobank.org:pub:E95E58A5-E0F1-4237-9D7C-4A81BB3120DD |
|
DOI |
https://doi.org/10.5281/zenodo.4339708 |
|
persistent identifier |
https://treatment.plazi.org/id/03FB87A9-FF80-FFEE-FEA8-6AE1FD42F9F4 |
|
treatment provided by |
Felipe |
|
scientific name |
Arganthomyza socculata ( Zetterstedt, 1847 ) |
| status |
|
Arganthomyza socculata ( Zetterstedt, 1847) View in CoL
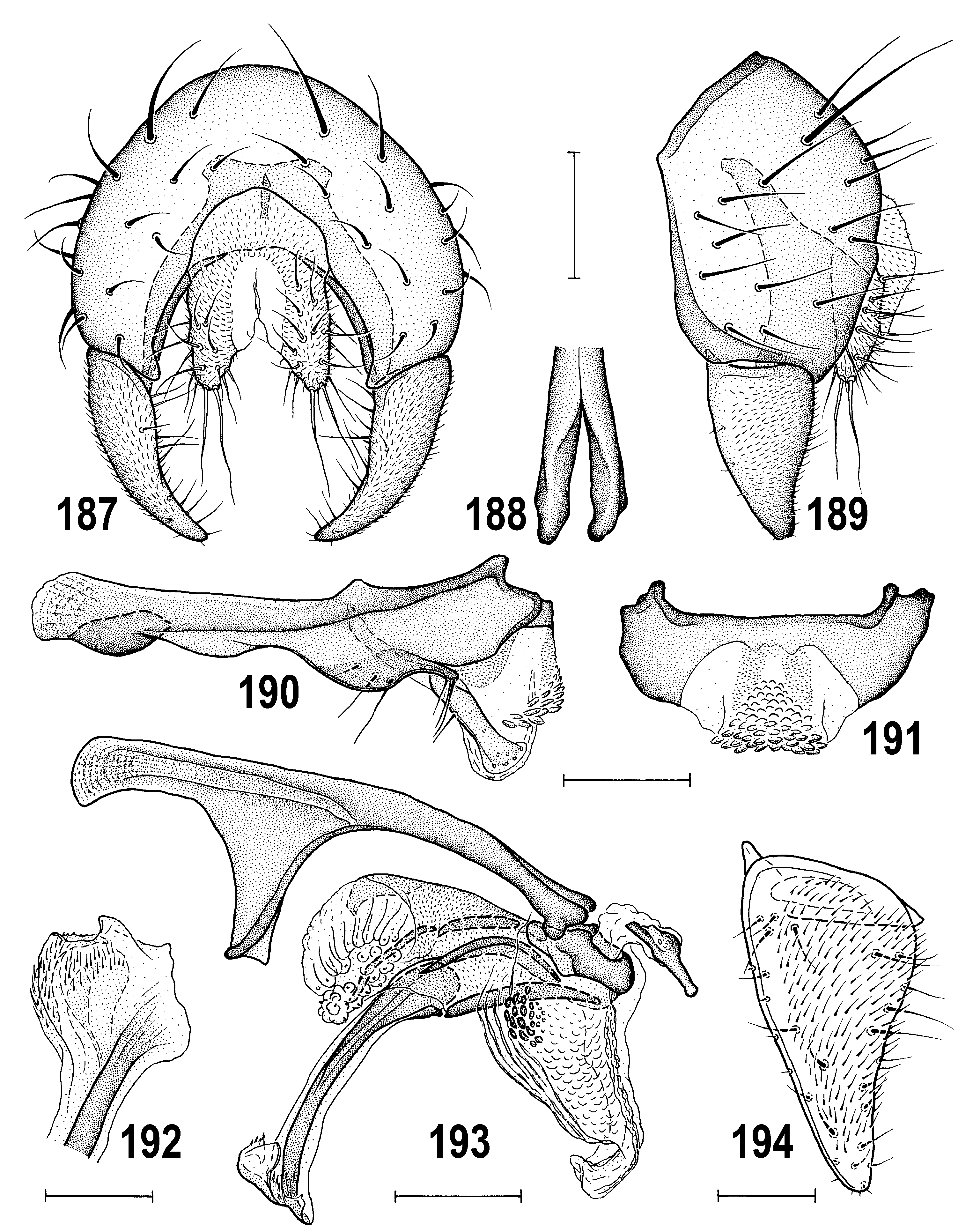
( Figs 185–202 View Figs 184–186 View Figs 187–194 View Figs 195–202 )
Geomyza socculata Zetterstedt, 1847: 2534 View in CoL .
Anthomyza socculata: ANDERSSON (1976) : 50 –51 (redescription, generic combination); SOÓS (1981): 111 (key); NDERSSON (1984b): 53 (catalogue); ROHÁĆEK (1984a): 393 (key), ROHÁĆEK (1987a): 55 –57 (dia gnosis), ROHÁĆEK (1998a): 173 (world checklist); ROHÁĆEK (2006a): 191 –196 (redescription and synonymy).
Arganthomyza socculata: ROHÁĆEK (2009a) View in CoL : 63, 74 (key, generic combination); ROHÁĆEK & BARBER (2013): 39 View Cited Treatment –41 (redescription, relationship).
Anthomyza ungulata Loew, 1873: 301 View in CoL ; CZERNY (1902): 252; CZERNY (1928): 5; COLLIN (1944): 267 (key); TROJAN (1962):39; STACKELBERG (1970):326 (key); ANDERSSON (1984b): 53 (catalogue); ROHÁĆEK (1984a):392 (synonymy).
Type material. Geomyza socculata Zetterstedt : HOLOTYPE: ♂, “ G.socculata ♂. Ostrog.Stenh.”, “ Lectotypus Geomyza socculata Zett. , design. 1974 H. Andersson ” [red label], “1969, 925” [green label], “Type No. 2095:1, Zool. Mus. Lund Sweden, Anthomyzidae ”, “ Holotypus ♂ Geomyza socculata Zett., J. Roháček des. 2005” [red label] and “ Anthomyza socculata (Zett., 1847) ♂, J. Roháček det. 2005” ( MZLU, genit. prep., examined). This type specimen was incorrectly designated as lectotype by ANDERSSON (1976: 45) but it is a holotype because only this male from Östergötland: Häradshammar , collected by Stenhammar, was listed in the original description.
Anthomyza ungulata Loew : LECTOTYPE: ♂ (designated by ROHÁĆEK 1984a: 392): “Kultuk, v. Maak”, “Coll. H. Loew”, “ Anthomyza ungulata Lw ” (Loew’s handwriting), “Type” [orange red label], “Zool. Mus. Berlin ” [yellow label], “ Lectotypus, Anthomyza ungulata Loew ♂, J. Roháček des. 1983” [label with red margin and the word “ Lectotypus ”] and “ Anthomyza socculata (Zett.) ♂, J. Roháček det. 1983”. The specimen is in good condition ( ZMHB, genit. prep., examined). PARALECTOTYPES: 2 ♀♀, with the same labels as the lectotype except for Loew’s determination label ( ZMHB, examined).
Other material examined. Palaearctic specimens, 311 ♂♂ 232 ♀♀ (see ROHÁĆEK 2006a, 2009a).Nearctic specimens, 35 ♂♂ 42 ♀♀ ( CASC, CNCI, DEBU, SMOC, USNM) (details in ROHÁĆEK & BARBER 2013).
Diagnosis. Male 1.90–2.42 mm, female 1.98–2.94 mm. Brown to blackish brown, distinctly though rather sparsely pale grey microtomentose and relatively shining; face, parafacialia, gena, postgena, mouthparts, haltere entirely and frons, antennae, legs largely contrasting ochreous, yellow or whitish yellow. Frontal triangle reaching up to anterior third of frons. Mid and hind basitarsus without short thickened setae. T1 and T2 almost separate, only laterally fused. Wing hyaline ( Figs 185–186 View Figs 184–186 ). Reference to the genitalia is necessary to differentiate conndently this species from A. disjuncta and A. acuticuspis .
Male genitalia (see Figs 187–194 View Figs 187–194 for details). Epandrium ( Figs 187, 189 View Figs 187–194 ) dark brown, distinctly higher than long. Gonostylus ( Figs 187, 189, 194 View Figs 187–194 ) ochreous to yellow, nat, slightly bent medially, of elongately subtriangular shape, with tapered but narrowly rounded apex, largely micropubescent on outer side and setose mostly on inner side; its shape variable: in Nearctic and E. Palaearctic specimens more slender and apically more acute (see Fig. 194 View Figs 187–194 ) on average than in European specimens but always with anterior margin convex and posterior margin concave (in largest extension view). Hypandrium ( Fig. 190 View Figs 187–194 ) relatively robust, with anterior internal lobes small, ventrally (anterior to pregonite) more distinctly excavated than in A. disjuncta ; posterior wide parts of hypandrium fused with transandrium. Pregonite ( Fig. 190 View Figs 187–194 ) fused to hypandrium, low, slightly projecting, with 2–3 posterior (1–2 longer on a tubercle-like process) and 3 anterior (all internal) setae. Aedeagal part of folding apparatus with dark granulose tubercles on proximal part (covering larger area on right side than on left) and usual nne striae ( Fig. 193 View Figs 187–194 ).
Female postabdomen and genitalia (see Figs 195–202 View Figs 195–202 for details). T7 and S7 completely fused into dark brown ring-shaped tergosternum, Nearctic and E. Palaearctic specimens dorsally often with distinctly paler posterior and sometimes also anterior band ( Fig. 195 View Figs 195–202 ), ventrally usually with dark, transverse anteromedial stripe ( Fig. 201 View Figs 195–202 ) which is sometimes reduced, and with long setae at posterior margin. Anterior margin of original S7 sometimes with a pair of small appendages (cf. Fig. 201 View Figs 195–202 ). Ventral receptacle ( Fig. 202 View Figs 195–202 ) hyaline, slender, long, strongly curved, gradually tapering towards blunt apex (somewhat thicker and shorter in Nearctic specimens). Spermathecae (1+1) short-pyriform, with dark transversely striated surface and slender base provided with rosette of 5–6 characteristic bell-shaped appendages (see Figs 196, 200 View Figs 195–202 ) some of which can have doubled apex in Nearctic specimens; duct with cervix developed but weakly sclerotized.
Variability. This widespread Holarctic species displays remarkable variability in the postabdominal characters. The Nearctic specimens closely resemble those from eastern Asia ( Mongolia, Russia: Far East), and compared to European specimens they seem to be generally smaller. The male gonostylus is narrower and more acute, the female T7+S7 is often anteriorly and/ or posteriorly pale-margined, the female T8 is narrower, the ventral receptacle is thicker, and the spermathecae often have doubled bell-shaped appendages.
Discussion. Arganthomyza socculata is very closely allied to A. disjuncta and forms with it a distinctive sister pair distinguished by the apomorphies diagnosing the A. socculata group (see ROHÁĆEK & BARBER 2013, and above). Arganthomyza socculata and A. disjuncta are distinct in the female sex but their males are very similar even in the genitalic structures (for differences see the key and in the description and discussion under A. disjuncta ).
The geographical variability discussed above, particularly the differences between European and E. Palaearctic + Nearctic specimens, will need future study to evaluate the differences between the populations in these areas. Future molecular studies may help to elucidate this problem, but for the time being these populations are considered conspecinc. However, we do not rule out the possibility that the Nearctic populations of A. socculata (hitherto known only from Alaska) interbreed in Alaska with the sympatric and closely allied A. disjuncta , because a few males found in some localities there could not be assigned to one or the other species. However, we cannot be sure that A. socculata is really restricted to Alaska in North America because much of the boreal component of the Nearctic Region has not been sunciently sampled for Anthomyzidae .
Biology. Old World habitats are varied but a common thread appears to be graminoids (both grasses and sedges) in open areas, sometimes along water bodies but also under birch canopy (see ROHÁĆEK & BARBER 2013 for details). Unfortunately no data on habitat and/or plant association are available for the Nearctic (Alaska) specimens, except for one specimen swept in tundra and one from vegetation on the edge of a lake. The habitat is nonetheless assumed to be similar to that of A. disjuncta (see above), at least in Alaska, where the two species co-occur at a few sites. Adults were recorded here in 15 June to 4 August.
Distribution. A Boreo-alpine circumpolar species widespread in the northern belt of the Palaearctic Region ranging from Iceland to Kamchatka but also occurring in montane ranges of more southern latitudes. Its Palaearctic distribution is summarized by ROHÁĆEK (2006a, 2009a) as follows: Austria, Czech Republic ( Bohemia), Estonia, Finland, Great Britain ( Scotland), Iceland, Kazakhstan, Kirghizia, Lithuania, Mongolia, North Korea, Norway, Poland, Russia (Central and North European Territory, W. and E. Siberia, Far East), Slovakia, Sweden, Switzerland, Ukraine. It was recorded for the nrst time from the Nearctic Region from Alaska ( United States of America) by ROHÁĆEK & BARBER (2013) (see Table 2, Fig. 600 View Fig ).
Arganthomyza socculata / disjuncta
Male specimens not distinctly referable to either of the two species. UNITED STATES OF AMERICA: ALASKA: 10 ♂♂ (CNCI, USNM) (details in ROHÁĆEK & BARBER 2013).



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Arganthomyza socculata ( Zetterstedt, 1847 )
| Roháćek, Jindřich & Barber, Kevin N. 2016 |
Anthomyza socculata:
| ROHACEK J. 2006: 191 |
| ROHACEK J. 1998: 173 |
| ROHACEK J. 1987: 55 |
| ROHACEK J. 1984: 393 |
| ANDERSSON H. 1976: 50 |
Anthomyza ungulata
| ANDERSSON H. 1984: 53 |
| ROHACEK J. 1984: 392 |
| TROJAN P. 1962: 39 |
| COLLIN J. E. 1944: 267 |
| CZERNY L. 1928: 5 |
| CZERNY L. 1902: 252 |
| LOEW H. 1873: 301 |
Geomyza socculata
| ZETTERSTEDT J. W. 1847: 2534 |