
May norm, Jäger & Krehenwinkel, 2015
|
publication ID |
https://doi.org/ 10.5733/afin.056.0209 |
|
persistent identifier |
https://treatment.plazi.org/id/03F987A3-ED4D-FFE4-357A-B013FCC90096 |
|
treatment provided by |
Felipe |
|
scientific name |
May norm |
| status |
sp. nov. |
May norm View in CoL sp. n.
Figs 78–99 View Figs 78–86 View Figs 87–93 View Figs 94–99
Etymology: This new species is named for Norman (“Norm”) Platnick, New York, for his eminent contributions to arachnological research and especially for his work on the most-used tool in arachnology, the World Spider Catalog; noun (name) in apposition.
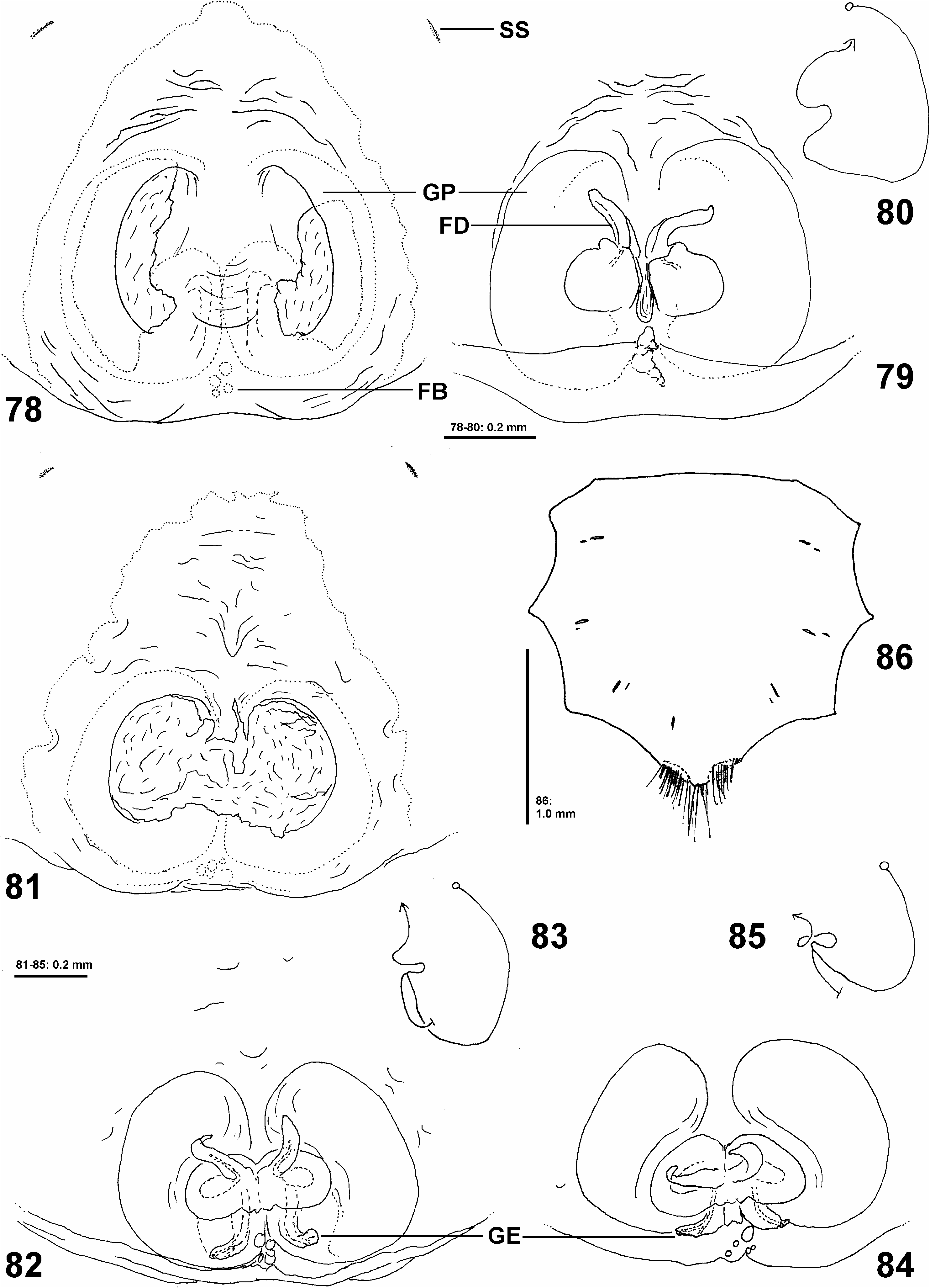
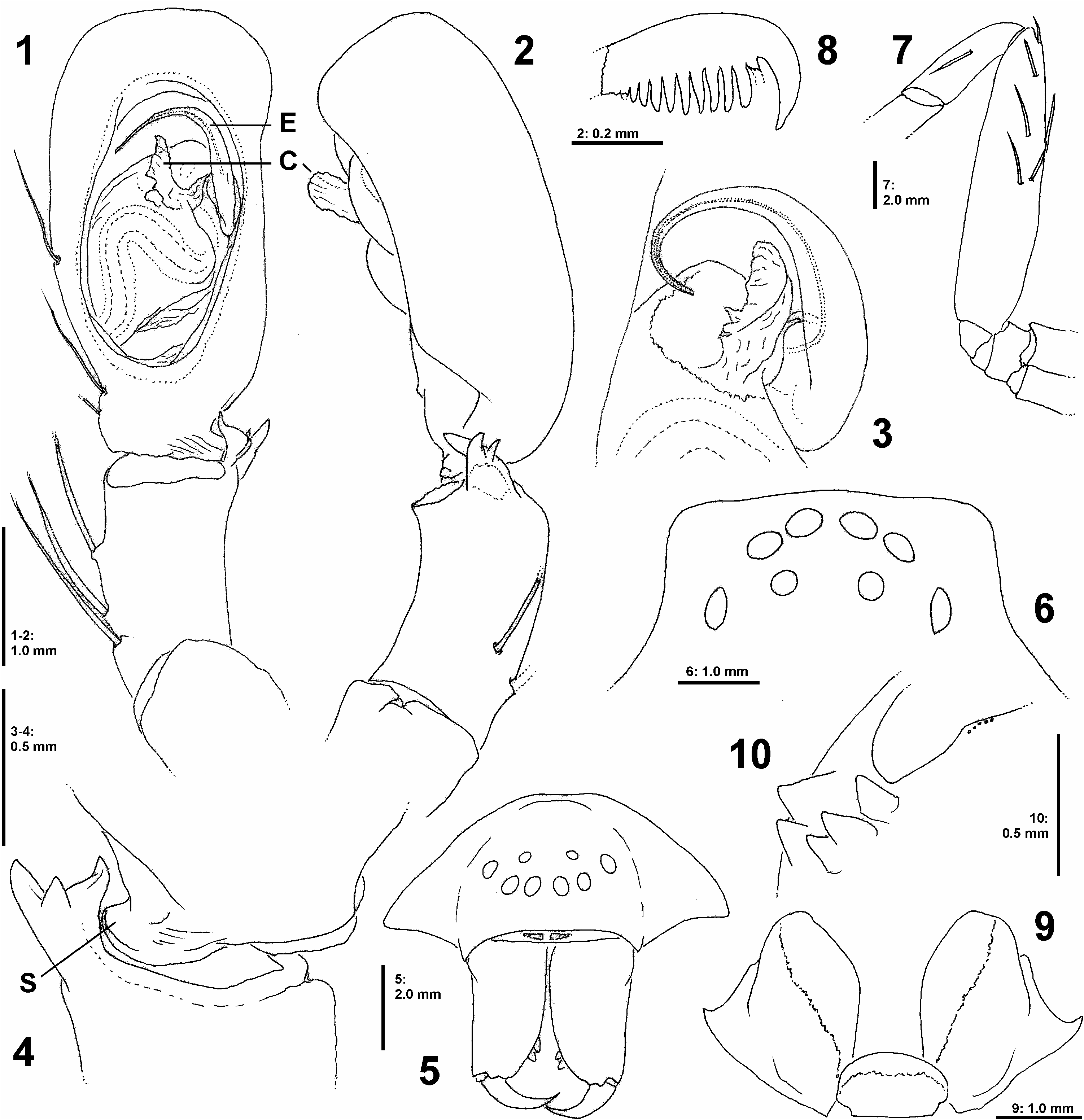
Diagnosis: Small to medium-sized Sparassidae of the African clade with body length of 8.1–11.6 in females. Females may be recognised by two bow-shaped lateral margins of the atrium ( Figs 78, 81 View Figs 78–86 ) and one pair of spherical structures in the internal duct system ( Figs 79, 82, 84 View Figs 78–86 ). Male unknown.
Description:
Female (holotype, PJ 1851).
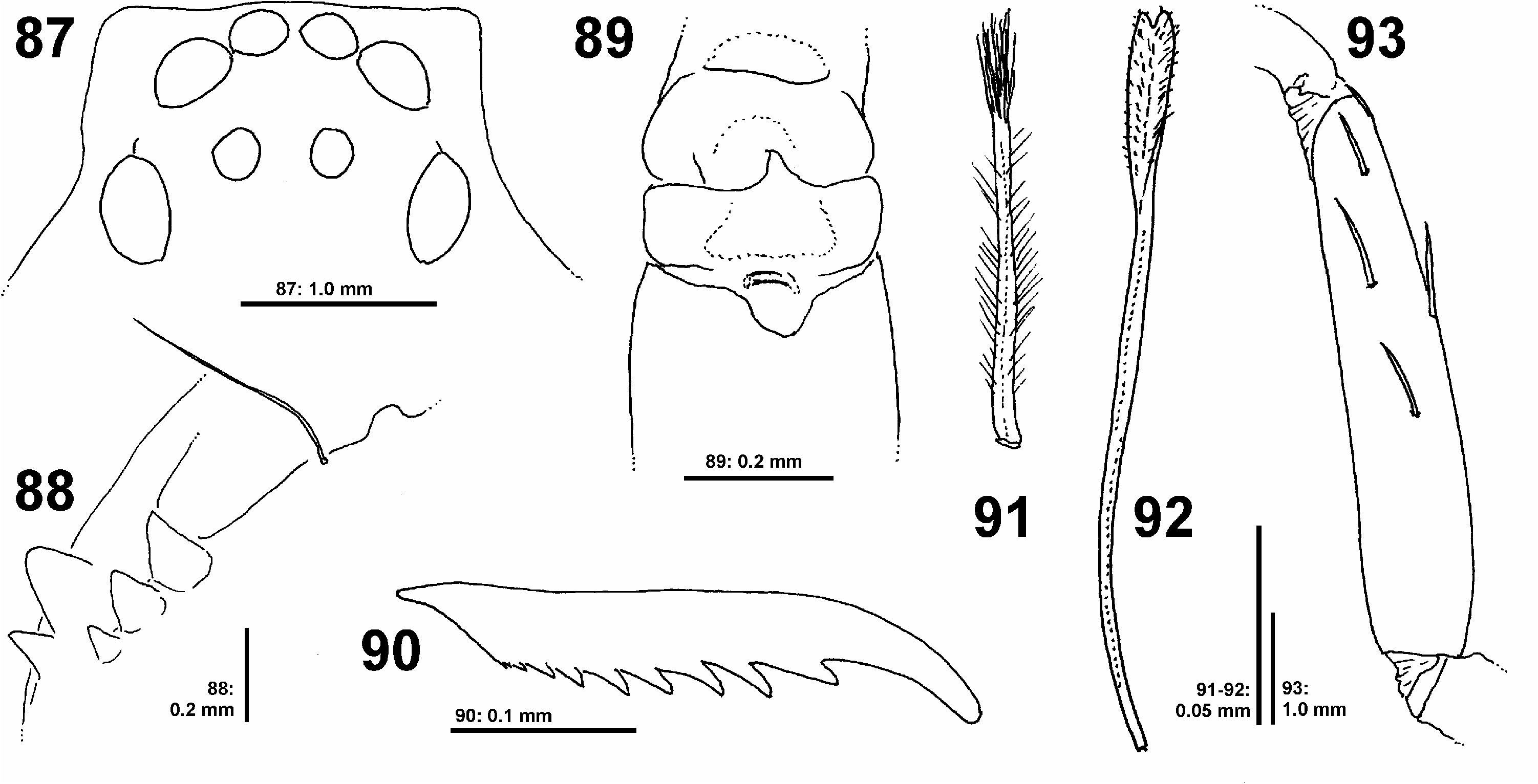
DS length 3.9, width 3.3, anterior width 2.0, OS length 4.5, width 2.8. Eyes ( Fig. 87 View Figs 87–93 ): AME 0.32, ALE 0.40, PME 0.20, PLE 0.60, AME–AME 0.05, AME–ALE 0.01, PME–PME 0.30, PME–PLE 0.29,AME–PME 0.34,ALE–PLE 0.21, clypeus height at AME 0.26, at ALE 0.31. Spination: palp: 131, 001, 0110, 0012; legs: femur I–III 323, IV 322; patella I 000, II–III 100, IV 101; tibia I 23(2)26, II–IV 2326; metatarsus I–III 3034, IV 3037. All metatarsi with moderately dense to sparse scopula, metatarsus IV ventrally with double row of bristles, continued with only few bristles in tarsus. Tarsus II ventrally with 6 claw slit sensilla (sensu RamÍrez 2014) in distal transverse suture. Leg formula: 3421. Measurements of palp and legs: palp 6.4 (1.9, 1.0, 1.4, -, 2.1), leg I 17.2 (4.9, 2.1, 4.6, 3.9, 1.7), leg II 18.7 (5.6, 2.2, 5.0, 4.1, 1.8), leg III 19.2 (5.5, 2.1, 4.8, 4.0, 1.8), leg IV 19.0 (6.0, 2.0, 5.0, 4.2, 1.8). Cheliceral furrow without denticles; promargin of chelicerae with 2 teeth, retromargin with 3 teeth ( Fig. 88 View Figs 87–93 ); with 1 bristle at fang base. Palpal claw with 10 teeth, claw of leg IV with 7 teeth.
Palp with two rows of long setae on femur, tibia and tarsus forming a basket. Sternum with 3 pairs of doublets of slit sense sensilla, each doublet consisting of a larger and a smaller sensillum (cf. Fig. 86 View Figs 78–86 ). Sternum ventrally and chelicerae frontally with long bristles. Gnathocoxae without serrula. Metatarsal stopper (sensu RamÍrez 2014) with lateral projections weakly developed, median hook well extending beyond lateral projections ( Fig. 89 View Figs 87–93 ).
Copulatory organ as in diagnosis ( Figs 78–85 View Figs 78–86 ): Epigynal field almost as wide as long, anteriorly converging, without slit sensilla, with fusion bubbles on posterior part. Internal duct system with semi-circular guiding pockets leading to pair of spherical structures (spermathecae?) and glandular extensions situated posteriorly; fertilisation ducts arising anteriorly from these structures, antero-laterad. Both sides of the epigyne were covered with mating plugs.
Coloration ( Figs 94–99 View Figs 94–99 ): Yellowish to reddish brown without distinct markings. White hairs around eyes, margin of prosoma, chelicerae and legs. DS with longitudinal fovea and slightly marked striae, centrally with dark setae, eye region with stiff bristles. OS with dark hairs especially anteriorly and around heart patch, otherwise with light short setae interspersed with dark setae. Spinnerets and ventral area in front of spinnerets with dark setae.
Male. Unknown.
Variation: Females (n= 5): DS length 3.8–5.6. OS length 4.3–6.0. Spination: Femur IV 322(1); patella I 101, II 101/000, III 100(1), IV 100. Palpal claw with 8 teeth. Tarsus II ventrally with 7 claw slit sensilla in distal transverse suture. Leg formula 4231 (n=3). One chelicerae of one female (PJ 3543) with 4 posterior teeth. Sternum with single additional slit sensillum in posterior part ( Fig. 86 View Figs 78–86 ). Epigynal field of two females exhibited no slit sensilla. In the epigyne of one female (PJ 3542) a mating plug was found only on the right side, in another female (PJ 3538) one mating plug stretched across the entire atrium, two more females showed two plugs, each on one side.
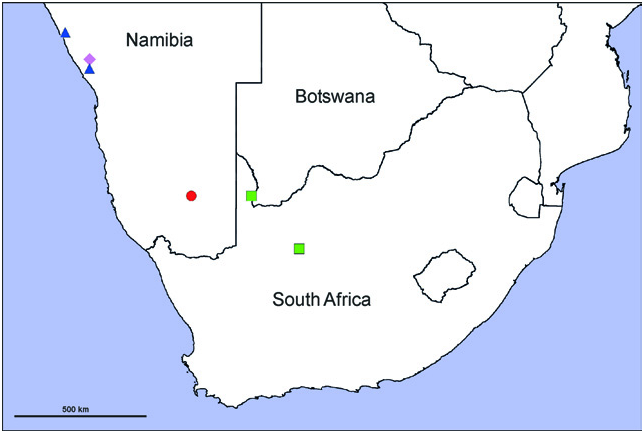
Holotype ♀ (PJ 1851): NAMIBIA: Erongo: Messum crater, 21°26.430'S 14°13.215'E, 420 m, E. & M. Griffin leg., 5.iv.2000 ( SMN 44693 ). GoogleMaps
Paratypes: 1♀ (PJ 1852), same data as holotype ( SMF) GoogleMaps ; 1♀ (PJ 1853), same data as holotype ( SMF) GoogleMaps ; 1♀ (PJ 3538), same data as holotype, on ground at night, gravel plain, M. Griffin leg., SD 1211 ( SMN 44723 ) GoogleMaps . 1♀ (PJ 3543), same data as holotype, on ground, at night, gravel plain, near rocky hillside, E. Griffin leg., 6.iv.2000 ( SMN 44688 ) GoogleMaps .
Other material examined: NAMIBIA: Kunene: 1♀ (PJ 3542), Skeleton Coast Park , 20°00.407'S 13°15.932'E [310 m], on ground at night, M. Griffin leg., 21.vi.2000 ( SMN 44700 ) GoogleMaps . Erongo: 3 juveniles, Damaraland, Brandberg, 21°26.430'S 14°13.215'E, M. Griffin leg., iv.1986 ( SMN 44415 ) GoogleMaps .
Distribution: Known from three localities in northern Namibia (Kunene, Erongo regions) ( Fig. 120 View Fig ).
Biology:According to information on the labels, spiders are nocturnal and active on the ground. Similarly to M. bruno sp. n., females of M. norm sp. n. exhibited scars, which may be an indicator that males bite the females during courtship. In the holotype female six such scars were located dorsally on right tibia II, one large scar ventro-proximally on the left palpal tibia, in the female paratype (PJ 3542) one scar each was located on left femur I prolaterally and on left coxa IV ventrally. One female (PJ 3538) had no scars, but a mating plug.
Note: Since this new species occurs in the same geographical range as the previous one and is only known from the female sex, whereas M. rudy sp. n. is described from the male sex only, both forms could be theoretically considered conspecific. However, considerable somatic differences, like the distinctly larger PLE, the considerably darker coloration with a higher number of dark setae, the (relatively and absolutely) much longer lateral spines on metatarsus IV, and the presence of a dense scopula on tarsus IV, with only few ventral bristles in M. norm sp. n., suggest that both forms in fact represent different species. Such strong differences between sexes of one species are not known from other Sparassidae nor from May bruno sp. n.
MOLECULAR RESULTS AND DISCUSSION
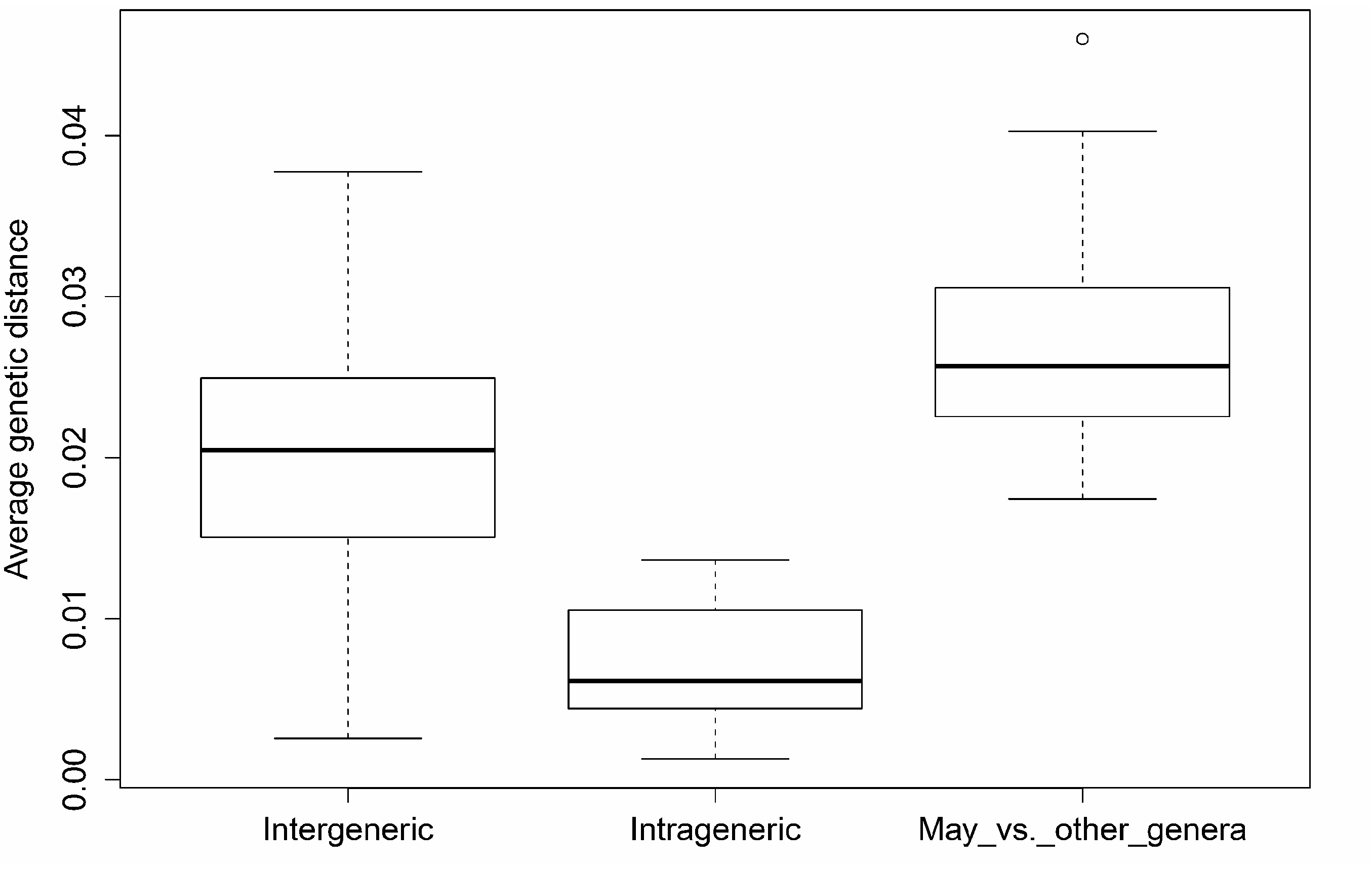
We analysed sequences from 124 Sparassidae specimens in 19 genera. However, many of these sequences were not unique. After pruning, 63 unique sequences in 19 genera remained. In addition, we included three sequences of May bruno gen. n. sp. n. Due to DNA degradation, we could not sequence other species of that genus. The intergeneric distances range from 0.26 % ( Palystella vs. Carparachne ) to 3.78 % ( Heteropoda Latreille, 1804 vs. Leucorchestris ). Within genera, we find an average distance of 0.68 % and a range from 0.13 % ( Arandisa ) to 1.37 % ( Pandercetes L. Koch, 1875 ) (Supplementary Tables 1 & 2). Intrageneric and intergeneric distances are significantly different (ANOVA, F =26.78, Games-Howell posthoc test, p <0.01). Nevertheless, the distributional tail of the intergeneric distance overlaps with the intrageneric distance. A simple distance measure or divergence cut-off thus does not allow us to distinguish between recently diverged genera and long separated congeneric species. Still, genetic distance emerges as a useful addition to morphological traits in supporting taxonomic hypotheses.
May gen. n. shows an average distance of 2.73 % compared to all other Sparassidae genera. The distances range from 1.74 % ( May vs. Arandisa ) to 4.60% ( May vs. Heteropoda ). The divergence between May bruno sp. n. and other Sparassidae genera
SUPPLEMENTARY TABLE 1 Average intergeneric distances (in percent) between Sparassidae genera used in this analysis.
Het Psp Pal Par Leu May Ceb Psm Pan Rem Ind Pan Ara Paa Car Cer Dam Oli Sin
Pseudopoda Psp Palystes Pal Parapalystes Par Leucochestris Leu May May Cebrennus Ceb Pseudomicrommata Psm Pandercetes Pan Remmius Rem gen. indet. Ind Panaretella Pan Arandisa Ara Palystella Paa Carparachne Car Cerbalus Cer Damastes Dam Olios Oli Sinopoda Sin is significantly higher than the average intergeneric distance (ANOVA, Games-Howell posthoc test, p <0.01) ( Fig. 121 View Fig ). The distance analysis for May has to be treated cautiously. As we rely on a single species to derive our measures, we artificially inflate the divergence of May and other genera.
May bruno sp. n. clusters with other African genera of the family in our phylogeny. Members of the “African clade” (sensu Moradmand et al. 2014) are among the least
Suppl. Fig. 1 View Figs 1–10 . Midpoint rooted phylogenetic tree for 850 bp of the 28SrDNA of 19 Sparassidae genera. Subfamilies and similar entities are indicated by labelled brackets to the right. Bootstrap values below 50 not shown.
distant from May gen. n. However, due to low support values the exact placement of the new genus within African Sparassidae cannot currently be resolved.A comparably large branch length separating it from other lineages indicates a long and unique evolutionary history of the genus (Supplementary Fig. 1 View Figs 1–10 ). Overall, the molecular data supports our morphological analysis very well, suggesting that May gen. n. is a distinct genus. However, with only 19 out of 84 described genera included ( World Spider Catalog 2015) our analysis is far from being exhaustive. The sequence data are deposited in the Dryad Digital Repository ( Jäger & Krehenwinkel 2015).

| SMF |
Forschungsinstitut und Natur-Museum Senckenberg |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |