
Thomasomys ucucha, VOSS, 2003
|
publication ID |
https://doi.org/10.1206/0003-0082(2003)421<0001:ANSOTR>2.0.CO;2 |
|
DOI |
https://doi.org/10.5281/zenodo.5818480 |
|
persistent identifier |
https://treatment.plazi.org/id/03FAB267-FFBF-FFC5-FF24-FF53DA07DBE6 |
|
treatment provided by |
Felipe (2021-05-03 18:53:22, last updated 2024-11-29 15:17:08) |
|
scientific name |
Thomasomys ucucha |
| status |
sp. nov. |
Thomasomys ucucha , new species
Figures 7–13 View Fig View Fig View Fig View Fig View Fig View Fig View Fig
TYPE MATERIAL: The holotype, UMMZ 155644 View Materials (skin, skull, and fluidpreserved carcass; original number RSV 660 ), is an adult male that I collected on 26 April 1980 at an elevation of 11,100 ft (3384 m) in the valley of the Río Papallacta (ca. 3–5 km by trail NNW Papallacta), Provincia Napo, Ecuador .
Fortytwo other specimens collected in 1978 and 1980, hereby designated as paratypes, are from 8.2 km (by road) W Papallacta, 12,200 ft ( UMMZ 127119 View Materials , 127120 View Materials , 155717 View Materials ) ; 7.5 km (by road) W Papallacta, 12,000 ft ( UMMZ 155652–155655 View Materials , 155722– 155732 View Materials ) ; 6.2 km (by road) W Papallacta, 11,700 ft ( AMNH 244611–244613 About AMNH ; UMMZ 127121 View Materials , 155742 View Materials , 155649–155651 View Materials , 155714– 155716 View Materials , 155718 View Materials , 155719 View Materials , 155733–155736 View Materials ) ; and the Río Papallacta valley [3–5 km by trail NNW Papallacta], 11,100 ft ( UMMZ 155643 View Materials , 155645–155648 View Materials , 155720 View Materials , 155721 View Materials ) . Three additional paratypes ( AMNH 46621 About AMNH , 46622 About AMNH , 46624 About AMNH ) View Materials were collected in 1903 by L. Söderström at Tablón, in Provincia Pichincha (see appendix 1) .
DISTRIBUTION: Known only from the crest of the Cordillera Oriental (between ca. 3400 and 3700 m) just south of the equator in the Ecuadorean provinces of Pichincha and Napo.
ETYMOLOGY: Ucucha is the local Quichua word for ‘‘mouse’’ ( Orr, 1978), here treated as a noun standing in aposition to the generic name.
DIAGNOSIS: A mediumsized, darkfurred, longtailed species of Thomasomys with short, blunt rostrum; narrow interorbital region with rounded supraorbital margins; widely flaring zygomatic arches; straight frontonasal profile; broad, vertically oriented zygomatic plate; short incisive foramina; separate buccinatormasticatory and accessory oval foramina; primitive (pattern 1) carotid circulation; small, uninflated auditory bullae; small, hypsodont molars lacking welldeveloped cingula and stylar cusps; very small upper third molars; broad and conspicuously procumbent upper incisors; and a distinctive range of morphometric variation ( table 1).
DESCRIPTION: Pelage dense, fine, and soft, about 13–15 mm long over the back and rump; somberly colored (dark) and not abruptly countershaded. Masseffect dorsal coloration near Smithe’s (1975) Brownish Olive (color 29) along flanks, shading to Dark Grayish Brown (color 20) middorsally. Ventral pelage Dark Neutral Gray (color 83) basally, with superficial wash of Light Neutral Gray (color 85) or Glaucous (color 80); not sharply set off from dorsal coloration. Mystacial vibrissae long, extending just behind pinnae when laid back alongside head. Ears sparsely covered with short, blackish hairs, not contrasting conspicuously with color of head. Hairs over metapodials and digits of manus and pes dark, but tufts of longer hairs at bases of pedal claws silvery. Pes neither very narrow nor conspicuously broad; digit V long (its claw extending almost to base of claw of digit IV), but apparently nonopposable. Tail substantially longer than combined length of head and body, uniformly dark in most specimens but occasionally tipped with white; sparsely haired except for 5–10 mm terminal pencil. Mammae six in inguinal, abdominal, and postaxial pairs.
Skull (in dorsal view) characterized by short, blunt rostrum flanked by shallow zygomatic notches; narrow, hourglassshaped interorbit with rounded (not beaded or squared) margins; broadly flaring zygomatic arches; and large, oblong braincase unmarked by prominent temporal scars or lambdoidal ridges. Dorsal profile (in lateral view) distinctively flattened from nasal tips to midfrontal region; anterior margin of zygomatic plate straight and nearly vertical, not conspicuously sloping backward from base. Incisive foramina widest just behind premaxillary/maxillary suture and short (averaging 60% of diastemal length), not approaching first molar alveoli. Palatal bridge broad, smooth (without prominent ridges or grooves), and short (not extending posteriorly behind molar rows); posterolateral pits small, simple, inconspicuous perforations (never large, complex, or recessed in shallow fossae). Mesopterygoid fossa broad, straightsided, extending anteriorly between third molars; bony roof complete or perforated by narrow, slitlike sphenopalatine openings flanking the presphenoid/basisphenoid suture. Parapterygoid fossae narrow, approximately triangular, with shallow (unexcavated) anterior limits. Alisphenoid strut present, separating buccinatormasticatory from accessory oval foramina. Carotid circulation primitive (pattern 1), as indicated by large stapedial foramen, prominent squamosalalisphenoid groove, and sphenofrontal foramen. Postglenoid foramen and subsquamosal fenestra subequal; tegmen tympani broadly overlaps posterior suspensory process of squamosal. Auditory bullae small, uninflated, flaskshaped; without sharply defined transition between capsular part and bony eustacian tube. Tympanic membrane pars flaccida present, large. Malleus with large orbicular apophysis.
Mandible with distinct capsular process for lower incisor alveolus on lateral surface posteroventral to base of coronoid process. Basihyal moreorless straight (not strongly arched), without entoglossal process.
Upper incisors large, broad, and conspicuously procumbent, with heavily pigmented enamel bands (near Smithe’s [1975] Spectrum Orange [color 17] in fresh material). Upper molars in parallel left and right series, small, pentalophodont, hypsodont when unworn (by comparison with more brachydont congeners), and lacking welldeveloped cingula and stylar cusps. M1 anterocone divided by anteromedian flexus into subequal anterolabial and anterolingual conules. Paralophs and metalophs (on M1 and M2) connect corresponding labial cusps to mesolophs and posterolophs, respectively, or to median mures, not to opposing lingual cusps. M3 conspicuously smaller than M2 (<50% as estimated by occlusal areas) and usually lacking a distinct lingual fold (hypoflexus). Lower molars similar to upper teeth in general design, but m1 anteroconid often undivided (even in unworn dentitions), and m3 not conspicuously reduced. Molar root formulas unknown (no specimens are available with loose teeth), but M1 and m1 apparently without accessory rootlets.
Stomach unilocularhemiglandular. Adult males with one pair each of dorsal prostate, anterior prostate, ampullary, vesicular, and bulbourethral glands; and with two pairs of ventral prostate glands. Macroscopic preputial glands absent. Glans penis small, short, and subcylindrical (weakly divided into right and left halves by a shallow middorsal trough and an inconspicuous midventral raphe but otherwise unmarked by external folds); externally covered with coarse spines except for broad rim of soft, crenulated tissue surrounding terminal crater; crater contents include three bacular mounds, bifurcate urethral flap, and one dorsal papilla; two small spinous patches of rugose epithelium present dorsolateral to bacular mounds, but remaining crater contents unarmed.
COMPARISONS: As restricted by Voss (1993) and González (2000), the genus Thomasomys consists of 6mammate pentalophodont Andean sigmodontines with very shallow zygomatic notches; hourglassshaped interorbital regions that lack welldeveloped beads or projecting supraorbital shelves; short palates lacking prominent posterolateral pits; and auditory bullae firmly attached to the skull by overlap of the tegmen tympani with a posterior suspensory process of the squamosal. Because all of these traits are currently thought to be plesiomorphies within the Neotropical muroid radiation ( Voss, 1993), the genus lacks compelling evidence of monophyly. Pending a comprehensive phylogenetic analysis of the ‘‘thomasomyine’’ group, however, there is no more restricted taxonomic category within which to assess the relationships of T. ucucha . Phenetically, the new species most closely resembles T. hylophilus Osgood (1912) , an allopatric taxon that occurs in northeastern Colombia and western Venezuela. 3
Thomasomys ucucha and T. hylophilus overlap in all external and craniodental measurements ( table 1), and they share many qualitative traits in common: both lack genal vibrissae but have moderately long mystacial hairs; relatively long tails; similarly proportioned hind feet; flattened frontonasal profiles; narrow interorbital regions with rounded supraorbital margins; straight, vertically oriented zygomatic plates; broad palates; alisphenoid struts that separate buccinatormasticatory and accessory oval foramina; complete (pattern 1) carotid arterial circulations; oblong braincases; small, uninflated auditory bullae; relatively hypsodont molars that lack well developed cingula and stylar cusps; and unilocularhemiglandular stomachs. Despite this suite of resemblances, ucucha and hylophilus differ in other points of comparison.
In dorsal cranial view (fig. 7), Thomasomys ucucha is distinguishable at a glance by its relatively short, broad rostrum and by its widely flaring, rounded zygomatic arches. By contrast, the rostrum of T. hylophilus is proportionately longer and narrower, and the zygomatic arches converge anteriorly from a widest point across their squamosal roots. In ventral view, the incisive foramina of ucucha are absolutely shorter than those of hylophilus , and they are also proportionately shorter in relation to the diastema: the ratio LIF/LD averages about 61% in the former species versus about 76% in the latter. The posterior opening of the alisphenoid canal, a tiny perforation behind each parapterygoid fossa in ucucha , is a conspicuously larger orifice in hylophilus .
Thomasomys ucucha and T. hylophilus are also dentally distinctive. The upper incisors of ucucha are broader, more deeply pigmented, and more procumbent than those of hylophilus . In sidebyside comparisons, the contrast in upper incisor procumbency between the two species is visually obvious (fig. 7), but measurements provide a more objective basis for discrimination. Measured with an ocular goniometer, the chord that subtends the exposed greater curvature of these teeth defines an average anterior angle of 87° with the occlusal plane of the upper molars in ucucha (observed range, 85–89°; N = 14), whereas the homologous angle has an average value of 77° in hylophilus (observed range, 76–79°; N = 11). 4 A correlated species difference in the lower incisors is likewise apparent: whereas the lower incisor root of ucucha is contained in a prominent capsular process on the lateral mandibular surface just below the base of the coronoid process (fig. 8A), the lower incisor root of hylophilus terminates in an inconspicuous bony ridge without a distinct process (fig. 8B).
The molar dentition provides several additional traits of diagnostic value. The upper toothrow is shorter on average in Thomasomys ucucha than in T. hylophilus ( table 1), a difference that is primarily attributable to the size of M3. That tooth ranges from 0.8 to 1.0 mm long and accounts for just 21% of average toothrow length in ucucha , versus 1.2– 1.3 mm long and 25% of toothrow length in hylophilus . Correlated species differences in occlusal complexity are also apparent (fig. 9). In the upper molars, the anterolophs and mesolophs of M1 and M2 are much more weakly developed in ucucha than in hylophilus , and M3 usually lacks a distinct lingual fold (hypoflexus) in ucucha that is consistently present in hylophilus . In the lower dention, the anteromedian flexid of m1 is shallower, less persistent with wear, or altogether absent in ucucha , whereas this fold is always present and usually persistent in hylophilus .
Incisor procumbency alone is sufficient to set Thomasomys ucucha apart from other congeners, only two of which approach the orthodont condition (defined in footnote 4, above). However, both of those species— T. australis with an index value of 85°, and T. daphne with an index value of 90°—have chisellike incisors that are much shorter and narrower than the more scooplike, longer, and broader teeth of ucucha (fig. 10), and neither species closely resembles ucucha in other respects. Thomasomys ucucha is readily distinguished from other congeneric species known to occur in the Cordillera Oriental of northern Ecuador ( T. aureus , T. baeops , T. cinnameus , T. erro , T. paramorum , T. rhoadsi , T. silvestris ; see below) by numerous qualitative and quantitative character differences that are summarized in table 2.
REMARKS: In a previous list of Papallacta mammals ( Voss, 1988: table 43), I referred to this taxon as ’’ Thomasomys sp. ’’
FIELD OBSERVATIONS: The 42 specimens of Thomasomys ucucha that I collected near Papallacta in 1978 and 1980 were trapped at elevations ranging from 3380 to 3720 m. Of these, 3 were taken in grassy páramo, 22 were taken in the shrubby páramo/forest ecotone or in grassy glades surrounded by forest, and 17 were taken deep inside Subalpine Rain Forest. Most recorded captures were on the ground, of which 18 were in rabbit trails or runways through dense grass or low herbs; 16 were in runways through moss or damp litter, under mossy debris, or at the bases of mossy trees; and 7 were along the wet margins of small streams. Only one specimen was trapped off the ground, on the mossy limb of a low tree. Other muroid species that were trapped syntopically (in the same habitats) with T. ucucha include Akodon latebricola , Akodon mollis , Anotomys leander , Chilomys instans , Microryzomys altissimus , M. minutus , Neusticomys monticolus , T. aureus , T. baeops , T. cinnameus , T. erro , and T. paramorum .
Gonzalez, E. M. 2000. Un nuevo genero de roedor sigmodontino de Argentina y Brasil (Mammalia: Rodentia: Sigmodontinae). Comunicaciones Zoologicas del Museo de Historia Natural de Montevideo 196: 1 - 12.
Orr, C. 1978. Dialectos quichuas del Ecuador. Cuadernos etnolinguisticos no. 2. Quito: Instituto Linguistico de Verano.
Osgood, W. H. 1912. Mammals from western Venezuela and eastern Colombia. Field Museum of Natural History Zoological Series 10: unpaginated frontispiece map + 33 - 66.
Smithe, F. B. 1975. Naturalist's color guide. New York: American Museum of Natural History.
Thomas, O. 1919. The method of taking the in- cisive index in rodents. Annals and Magazine of Natural History 7 (17): 421 - 423.
Voss, R. S. 1988. Systematics and ecology of ichthyomyine rodents (Muroidea): patterns of morphological evolution in a small adaptive radiation. Bulletin of the American Museum of Natural History 188: 259 - 493.
Voss, R. S. 1993. A revision of the Brazilian muroid rodent genus Delomys with remarks on '' thomasomyine' ' characters. American Museum Novitates 3073: 1 - 44.
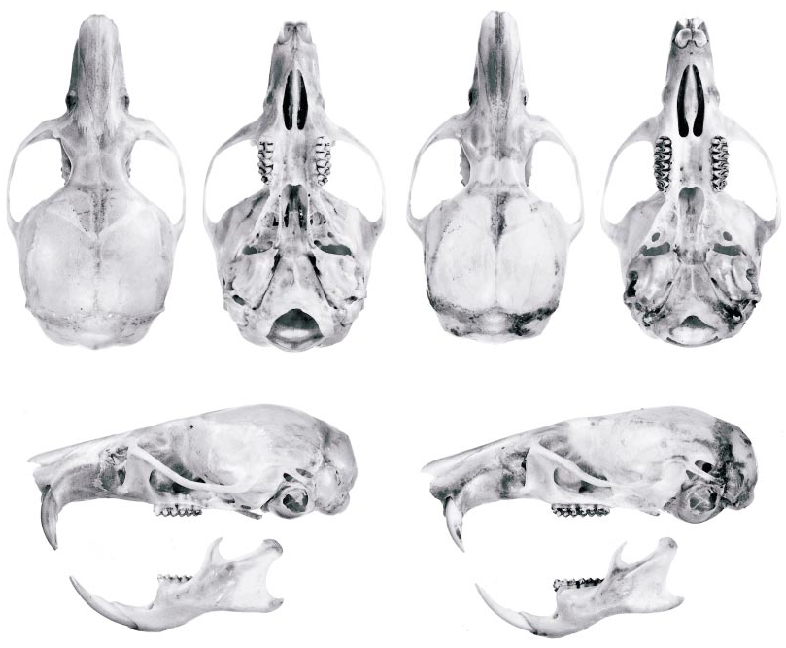
Fig. 7. Dorsal, ventral, and lateral cranial views of Thomasomys ucucha (left, UMMZ 155644, holotype) and T. hylophilus (right, FMNH 18593, paratype). Approximately twice life size.
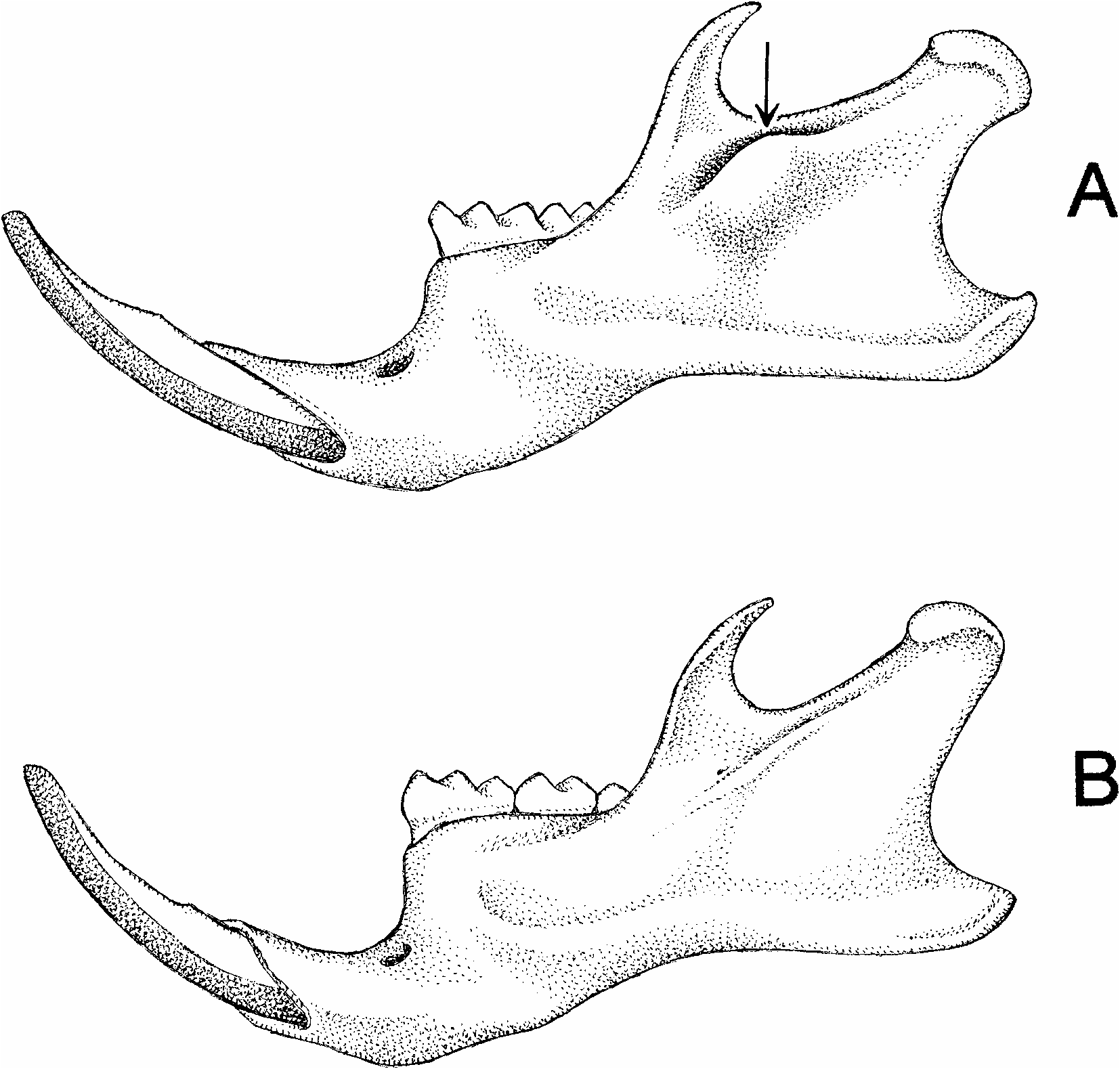
Fig. 8. Lateral mandibular views of Thomasomys ucucha (A, UMMZ 155644, holotype) and T. hylophilus (B, FMNH 18593, paratype). Among other species differences, a distinct capsular process for the lower incisor alveolus is present posteroventral to the coronoid process in T. ucucha (arrow), whereas this structure is absent in T. hylophilus.
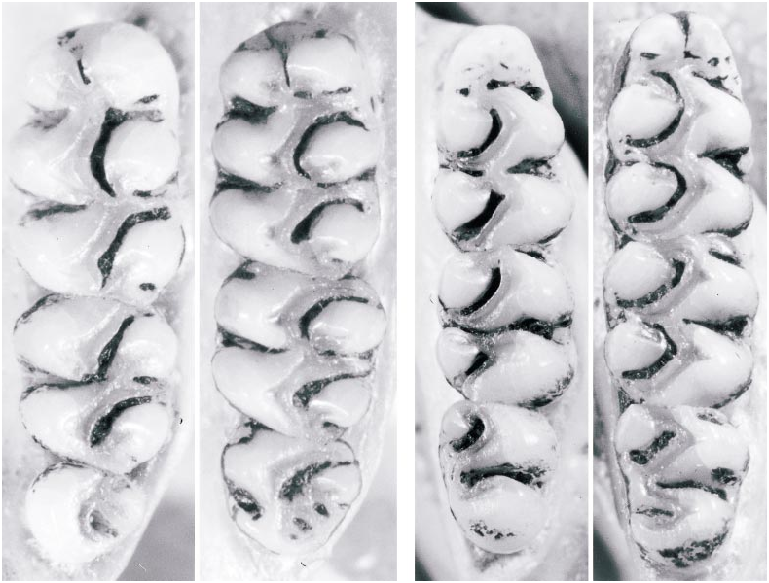
Fig. 9. Left upper and right lower molars of Thomasomys ucucha (lefthand photo of each pair, UMMZ 155722) and T. hylophilus (righthand photo of each pair, USNM 442322). Note the relatively smaller M3 of T. ucucha, the presence of a distinct lingual fold on the M3 of T. hylophilus, and the relatively larger (more robust) anterolophs and mesolophs of the latter species.
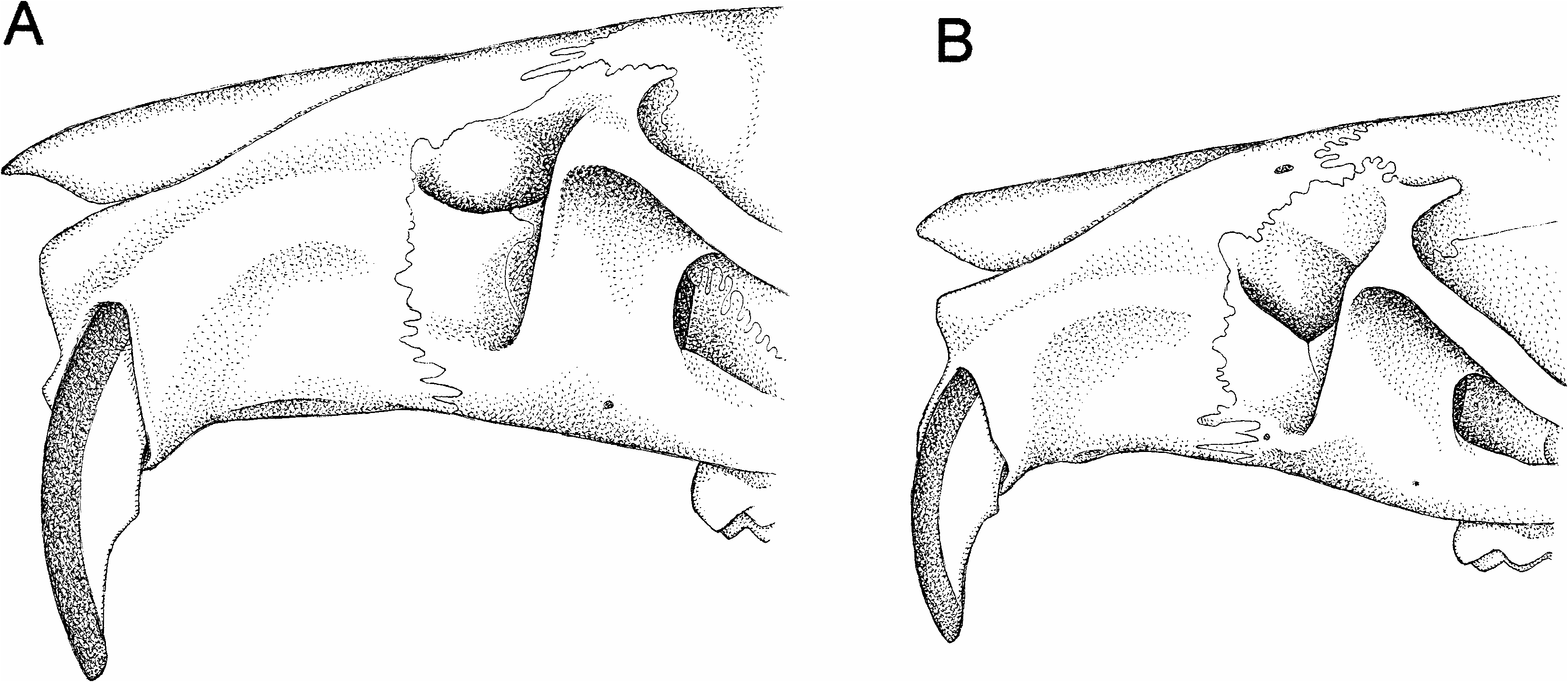
Fig. 10. Lateral view of rostrum of Thomasomys ucucha (A, UMMZ 155652) and T. daphne (B, UMMZ 160579). Although both species have procumbent (orthodont) incisors, those of ucucha are longer, broader, and more scooplike than the shorter, narrower, chisellike teeth of daphne.
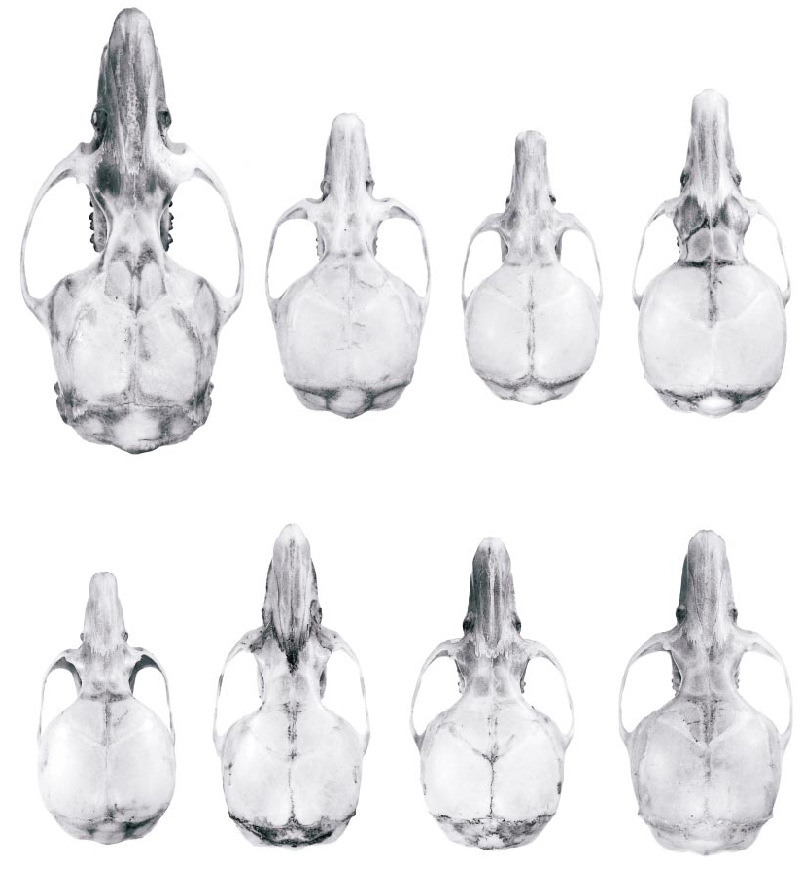
Fig. 11. Dorsal cranial views of eight Thomasomys species that occur in the Cordillera Oriental of northern Ecuador. Top row (left to right): T. aureus (UMMZ 127114), T. baeops (UMMZ 155708), T. cinnameus (UMMZ 155671), T. erro (UMMZ 155713). Bottom row (left to right): T. paramorum (UMMZ 155737), T. rhoadsi (AMNH 66256), T. silvestris (USNM 513592), T. ucucha (UMMZ 155644).
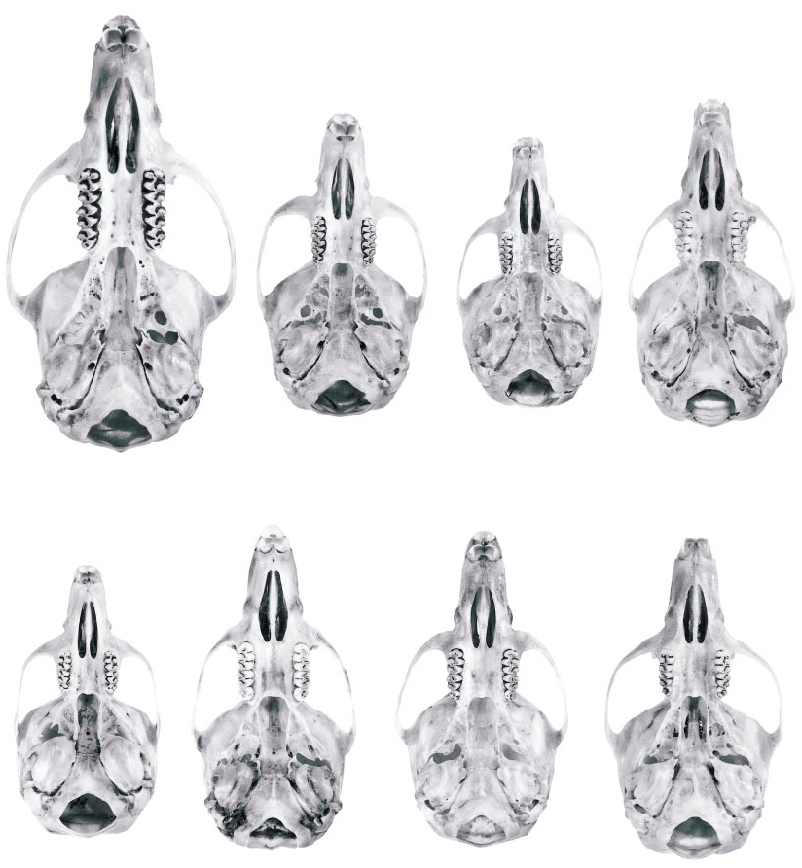
Fig. 12. Ventral cranial views of eight Thomasomys species from the Cordillera Oriental of northern Ecuador. Top row (left to right): T. aureus (UMMZ 127114), T. baeops (UMMZ 155708), T. cinnameus (UMMZ 155671), T. erro (UMMZ 155713). Bottom row (left to right): T. paramorum (UMMZ 155737), T. rhoadsi (AMNH 66256), T. silvestris (USNM 513592), T. ucucha (UMMZ 155644).

Fig. 13. Lateral cranial and mandibular views of eight Thomasomys species from the Cordillera Oriental of northern Ecuador. Left side (top to bottom): T. aureus (UMMZ 127114), T. baeops (UMMZ 155708), T. cinnameus (UMMZ 155671), T. erro (UMMZ 155713). Right column (top to bottom): T. paramorum (UMMZ 155737), T. rhoadsi (AMNH 66256), T. silvestris (USNM 513592), T. ucucha (UMMZ 155644).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
1 (by felipe, 2021-05-03 18:53:22)
2 (by tatiana, 2021-10-28 12:24:29)
3 (by ExternalLinkService, 2021-10-28 12:36:23)
4 (by ExternalLinkService, 2021-10-28 13:14:40)
5 (by tatiana, 2022-01-04 18:37:25)
6 (by ExternalLinkService, 2022-01-04 18:42:33)
7 (by ExternalLinkService, 2022-12-14 18:41:03)
8 (by plazi, 2023-11-02 08:04:52)
9 (by ExternalLinkService, 2023-11-02 15:15:31)
2022-12-14 16:35:59: BeatEstermann linked materials citation to GBIF Specimen Record URL: https://www.gbif.org/occurrence/859542216
2022-12-14 16:35:59: BeatEstermann linked materials citation to GBIF Specimen Record Key: 859542216