
Mini mum, Scherz & Hutter & Rakotoarison & Riemann & Rödel & Ndriantsoa & Glos & Roberts & Crottini & Vences & Glaw, 2019
|
publication ID |
https://doi.org/10.1371/journal.pone.0213314 |
|
persistent identifier |
https://treatment.plazi.org/id/03F887F6-FF9E-0A59-FD8B-0501DE896AC8 |
|
treatment provided by |
Plazi |
|
scientific name |
Mini mum |
| status |
sp. nov. |
Mini mum View in CoL sp. nov.
urn:lsid:zoobank.org:act:237AA825-4612-4591-9CC8-764DAD646B48
( Figs 1–6 View Fig 1 View Fig 2 View Fig 3 View Fig 4 View Fig 5 View Fig 6 , Tables 1 and 2 View Table 2 )
Remark. This species was previously listed as Stumpffia sp. 10 [ 17]; Stumpffia sp. 15/Ca15 [ 15, 40]; Stumpffia sp. aff. tetradactyla “Southeast” [ 41]; and Stumpffia sp. 10 KCW-2008 ( EU341082 View Materials ) [ 20].
Holotype ( Figs 2 View Fig 2 , 4 View Fig 4 and 6 View Fig 6 ). ZSM 861 View Materials /2014 ( ZCMV 14788 ), an adult presumed male specimen collected in Manombo Special Reserve (23.0294˚S, 47.7312˚E, 7 m a.s.l.), Atsimo-Atsinanana Region, former Fianarantsoa province, southeastern Madagascar on 30 November 2014 by A. Rakotoarison and E. Rajeriarison.
Paratypes ( Fig 4 View Fig 4 ). ZSM 862 View Materials /2014 ( ZCMV 14789 ), an adult presumed male specimen with the same collection data as the holotype ; ZMA 20172 ( ZCMV 557 ), an adult presumed male specimen, and ZMA 20191 ( ZCMV 558 , GenBank accession number EU341082 View Materials for 12S rRNA gene, tRNA-Val, and 16S rRNA gene), an unsexed specimen collected in Manombo Special Reserve (23.0284˚S, 47.7316˚E, 44 m a.s.l.) on 2 February 2004 by D.R. Vieites and C. Woodhead GoogleMaps ; ZMB 81993 ( NSH 2584 ) and ZMB 83194 ( NSH 2583 ), adult male and female specimens (respectively) collected in Manombo Special Reserve (23.0249˚S, 47.7311˚E, ca. 20 m a.s.l.) GoogleMaps
on 28 March 2012 by J.C. Riemann, S.H. Ndriantsoa, A. Rakotoarison, J. Glos and M.-O. Rödel.
Diagnosis. An extremely miniaturised frog assigned to Mini gen. nov. on the basis of its small size, curved clavicles, laterally displaced and reduced nasals, and fusion or loss of carpal 2. This assignment is supported by its genetic affinities ( Fig 1 View Fig 1 ; [ 15, 17, 20]). It is separated by uncorrected p-distances of 10.0–11.2% in the analysed 3’ fragment of the 16S rRNA gene from other members of the genus Mini gen. nov., and 8.3–12.4% from all members of the genus Plethodontohyla .
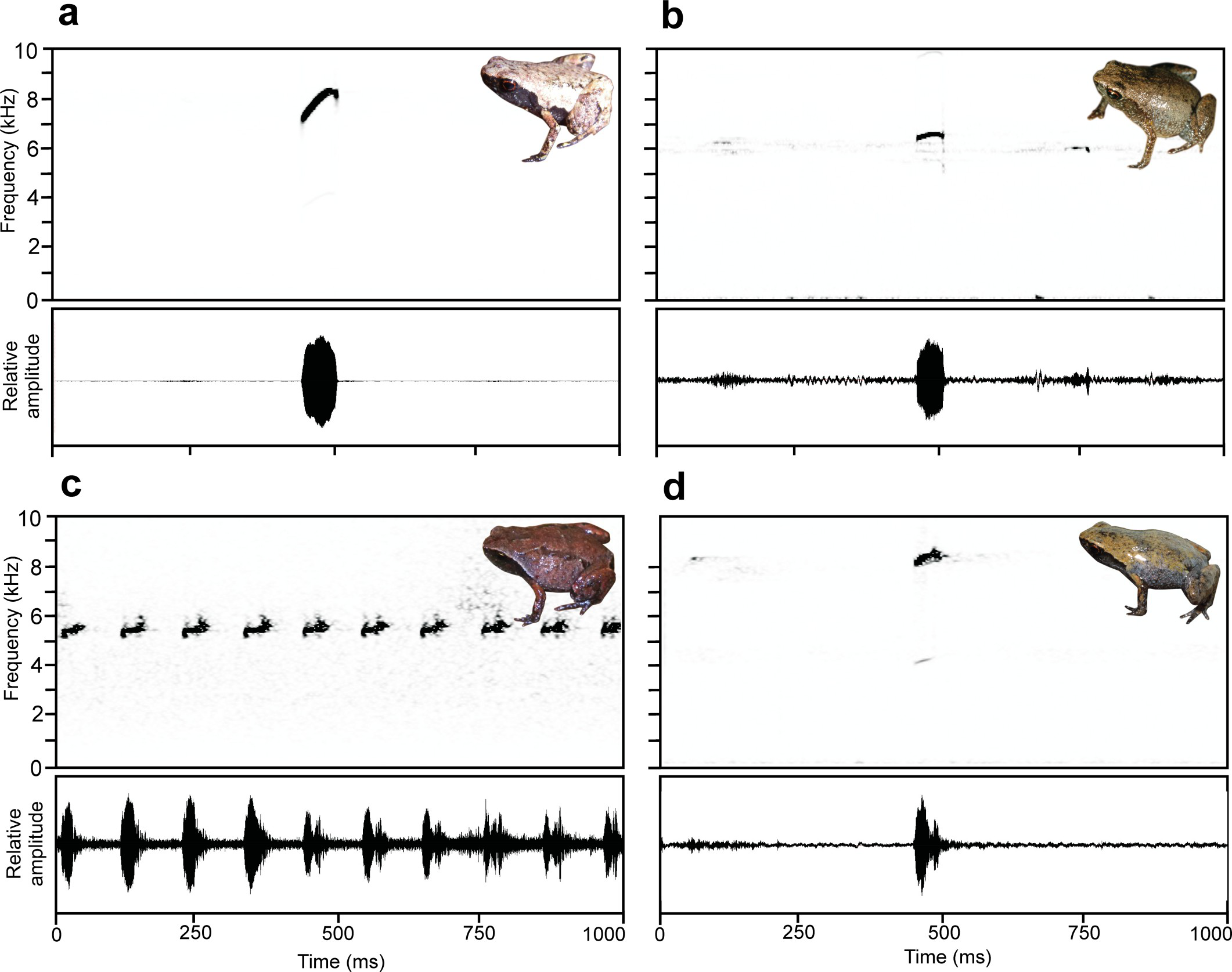
Mini mum sp. nov. is characterised by the unique combination of the following characters (n = 4 male specimens, 1 female specimen): (1) male SVL 8.2–9.7 mm, female SVL 11.3 mm; (2) ED/HL 0.38–0.56; (3) HW/SVL 0.28–0.37; (4) FARL/SVL 0.30–0.38; (5) TIBL/SVL 0.39– 0.50; (6) HIL/SVL 1.47–1.72; (7) fingers 1, 2, and 4 strongly reduced; (8) toe 1 absent, toes 2 and 5 quite reduced; (9) maxillary and premaxillary teeth absent; (10) vomerine teeth absent; (11) strong lateral colour border present; (12) black inguinal spots absent; (13) postchoanal vomer present, spatulate, medially fused to parasphenoid; (14) nasal cultriform and laterally displaced; (15) quadratojugal-maxilla contact weak; (16) zygomatic ramus of squamosal short, narrow, and horizontal; (17) clavicles present, curving with simple lateral articulations, medially not bulbous; (18) prepollex small or absent; (19) carpal 2 absent or fused to post-axial carpal 3–5 element; (20) finger phalangeal formula 1-2-3-2; (21) toe phalangeal formula 1-2-3-4- 3; (22) single-note, unpulsed calls, not emitted in series; (23) frequency modulated calls; (24) call dominant frequency 8089 ± 140 Hz (n = 35); (25) call duration 74.8 ± 7.0 ms (n = 35); (26) inter-call interval 4299.8 ± 1604.9 ms (n = 34).
Within the genus Mini gen. nov., the new species is unique in lacking teeth, and possessing a strong lateral colour border. See other species described below for respective diagnoses.
This species is particularly similar to some extremely miniaturised Stumpffia species, but it can be distinguished from all Stumpffia based on the condition of the carpals, and all Stumpffia except S. tridactyla, S. contumelia, and S. obscoena by the extent of reduction of its fingers and toes. It differs from all of these in possessing curved clavicles (vs absent in S. contumelia and S. obscoena and straight or absent in S. tridactyla; unpublished data), and presence of neopalatine and divided vomer (vs absence of neopalatine and non-divided vomer in S. obscoena, S. tridactyla, and S. contumelia; unpublished data).
Calls resemble those of S. miery, S. tridactyla, S. contumelia, and S. obscoena, but are shorter in duration and lower in frequency than S. obscoena (duration 74.8 ± 7.0 ms vs 144 ± 8 ms; frequency 8089 ± 140 Hz vs 8361 ± 69 Hz); longer in duration and higher in frequency than S. contumelia (duration 74.8 ± 7.0 ms vs 42 ± 4 ms; frequency 8089 ± 140 Hz vs 7493 ± 50 Hz); shorter in duration and higher in frequency than S. tridactyla (duration 74.8 ± 7.0 ms vs
132 ± 23 ms; frequency 8089 ± 140 Hz vs 7244 ± 200 Hz); and slightly longer inter-call interval than S. miery (4299.8 ± 1604.9 ms vs 3102 ± 456 ms).
Holotype description. Specimen in a good state of preservation, a piece of the left thigh removed as a tissue sample. Body oblong; head wider than long, narrower than body width; snout rounded in dorsal view, pointed in lateral view; nostrils directed laterally, not protuberant, slightly further from tip of snout than from eye; canthus rostralis indistinct, straight; loreal region flat, vertical; tympanum indistinct, round, about 49% of eye diameter; round pupil; supratympanic fold absent; tongue long, broadening slightly posteriorly, attached anteriorly, not notched; maxillary teeth absent; vomerine teeth absent; choanae small and round. Forelimbs slender; subarticular tubercles single, indistinct except on third finger; outer metacarpal tubercle rounded; inner metacarpal tubercle indistinguishable from reduced first finger; hand without webbing; first, second, and fourth fingers strongly reduced, third finger basally broadened; relative length of fingers 1 <4 <2 <3, fourth finger slightly more reduced than second; finger tips not expanded into discs. Hindlimbs slender; TIBL 50% of SVL; lateral metatarsalia strongly connected; inner metatarsal tubercle indistinguishable from completely reduced first toe; outer metatarsal tubercle absent; no webbing between toes; first toe absent, second and fifth toes extremely reduced; relative length of toes 2 <5 <3 <4; fifth toe distinctly shorter than third. Skin on dorsum smooth, without distinct dorsolateral folds. Ventral skin smooth.
After four years in 70% ethanol, the dorsum is metallic silver centrally on the trunk, bluish silver on the head, and laterally light silver, with dark oblong markings in the inguinal region ( Fig 2 View Fig 2 ). There is a strong dorsolateral colour border to the ebony lateral colouration, extending from the side of the head to the legs. The lateral colouration fades to the more burnt umber ventral colouration, especially dark anteriorly, flecked with beige, fading posteriorly through larger fleck sizes to beige at the posterior abdomen. Dorsally, the legs are mottled cream and grey brown with a dark cloacal region. Ventrally the legs are brown flecked with beige. The arms are silvery dorsally and ebony laterally and ventrally. Colour in life ( Fig 4 View Fig 4 A–4C) as in preservative but browner in every aspect and less obviously iridescent, with a red iris.
Variation. For variation in measurements among specimens, see Table 1. Non-ovigerous specimens with darkened throats are presumed to be males, in keeping with the one call voucher, ZMB 81993. In general, all examined specimens agree with the holotype in morphology, but female ZMB 83194 is more rotund in body shape in preservative, but had a longer, depressed body profile in life ( Fig 4F and 4G View Fig 4 ). ZMB 83194 and 83193 varied in life from smooth to slightly granular skin, with a very faintly bulging vertebral line. The colouration in life varied rather strongly ( Fig 4 View Fig 4 ). Lateral and ventral colouration was more or less consistently dark brown with light flecks, but ZMB 81993 had bright bluish-white flecks laterally. The strength of the flank colour border varied from stark in ZSM 862/2014 to weak in ZMB 83194. Dorsal colouration varied from solid tan in ZSM 862/2014 to mottled beige and dark brown in ZMB 83194. Iris colouration was consistently red to reddish copper. In preservative, paratypes are less iridescent than the holotype, and ZMB 81993 and 83914 are faded to light brown.
Bioacoustics. Calls recorded from specimen ZMB 81993 ( Fig 5A View Fig 5 , Table 2 View Table 2 ) during the day on 28 March 2012 (see paratype section for locality data). Air temperature was 21.7˚C. The specimen was calling under dense leaf litter. Estimated call parameters were as follows (n = 35 in all cases except inter-call and call intervals, where n = 34): Calls consisting of a single note were emitted at regular intervals without defined call series. Calls had linear upward frequency modulation with a downward hooked tail, with an initial dominant frequency around 7000 Hz, rising gradually to ca. 8250 Hz, and with a tail-end dropping down to ca. 7500 Hz again. For detailed parameters, see Table 2 View Table 2 .
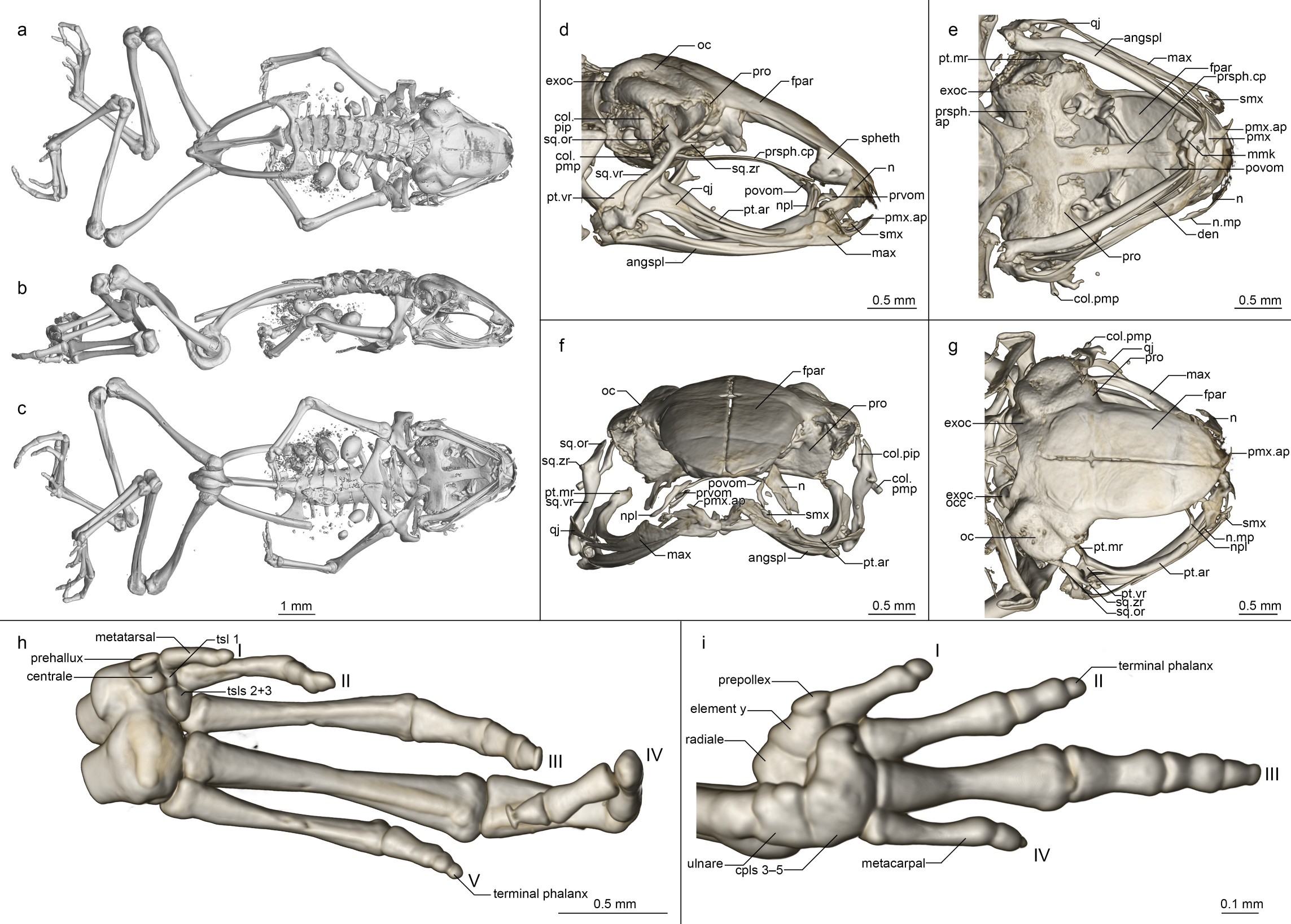
Osteology ( Fig 6 View Fig 6 ). Based on ZSM 861/2014 (figured), and ZSM 862/2014, ZMA 20172, ZMB 81993 and ZMB 83194 (not figured). Note that the skulls of ZSM 861–862/2014 are somewhat distorted in fixation, especially with respect to the maxillary arcade and mandible. The skull and pectoral girdle of ZMB 81993 are quite badly damaged, and both of its hindlimbs are fractured, as are the coracoids and left ilium of ZMB 83194.
Cranium ( Fig 6 View Fig 6 D–6G). Shape and proportions. Skull short and rounded, longer than wide, widest at the bowing of the quadratojugal roughly in line with the anterior face of the prootic. Braincase proportionally broad, with an extremely short rostrum.
Neurocranium. Ossification generally high, lower in ZSM 862/2014 than other specimens. Anterior cone of sphenethmoid ossified and in contact with the frontoparietals in ZSM 861/ 2014 and ZMB 83194 and 81993, but no sphenethmoid ossification in the other specimens. Prootic in dorsal contact with lateral flange of frontoparietal, ventral contact with parasphenoid alae, not approaching contralateral ventrally. Septomaxilla miniscule, very tightly curled, not further discussed due to low ossification and insufficient resolution. Columella (stapes) well ossified, pars media plectra (stylus) long and nearly straight, broadening distally, posteriorly and dorsally oriented toward the dorsally elongated pars interna plectra (baseplate). Nasal narrow and cultriform, laterally displaced (in line with anterior end of frontoparietal), curved downward laterally, the acuminate maxillary process not distinct and not closely approaching maxillary pars facialis, broadly separated from contralateral. Frontoparietal with rounded anterior edge, laterally rather straight-edged, with short lateral flange covering prootic, posteriorly strongly connected to exoccipital, anteroventrally contacting sphenethmoid in ZSM 861/2014, lacking any dorsal process, separated from contralateral by a narrow gap with a small rhomboid facet at the level of the prootics, possibly constituting the pineal foramen.
Parasphenoid with narrow, rather straight-edged cultriform process and slightly broader perpendicular alae, considerably shorter than frontoparietals, in contact with exoccipitals posterodorsally, prootics dorsally along the edges of the alae, anteroventrally in contact with postchoanal vomer and not in contact with neopalatine; posteromedial process not participating in foramen magnum. Vomer divided into pre- and postchoanal portions; prechoanal portion narrow, simple, sickle-shaped, without a lateral ramus; postchoanal portion spatulate and edentate, narrowly separated from its contralateral on the midline, in dorsal contact with the parasphenoid proximally and the neopalatine distally, lacking an anterior projection. Neopalatine simple, straight, almost indistinguishable from lateral postchoanal portion of vomer, laterally broadly separated from the maxilla, not exceeding the lateral-most point of the postchoanal vomer.
Maxillary arcade gracile, maxilla and premaxilla edentate, anterior extension of maxilla not exceeding lateral extent of premaxilla. Premaxilla with a narrow acuminate dorsal alary process rising laterally, pars palatina shallowly divided into a narrow palatine process and broad, squared lateral process. Maxilla with a low triangular pars facialis and a narrow pars palatina, its posterior tip acuminate and barely contacting the quadratojugal, the lingual surface of the pars palatina followed by but not contacting the anterior ramus of the pterygoid, presumably separated by the pterygoid cartilage. Pterygoid with an exceptionally short medial ramus, long anterior ramus, and broad posterior ramus, posteriorly weakly calcified to the quadratojugal complex. Quadratojugal bowed laterally, broadly connected to the ventral ramus of the squamosal, bearing a small posteroventral knob, weakly anteriorly connected to the maxilla; the articulation of the mandible is apparently somewhat fortified by the mineralisation of the posterior ramus of the pterygoid+squamosal+quadratojugal posteroventral knob. Squamosal with a slender, rather straight ventral ramus, broadened, nearly vertical otic ramus, and short, thin, horizontal zygomatic ramus.
Mandible slim and edentate, largely unremarkable, with a weakly raised coronoid process on the angulosplenial. Mentomeckelians separated from the dentary, with small hooked ventrolateral projections.
Posteromedial processes of hyoid proximally rounded with a broad medial crista.
Postcranial Skeleton ( Fig 6 View Fig 6 A–6C, 6H and 6I). Eight procoelous presacrals, all much broader than long, lacking neural spines, with round posterior articular processes, presacral I with a complete neural arch, presacrals II–IV with thicker and longer transverse processes than V–VIII. Sacrum with expanded diapophyses, the leading and trailing edges roughly equally angled, the articulation type IIB sensu Emerson [ 42]. Urostyle bicondylar, long, broadening posteriorly, with a somewhat flared head and a low dorsal ridge.
Pectoral girdle without ossified prezonal or postzonal elements, with ossified clavicles. Clavicle thin and weakly curved, with a simple lateral junction, slightly shorter than the coracoid. Coracoid fairly narrow, weakly flared laterally, strongly flared medially with a straight medial articular surface with the contralateral. Scapula slender, with a thin pars acromialis, the cleitheral border straight. Cleithrum ossified for half the width of the scapular border, thickened anteriorly. Suprascapula unossified.
Humerus with a well-developed crista ventralis and no medial or lateral cristae. Radioulna slender with a distinct sulcus intermedius. Carpals apparently reduced, composed of radiale, ulnare, element Y, prepollex, and a large post-axial element formed by carpals 3–5. Carpal 2 has either been lost or fused to the latter element. Finger phalangeal formula is reduced (1-2-3-2), and the terminal phalanges of the first, second and fourth fingers are small, round elements.
Pubis ossified; iliac shafts passing ventral to and beyond sacrum, oblong in cross-section, with a weak dorsal crest, without a dorsal prominence and with a shallow oblique groove. Femur weakly sigmoid, lacking a posterior crest. Tibiofibula slightly longer than femur in length, with a sulcus intermedius. Tibiale and fibulare fused proximally and distally. T1 and T2 +3 tarsals present, T1 considerably smaller than T2+3. Centrale present, slightly smaller than T2+3. Prehallux small. Phalangeal formula reduced (1-2-3-4-3). Terminal phalanges of toes 3 and 4 with knobs, those of other toes small, round elements.
Distribution, natural history, and conservation status. This species is known only from Manombo Special Reserve, southeast Madagascar ( Fig 3 View Fig 3 ). The habitat consists of low, comparatively open forest with small trees, many lianas and a very thick layer of dead leaves ( Fig 4J View Fig 4 ). Calls were emitted by males during the day, hiding within the leaf litter or between roots, separated from other calling males by several metres. The female paratype ZMB 83194 contains four eggs (visualised by micro-CT scan, not extracted for physical examination). Manombo Special Reserve covers an area of 52.66 km 2. We estimate that this species occurs from 0–100 m a.s.l. within lowland forests in and around this reserve. Although lowland species from areas with low topographical complexity tend not to be extreme micro-endemics [ 43], extremely miniaturized frogs in Madagascar almost always are [ 14, 17], so the full extent of this species’ range is not likely to be large. Littoral forest in the area where the species occurs is extremely reduced, so other sites outside the Special Reserve are likely to be small and under high pressure. We therefore recommend this species be listed as Critically Endangered according to the IUCN Red List Criterion CR B1ab(iii) [ 44], in line with other endemics from Manombo Special Reserve (e.g. Guibemantis diphonus [ 45]).
The gut of ZSM 861/2014 contains four or five arthropods visible from micro-CT scans, tentatively identified as oribatid mites.
Etymology. We use the specific epithet ‘mum’ as an arbitrary combination of letters, in order to form a pun on ‘minimum’ from the name in apposition, in reference to the fact that this is among the smallest known frogs from Madagascar and the world. It is to be regarded as an invariable noun.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.