
Bornella stellifer
|
publication ID |
https://doi.org/ 10.5281/zenodo.185130 |
|
DOI |
https://doi.org/10.5281/zenodo.5689391 |
|
persistent identifier |
https://treatment.plazi.org/id/03F98793-FF96-723C-2BCF-FAA2A337FE7B |
|
treatment provided by |
Plazi |
|
scientific name |
Bornella stellifer |
| status |
|
Bornella stellifer View in CoL (Adams and Reeve in Adams, 1848)
( Figures 1 View FIGURE 1 A–B, 2, 3, 4A)
Dendronotus stellifer Adams and Reeve in Adams, 1848: 494–495.
Dendronotus tenellus Adams and Reeve in Adams, 1848: 494–495.
Bornella adamsii J. E. Gray View in CoL in M. E. Gray, 1850: 107, Pl.196, fig.6; Adams and Reeve, 1850: 66–67, Pl.19, fig.3; Bertsch, 1980: 33 (for synonymy).
Bornella digitata Adams and Reeve, 1850: 66 View in CoL –67, Pl.19, fig.1; Alder and Hancock, 1864: 140–141, Pl. 33, figs 8–9; Bergh, 1874: 301–308, Pl. 37 figs 14–19, Pl 38 figs 13–22; Vayssière, 1912: 96–109, Pl. 8 figs 117–125, Pl. 9 figs 126–139; Risbec, 1953 (as`Adams 1861'): 112–113, Figs 67b, 68h, 69b.
Bornella hancockana Kelaart, 1859: 269 View in CoL –270. syn. nov.
Bornella arborescens Pease, 1871: 302 View in CoL , Pl.20, figs 3a–c; Bergh, 1890: 886 –889, Pl 88 figs 26–30, Pl 89 figs 1-2. syn. nov.
Bornella caledonica Crosse, 1875a: 318 View in CoL –319, Pl.12, fig.10; Crosse, 1875b: 325; Risbec, 1928: 216 –219, Fig.66, Pl.C, fig.3, Pl.9, fig.7; Risbec, 1930: 264. syn. nov.
Bornella marmorata Collingwood, 1879: 737 View in CoL –738 (nomen nudum)
Bornella marmorata Collingwood, 1881: 138 View in CoL –139, Pl. X, figs 34–38. syn. nov.
Bornella View in CoL ? digitata View in CoL Ad. (var. Caledonica Crosse View in CoL ): Pruvot-Fol, 1930: 231.
Bornella stellifer: Rudman, 1984: 103 View in CoL (for synonymy).
Material examined: Papua New Guinea: CASIZ 0 65739, Papua New Guinea, north coast, Madang, near lighthouse, 8 m depth, one adult specimen 28 mm preserved, dissected, 22 January 1988, coll: R.C. Willan. CASIZ 0 86313, Papua New Guinea, north coast, near Madang, south side of Rasch Passage, one adult specimen 15 mm preserved, dissected, 17 June 1992, coll: T.M. Gosliner. CASIZ 0 75076, Papua New Guinea, north coast, near Madang, south side Magic Passage (between Kranket Island and Paeowai Island), 34 m depth, one specimen 16 mm preserved, dissected, 14 November 1990, coll: T.M. Gosliner and G. Williams. CASIZ 0 75894, Papua New Guinea, near Madang, east side “Pig Island ” [Tab Island], Barracuda Point, 6 m depth, one specimen 8 mm preserved, dissected, 19 November 1990, coll: T.M. Gosliner. CASIZ 0 65421, Papua New Guinea, northern coast, South extremity of Bagabag Island, entrance to New Years Bay, 8 m depth, coral rubble, one adult specimen 17 mm preserved, 5 February 1988, coll: T.M. Gosliner. CASIZ 0 68653, Papua New Guinea, north coast, north of Madang, near Hussein Village, hole in the wall, 11 m depth, one adult specimen 16 mm preserved, 16 August 1989, coll: T.M. Gosliner. CASIZ 0 65744, Papua New Guinea, north coast, approx. 20 km north of Madang, Rempi Lagoon, 14 m depth, one specimen 15 mm preserved, 16 August 1989, coll: T.M. Gosliner. CASIZ 0 75894, Papua New Guinea, near Madang, east side “Pig Island ” [Tab Island], Barracuda Point, 30.5 m depth, three specimens 20 mm preserved, August 1989, coll: T.M. Gosliner and M. Jebb. CASIZ 0 65716, Papua New Guinea, north coast, Madang, near lighthouse, 8 m depth, one specimen 5 mm preserved, 1 February 1988, coll: T.M. Gosliner. CASIZ 0 65720, Papua New Guinea, north coast, 10 km south of Madang, Planet Rock, 30.5 m depth, one specimen 7 mm preserved, 16 February 1988, coll: T.M. Gosliner. CASIZ 0 70455, Papua New Guinea, near Madang, West side “Pig Island ” [Tab Island], Barracuda Point, 9 m depth, one specimen 10 mm preserved, 18 February 1988, coll: T.M. Gosliner. Malaysia: CASIZ 0 78438, Malaysia, Malay Peninsula, South China Sea, 16 km South of Kuala Terengganu, off Marang, Kopas Island, one specimen 35 mm preserved, 27 July 1991, coll: T.M. Gosliner. Pacific Ocean: CASIZ 0 72680, Pacific Ocean, Fiji Island, Viti Levu Island, Suva Reef, 2 specimens 12 and 28 mm preserved, dissected, November 1978, coll: M.P. Morse. CASIZ 0 72761, Pacific Ocean, Fiji Island, 2 specimens 20 mm preserved, November 1978, coll: M.P. Morse. CASIZ 0 0 6065, Pacific Ocean, Samoa Islands, one specimen 20 mm preserved, identified by MacFarland, confirmed by T.M. Gosliner, December 1991. CASIZ 0 71931, Pacific Ocean, Marshall Islands, Enewetak Atoll, Lagoon side of Enewetak Island, under coral, 5 m depth, one specimen 24 mm alive, 18 May 1983, coll: S. Johnson. C.156631, New Caledonia, Rocher a la Voile, Anse Vata, Noumea, 22° 18 S, 166° 26 E, 12 October 1988. Hawaii: CASIZ 0 71934, Hawaii, Oahu, Pupukea, in cave, one specimen 15 mm preserved, 13 August 1984, coll: S. Johnson. CASIZ 167989, Hawaii, Lanai, one specimen 20 mm preserved, February 2003, coll: T. Powers. Australia: CASIZ 168868, Australia, Queensland, Mooloolaba, southern Queensland, Inner Gneerings, The Caves, 13 m depth, one specimen 17 mm preserved, dissected, 8 July 2004, coll: S. Fahey and T.M. Gosliner. AM C.108191, Australia, Queensland, Great Barrier Reef, Capricorn Group, Heron Island, 23° 26 S, 151° 57 E, 6 specimens, October 1997, coll: A. Healy. AM C.118074, Australia, Queensland, Great Barrier Reef, Capricorn Group, Heron Island, 23° 26 S, 151° 57 E, two specimens, August 1979, coll: J. Hunter and H. Woodward. AM C.124324, Australia, Queensland, Great Barrier Reef, Capricorn Group, Heron Island, 23° 26 S, 151° 57 E, one specimen, 10 October 1980, coll: A. Healy. AM C.131378, Australia, Queensland, Great Barrier Reef, Capricorn Group, Heron Island, Blue Pools, 23° 26.216 S, 151° 54.95 E, 2 September 1981, coll: G. Avern. AM C.132316, Lord Howe Island, Neds Beach, 31° 31 S, 159° 4 E, 17 October 1981, coll: J. Hunter and H. Woodward. AM C.138555, Australia, Queensland, Great Barrier Reef, Capricorn Group, Heron Island, 23° 26 S, 151° 57 E, one specimen, 9 June 1983, coll: W.B. Rudman and G. Avern. AM C.153786, Australia, Queensland, Great Barrier Reef, Endeavour Reef, e. end, 15° 46 S, 145° 36 E, one specimen, 8 August 1987, coll: I. Loch. AM C.315471, Australia, Queensland, Great Barrier Reef, Capricorn Group, Tryon Island, 23° 15 S, 151° 47 E, one specimen, 9 August 1990, coll: I. Loch. Thailand: AM C.162372, Thailand, Phi Phi Island, 7° 45 N, 98° 47 E, one specimen, 29 November 1989, coll: D.J. Brunckhorst. India: AM C.458264, India, Pirotan Island, Gulf of Kutch, one specimen, 27 May 1971, coll: W.B. Rudman. South Africa & Madagascar: CASIZ 0 76001, South Africa, Natal Province, two specimens 20 & 30 mm preserved, 27 December 1958, identified by T.M. Gosliner. CASIZ 0 70402, Madagascar, Nosy Be, reef just north of small Island, off Les Cocotiers Hotel, two specimens 7 and 12 mm preserved, dissected, 17 April 1989, coll: T.M. Gosliner.
Geographic Distribution: Indo-West Pacific: originally described from the South China Sea ( Adams & Reeve 1848, present study) this species is also found in Australia ( Angas 1864; Smith 1884; Allan 1950; Burn 1965; Kenny 1970; Coleman 1989, 2001; Debelius 1998; Marshall & Willan 1999; present study), Korea ( Koh 2006), Indian Ocean ( Kelaart 1859; Collingwood 1881; Eliot 1905; Vayssière 1912; Yonow 1990; present study), South Africa ( Gosliner 1987a; present study), East Africa ( Eliot 1904), Philippines ( Bergh 1874), Arafura Sea ( Bergh 1884), China Sea ( Collingwood 1868), Japan ( Eliot 1913; Baba 1937; Suzuki 2000), Hong Kong ( Orr 1981), Tahiti ( Pease 1871), New Caledonia ( Crosse 1875a; Risbec 1928, 1932; present study), Taiwan ( Garthwaite 2002), Thailand, Marshall Islands and Papua New Guinea (present study).
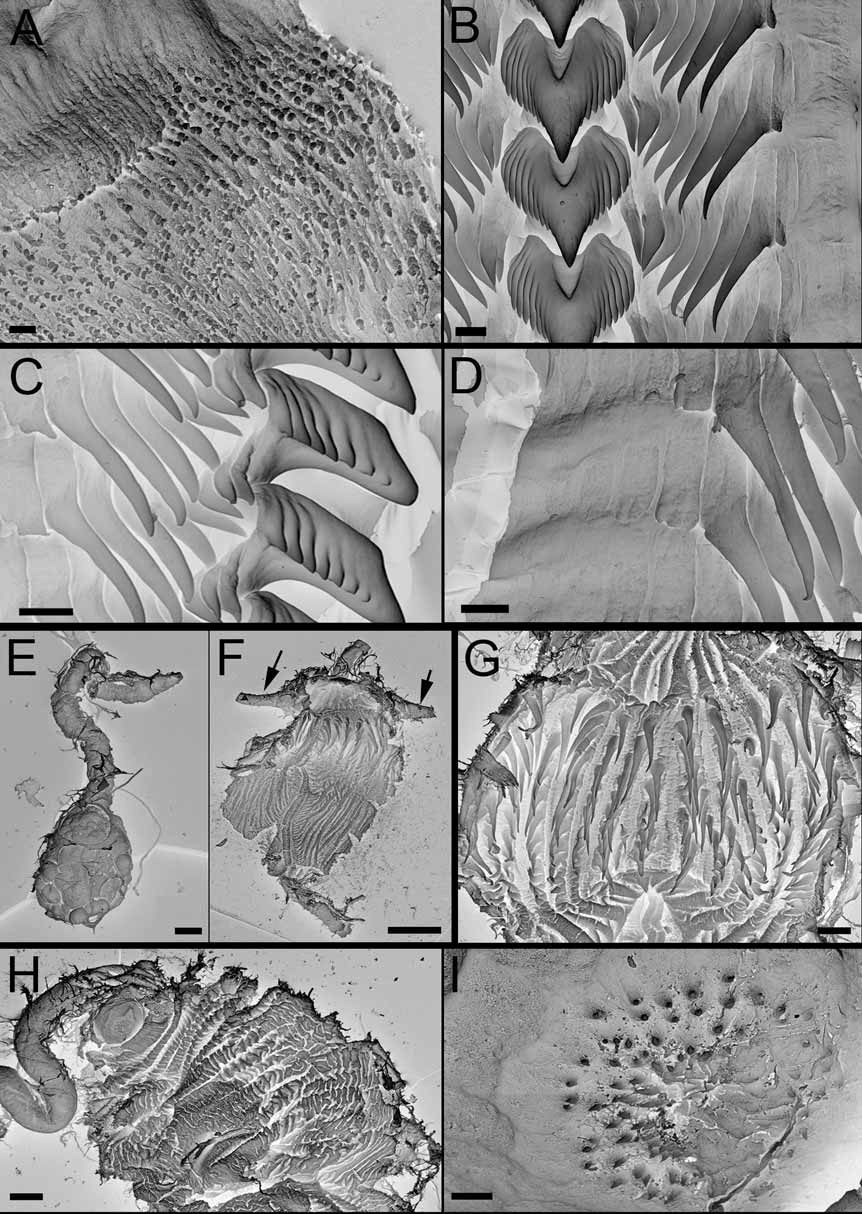
External morphology: The general body shape is elongate and limaciform with the posterior end of the foot being long and tapering. The living adults are reported up to 48 mm in length. The animal is translucent cream-white with a network of orange markings on the back and sides of the body and scattered sub epidermal opaque white granules ( Figs. 1 View FIGURE 1 A,B). This color pattern extends to the rhinophore sheaths. The lobe-like oral tentacles, rhinophoral papillae and dorsolateral processes are translucent white and have a subapical orange ring. The transparency of the body walls varies in different specimens. The head is rounded and bears a modified lobe-like oral tentacle on each side of the mouth. In most specimens each lobe-like oral tentacle is a palmate structure with up to 13 finger-like papillae, of unequal length, arranged in two distinct marginal rows. In several immature specimens the number of digitiform papillae was lower, between seven and nine. The rhinophores are perfoliate with about 13 to 20 translucent white lamellae. The rhinophore sheath has a tall stalk which branches distally into three elongate anterior and antero-lateral papillae and one posterior bulbous, slightly laterally compressed papilla, about twice the height of the others. In one specimen, we found two posterior papillae ( Fig. 1 View FIGURE 1 B) but they were both unbranched. Posterior to the rhinophores, there are five to six pairs of dorsolateral processes, followed by two single, small processes in the dorsal midline. The number of papillae on the dorsolateral processes varies among specimens. Usually, there are three bulbous, pointed papillae on the first and second pair, while in the others there are only two, except for the last pair, which is always simple. The anterior four pairs of dorsolateral processes bear a translucent tripinnate gill on the inner surface, but the fifth pair (and sixth when present) and the single posterior-most process lack gills. The anus is small, located on the right side of the dorsum between the first and second pair of dorsolateral processes, closer to the second. The reproductive opening is located on the right side, midway between the rhinophore sheath and the first dorsolateral process.
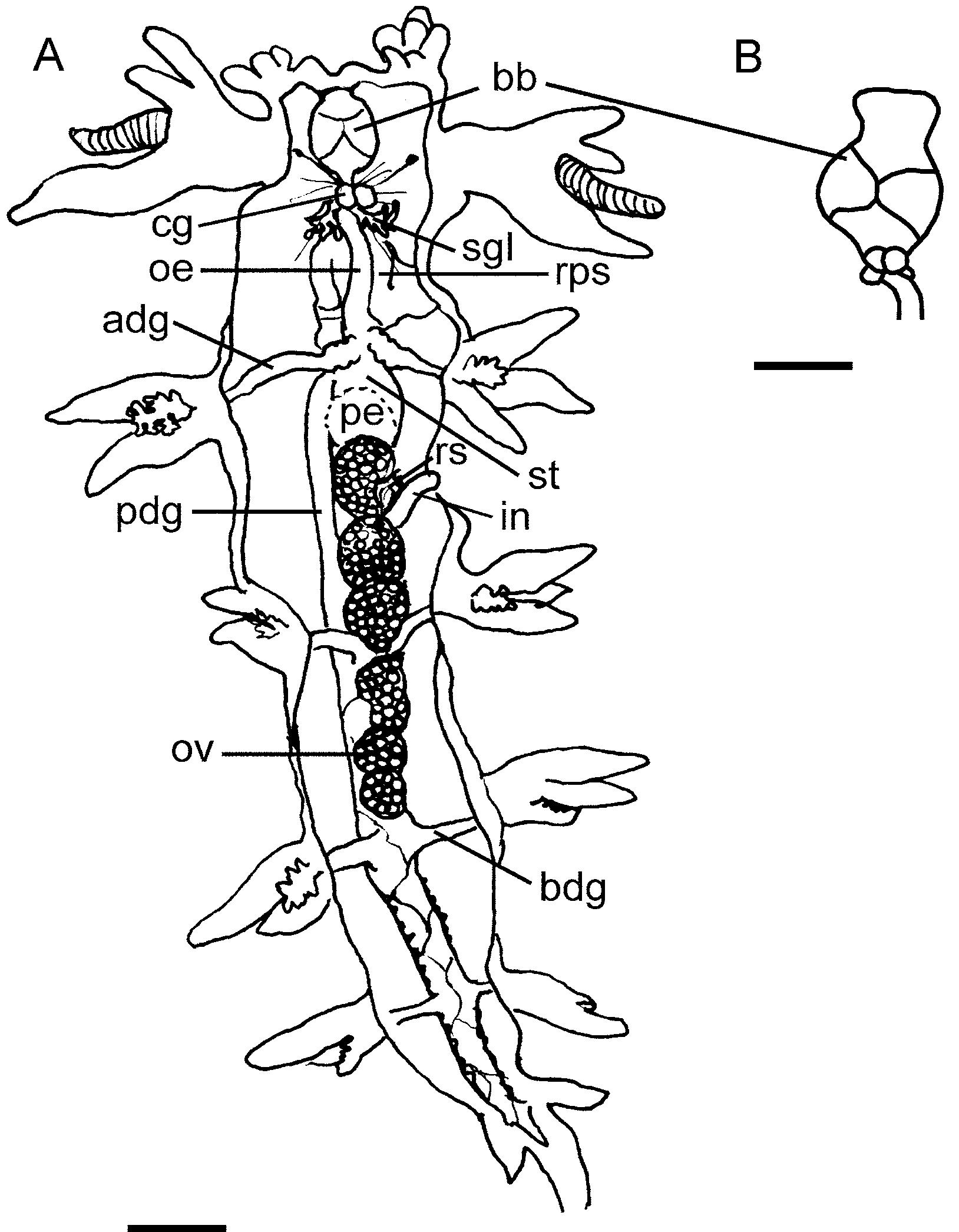
Alimentary Canal: ( Figure 2 View FIGURE 2. A ) The buccal bulb is relatively small in comparison with the other species reported here, and the labial cuticle is thin, consisting of small overlapping scales, arranged in fairly regular rows. The rodlets are elongate but not as much numerous as in other species of the genus ( Fig. 3A View FIGURE 3. A – F , present study). The jaws are roughly oval, without a distinct masticatory process. The radular formulae of some of the specimens dissected are: 35 x 14.1.14 (CASIZ 0 72680, 28 mm preserved), 35 x 9.1.9 (CASIZ 168868, 17 mm preserved), 33 x 9.1.9 (CASIZ 0 65739, 28 mm preserved), 28 x 8 –9.1.8–9 (CASIZ 0 70402, 12 mm preserved, CASIZ 0 75076, 16 mm preserved) and 27 x 7.1.7 (CASIZ 0 86313, 15 mm preserved). The rachidian teeth are stout, with about seven to 10 denticles (usually nine) on either side of a pointed cusp, increasing in size away from the cusp ( Figs. 3B,C View FIGURE 3. A – F ). Most of the specimens examined have from seven to nine laterals ( Figs. 3B,C View FIGURE 3. A – F ), but in some specimens the number rises to 14. The inner laterals have a long basal portion and then are bladelike and pointed, increasing in size away from the center. But after the first five to nine blade-like laterals, there is usually a denticle with a short cusp followed by from one to three (even four) plate-like laterals ( Figs. 3B–D View FIGURE 3. A – F ). A long unpaired oral gland is found ventrally ( Fig. 3E View FIGURE 3. A – F ). It opens in the ventral midline of the mouth and extends back, below the buccal bulb, to the region of the reproductive system. The paired, branched, salivary glands are yellowish in preservative, and attached along the sides of the oesophagus. They open into the posterior buccal bulb on each side of the oesophageal opening. The oesophagus is elongate, entering straight into the rounded stomach. On the upper surface of the stomach are the openings of the two anterior digestive glands ( Fig. 3F View FIGURE 3. A – F ). Each anterior digestive gland has a single branch to the first dorsolateral process on its side. On the lower left surface of the stomach is the third digestive gland opening, which leads to the posterior portion of the digestive gland. All dorsolateral processes, other than the anteriormost pair already mentioned, receive branches from the posterior portion of the digestive gland. The posterior section of the stomach has become a gizzard-like chamber armed with spines, possibly to prevent undigested particles escaping into the intestine. In B. stellifer this posterior chamber has thin walls and is much more elongate than in the remaining species of the genus (present study) ( Fig. 3F View FIGURE 3. A – F ). It is armed with about 12 to 15 longitudinal rows of chitinous brownish spines. These spines are straight and acutely pointed ( Fig. 3 View FIGURE 3. A – F G). The rest of the walls are raised into well-defined longitudinal folds. From the posterior stomach, the intestine folds down and then bend dorsally to the anus.
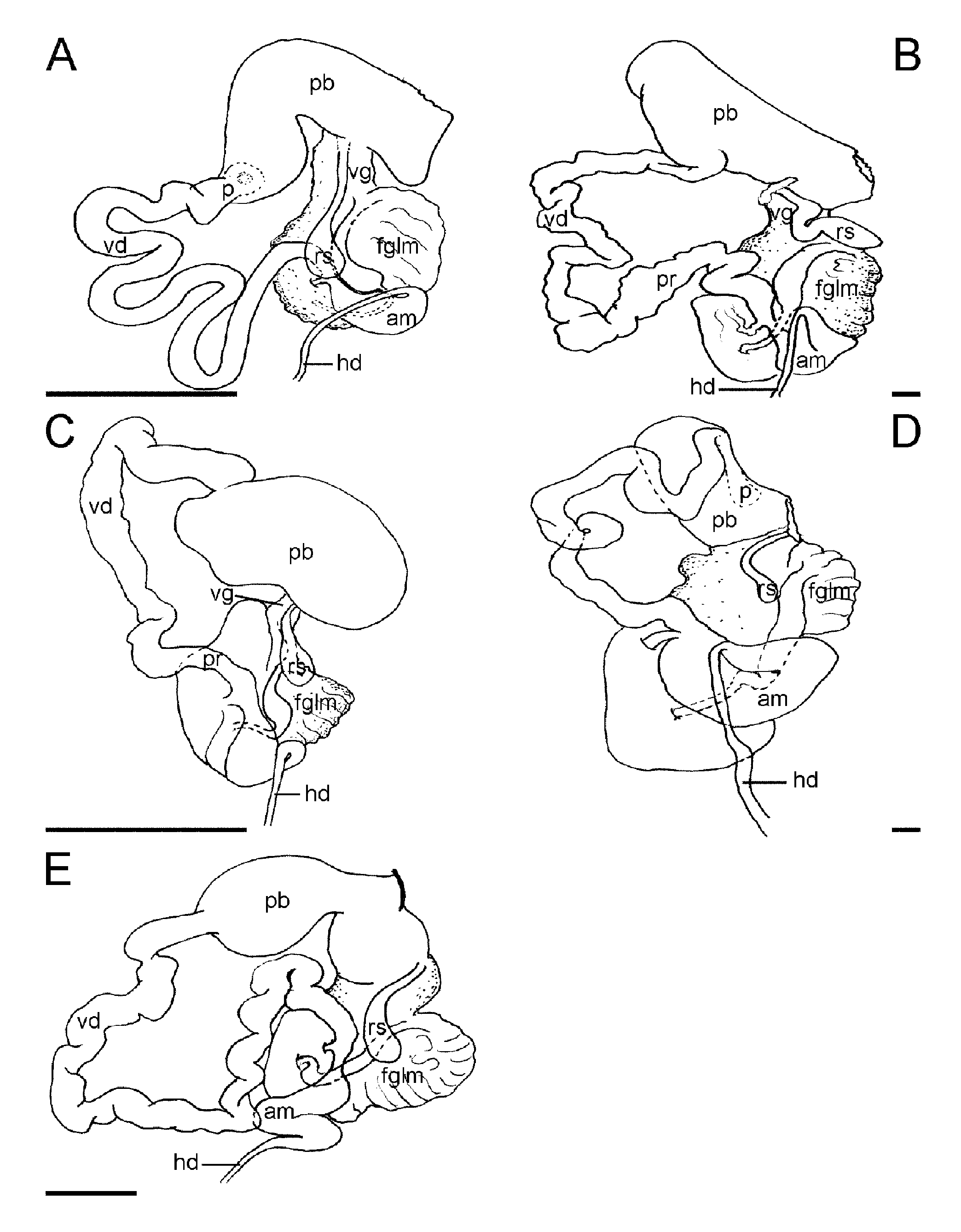
Reproductive system: ( Fig. 4A View FIGURE 4. A – E ) The ovotestis consists of about five to six closely packed, rounded to pyriform follicles lying in the middle third of the body cavity above the posterior digestive gland ( Fig. 2 View FIGURE 2. A ). From each follicle, a thin-walled duct passes forwards to the hermaphroditic duct, which passes around the left side and below the stomach to join the ampulla on the dorsal side of the female gland mass. At its anterior end the ampulla narrows, branches into two ducts, the vas deferens and the oviduct. The vas deferens is long and somewhat folded. It lacks a discrete prostate gland mass, but its whole length is lined with a uniform layer of prostate gland tissue except for a short muscular region near the penial bulb. The vas deferens opens directly into the large curved penial bulb, the opening being armed with more than two circular rows of chitinous hooked spines ( Figs. 3 View FIGURE 3. A – F H, I). There is no distinct penial papilla, but it is possible that the non-prostatic section of the vas deferens everts to form a temporary papilla during mating. From its branching with the vas deferens, the oviduct opens immediately into the female gland mass. Without histological studies it is not possible to determine the passage of eggs and allosperm within the female gland mass, but from gross anatomy it appears to be similar to that described from Tritonia hombergi (see Thompson 1961). A medium-sized stalked distal allosperm receptacle opens into the common genital opening alongside the opening of the female gland mass.
Remarks: Bornella stellifer has a complicated nomenclatural history, which has been reviewed by Bertsch (1980) and Rudman (1984), the latter author showing that the correct name for the type species is B. stellifer . Externally, B. stellifer is easily distinguishable from other species of the genus by the presence of an orange subapical ring located on the lobe-like oral tentacles, papillae of the rhinophore sheaths and dorsolateral processes. Additionally, this species has simple bulbous papillae on the posterior side of each rhinophore sheath and on the dorsolateral processes. Internally, this species differs from others in the genus in having a much more elongated posterior chamber to the stomach with fewer rows of sharpened spines. Within the reproductive system there is also a good character that differentiates it from the other species. Bornella stellifer is the only species in which the penial spines are arranged in more than two rings around the penial opening. In B. johnsonorum they are arranged in two rings, while in B. pele , B. dotoides and B. hermanni there is a single ring. In B. calcarata , B. valdae and B. anguilla the penial spines are arranged on a raised fleshy ridge and in Bornella sarape penial spines are absent. Bornella stellifer is most similar to B. johnsonorum sp. nov. from the Marshall Islands as we discuss under that species. Bornella stellifer is common throughout the Indo- West Pacific. Its name refers to the star-like arrangement of the papillae on the oral lobes. The name Bornella digitata , a synonym of this species, also referred to these lobes, describing them as 'finger-like'. Apart from the earlier synonyms discussed by Bertsch (1980) and Rudman (1984), four other names, which were all described from external features only, are considered here to be synonyms of B. stellifer . The description of both Bornella hancockana Kelaart, 1859 from Sri Lanka and Bornella arborescens Pease, 1871 , from Tahiti, include mention of the reticulate colour pattern and in particular red subterminal rings on the dorsolateral processes. They also mention four or more pairs of dorsolateral processes, which precludes B. hermanni . Bornella caledonica Crosse, 1875 a from New Caledonia is not well categorized. Its colour is described as yelloworange, irregularly spotted with ‘rouge vermillon’ on the dorsum and branchial appendages. Although no actual mention of orange subterminal rings is mentioned, the color painting accompanying the description shows five pairs of dorsolateral processes which suggest this species is B. stellifer . The fourth name, B. marmorata Collingwood, 1881 is based on three animals collected in Aden. Unfortunately no illustration of the whole animal was published but the rhinophore sheath and posterior appendage is as found in B. stellifer . The color description mentions streaks of ‘vermilion’ and the dorsal processes are described as ‘tipped with vermilion’. There is no mention of a subterminal ring but in some specimens the orange ring forms a broad band reaching almost to the tip of the process. The mention of seven pairs of ‘marginal processes’ would also suggest this species is B. stellifer . Another character, which is typical of B. stellifer , and present in the illustrations of all these four species, is that the posterior appendage on the rhinophore sheath is large and unbranched. One further name, Bornella semperi Crosse 1875 b is most probably B. stellifer but is unidentifiable and considered here to be a nomen nudum. It is based on a painting from the Semper expeditions, published but left unnamed by Bergh (1870). As Bergh (1884: 36) vociferously argues, “The creation of this species is a good example of the “species manufacturing” of many writers. …..Writers of this kind are a burden to science, and Malacology has had enough of them”.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Bornella stellifer
| Pola, Marta, Rudman, William B. & Gosliner, Terrence M. 2009 |
Bornella stellifer:
| Rudman 1984: 103 |
Bornella
| Pruvot-Fol 1930: 231 |
Bornella marmorata
| Collingwood 1881: 138 |
Bornella marmorata
| Collingwood 1879: 737 |
Bornella caledonica
| Risbec 1930: 264 |
| Risbec 1928: 216 |
| Crosse 1875: 318 |
| Crosse 1875: 325 |
Bornella arborescens
| Bergh 1890: 886 |
| Pease 1871: 302 |
Bornella hancockana
| Kelaart 1859: 269 |