
Cotylegaleata iskenderunensis, De Smet, Willem H. & Bozkurt, Ahmet, 2016
|
publication ID |
https://doi.org/10.11646/zootaxa.4193.1.9 |
|
publication LSID |
lsid:zoobank.org:pub:CE0D352E-C155-4D3D-830D-1877683483E0 |
|
DOI |
https://doi.org/10.5281/zenodo.6078138 |
|
persistent identifier |
https://treatment.plazi.org/id/03F887CC-5901-8357-FF29-FE26FB4F1B2B |
|
treatment provided by |
Plazi |
|
scientific name |
Cotylegaleata iskenderunensis |
| status |
sp. nov. |
Cotylegaleata iskenderunensis n. sp.
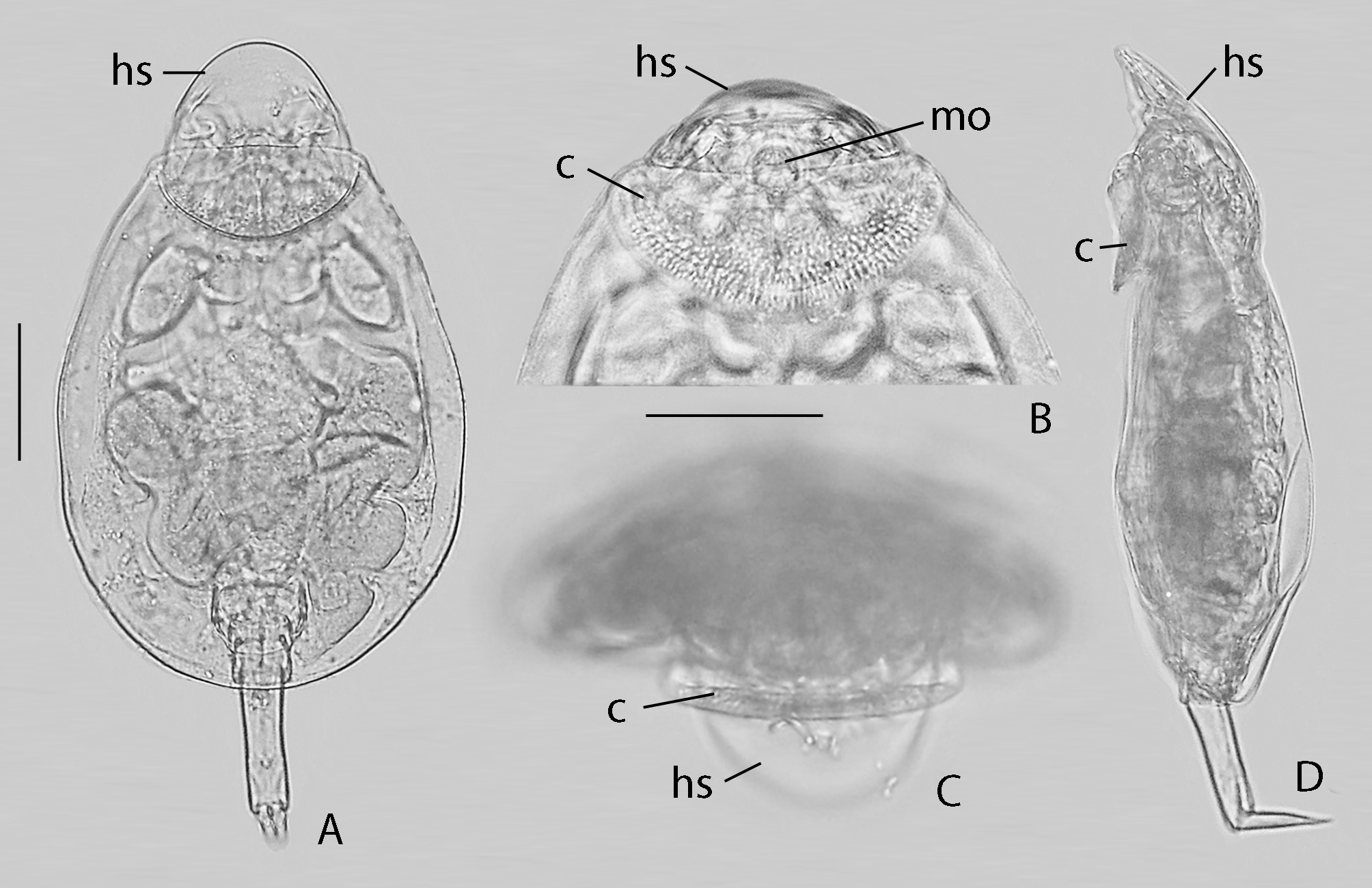
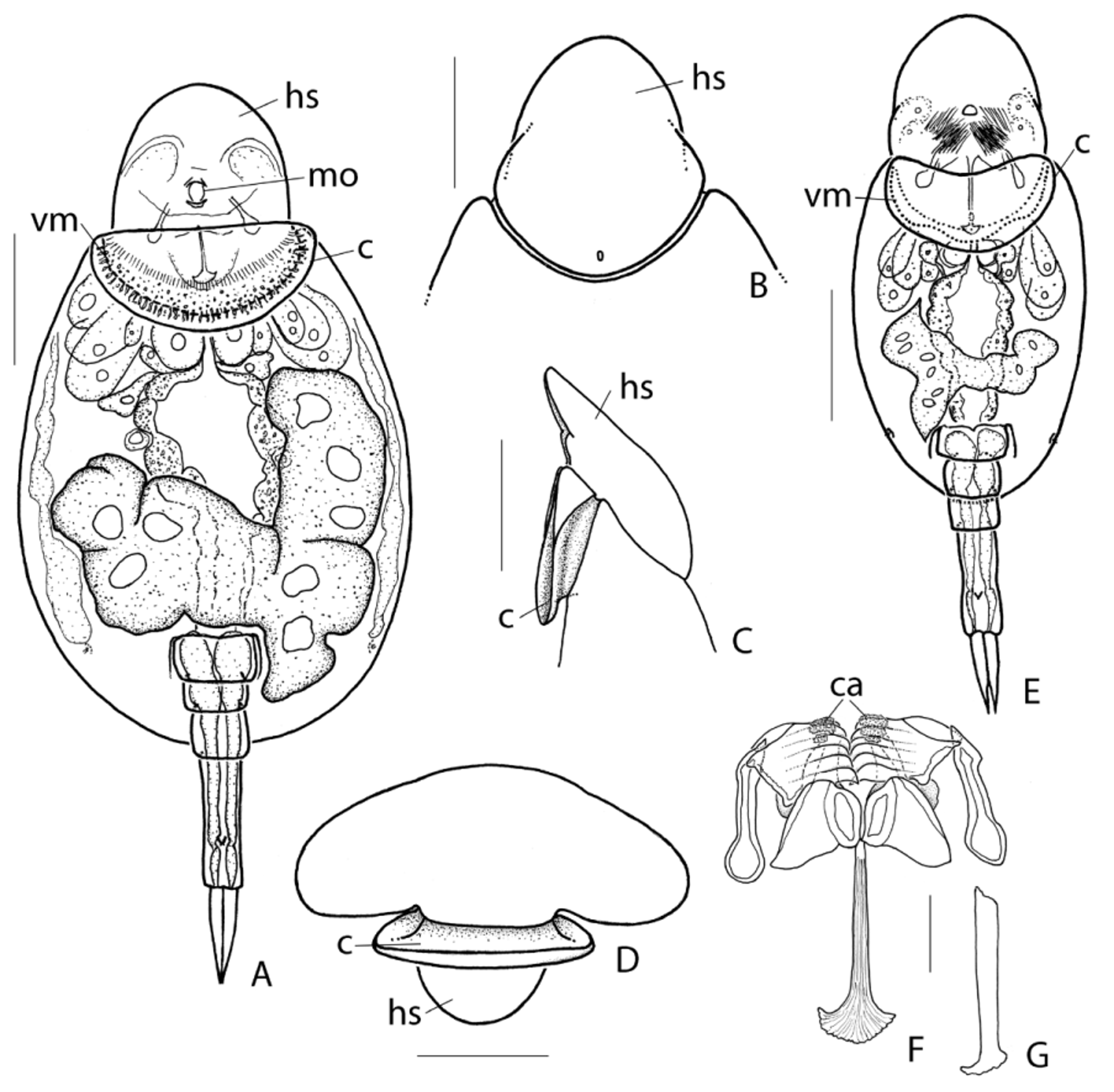
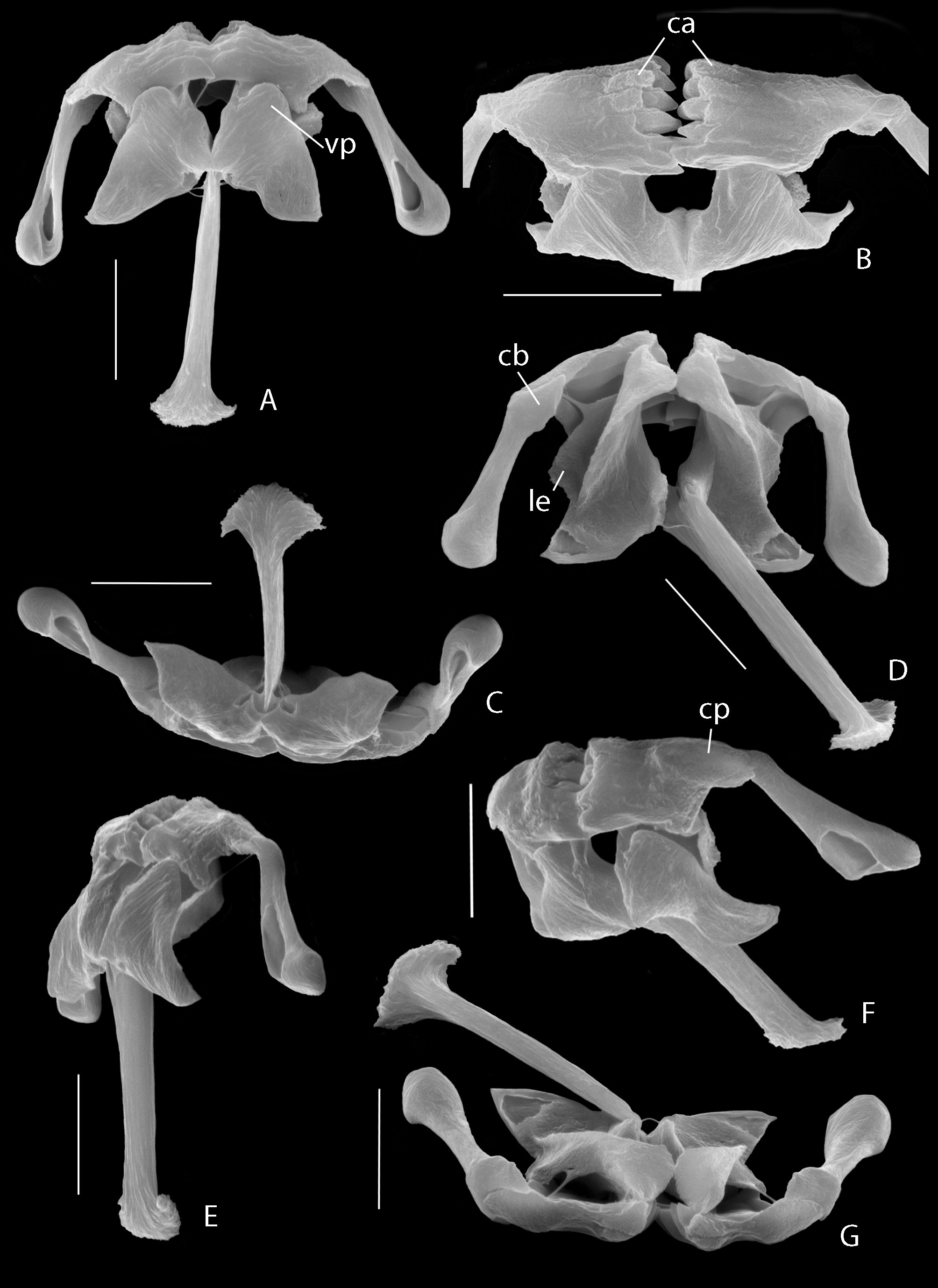
( Figs 1−3 View FIGURE 1 View FIGURE 2 View FIGURE 3 )
Diagnosis. Loricate. Body outline oval in dorsal view, moderately compressed dorso-ventrally. Head with ventral auricular cotyle and dorsal non-retractile shield. Distal foot pseudosegment long, with two toes; no ventro-lateral spurs. Trophi specialized virgate, symmetrical; unci a massive plate composed of 5−6 fused unci teeth; dorsal proximal corner of unci fused to manubria forming immobile joint; head of manubria forked, cauda rounded with opening.
Material examined. 33 females of which 22 adult and 11 juvenile, all from type locality.
Holotype. An adult female in a permanent glycerine glass slide mount deposited in the Royal Belgian Institute of Natural Sciences (R.B.I.N.S.), Brussels, Belgium, IG. 3327, RIR. 262.
Paratypes. An adult (IG. 33271, RIR. 263) and juvenile (IG. 33271, RIR. 264) female in R.B.I.N.S.; 7 stubs each with trophi preparation for SEM and 3 slide preparations in glycerine-gelatine for light microscopy (LM) in the Department of Biology, University of Antwerp.
Type locality. Sariseki Wetland ( 36° 40’ 56.29’’ N, 36° 12’ 30.81’’ E), Iskenderun , province of Hatay, Turkey. GoogleMaps
Etymology. The species name is a latinization of the type locality, Iskenderun, as a noun in the genitive case.
Description of female ( Figs 1−3 View FIGURE 1 View FIGURE 2 View FIGURE 3 ). Loricate. Body outline oval in dorsal view. Head relatively small, offset from trunk by distinct neckfold, covered by non retractile suboval shield; latero-ventral margins of shield near distal third with small incision between two plate-shaped extensions and couple of shallow indentations; head shield can be tilted downward at right angle. Dorsal antenna in shallow elliptic depression near posterior margin of head. Cotyle auricular ( Figs 1 View FIGURE 1 B −D, 2A, C −E: c), hollow, anterior margin almost straight to weakly concave, posterior margin arched and weakly recurved dorsally; integument with series of thickenings: from the outside inwards a series of coarse irregular and radially placed stripes, a field with irregular spots, and a series of faint radially placed stripes ( Figs 1 View FIGURE 1 B, 2A). Mouth opening ventral, slightly protruding, at distal margin of cotyle. Corona reduced. Trunk oval in dorsal view, moderately compressed dorso-ventrally, in cross-sectional view broadly arched dorsally, flattened ventrally with weakly elevated narrow median zone delimited by very shallow longitudinal furrows; lateral margins broadly rounded; antero-dorsal margin of trunk concave, joining proximal margin of head shield; antero-ventral margin ( Figs 2 View FIGURE 2 A, E: vm) broad, smoothly concave, or with very shallow broad median sinus. Lateral antennae small, near distal fifth of trunk. Foot opening ventral, subdistal, a broad inverted U-shaped slit. Foot long, composed of four pseudosegments; distal pseudosegment longest, almost as long as the three proximal ones, cylindrical with dorsal sensory pit near distal third, bearing two equal toes; the other pseudosegments of similar length; toes almost straight, elongate conical with weakly offset tips; toes often pointing dorsally in fixed material. Eyespot(s) absent. Brain fairly large, saccate. Oesophagus short; stomach large, continuing into intestine without distinct constriction. Anterior part of trunk with huge compacted mass of large and smaller cells of which it was impossible to decide whether they belong to the coronal, brain or mastax system. Gastric glands small, stalks medium long. Bladder not observed. Pedal glands in proximal foot pseudosegments with broad long ducts ending in small reservoirs in distal part of distal pseudosegment. Vitellarium grossly t-shaped, with longitudinal lobe in left or right half of trunk and a transversally projecting lobe.
Immature females ( Fig. 2 View FIGURE 2 E) show a proportionally shorter and narrower trunk than the adult ones; the anteroventral margin of the trunk often with shallow, rounded median sinus more pronounced than in adults ( Fig. 2 View FIGURE 2 E: vm).
Trophi ( Figs 2 View FIGURE 2 F, G, 3) specialized virgate, symmetrical, unci fused to manubria into immobile joint, mallei and incus strongly connected by ligaments.
Rami triangular, forming angle of about 140° with fulcrum; more or less acute, fairly broad postero-lateral alulae; distal part of rami bent upward and inward dorsally; the strongly inward bent apical part plate-shaped with rounded, weakly denticulate inner margin; medio-dorsally from these apical plates is a more or less acute tip followed by a dorsal inwardly projecting rounded expansion; a lamellar expansion ( Fig. 3 View FIGURE 3 D: le) projects laterally at bend of distal rami part, showing ligament connected with proximal inner dorsal margin of unci ( Fig. 3 View FIGURE 3 D); large and rounded shallow ventral processes ( Fig. 3 View FIGURE 3 A: vp), forming joint with rami wherein ventral margin of unci fits; subbasifenestrae fairly small ( Fig. 2 View FIGURE 2 C); a small, single ventro-caudal rami opening at proximal end of fulcrum; a dorsal opening at base of alulae. Fulcrum long, c. 1.5 times ramus length, in dorsal view gradually and weakly widening to end more or less abruptly in a broad fan-shaped and dorsally recurved expansion; in lateral view rather shallow, slightly decreasing in height distally. Unci massive, composed of 5 (left) to 6 (right) almost equally strong teeth fused into plate; plate parallel-sided with oblique proximal margin; proximal margin with free short expansion at ventral corner and large dorsal corner; dorsal uncus corner fused into immobile joint with lateral branch of head of manubria, and both the lateral branch and uncus corner outwards fused to an elliptical connecting-platelet ( Fig. 3 View FIGURE 3 F: cp); angle between unci and manubria fixed at about 125° in both mallei; outer surface of uppermost 2−3 unci teeth with distinct callosities prior to tip ( Fig. 2 View FIGURE 2 F, 3B: ca); inner margin of unci teeth a rather acute rim, tip weakly hook-shaped; outer surface of unci teeth with longitudinal sulcus mostly filled with amorphous sclerite material; ventral uncus tooth with thin appressed preuncus. Manubria stout, partially twisted around axis; head bifurcate with stout dorsal and smaller lateral conical branch ( Fig. 3 View FIGURE 3 D: cb), lateral branch fused to dorsal corner of unci and elliptical sclerite; cauda club-shaped, distally rounded with oval latero-ventral opening.
Trophi of immature females fully developed.
Measurements. Adult female, N=10, [values for a juvenile of 270 µm total length]. Total length: 293–372 (mean 349) µm; head shield, length × width: 80–108 × 67–87 (mean 92 × 75) µm [80 × 59 µm]; trunk, length × width: 195–242 × 152–183 (mean 210 × 164) µm [133 × 89 µm]; trunk, height: 62–75 (mean 68) µm; foot: 115– 138 (mean 125) µm [114 µm]; distal pseudosegment: 50–53 (mean 52) µm [44 µm]; toe: 32–42 (mean 35) µm [32 µm]; cotyle, width × length: 80–100 × 37–57 (mean 87 × 44) µm [65 × 38 µm]. Trophi: ramus: 17.9–19.8 (mean 18.5) µm (N=6); fulcrum: 21.1–26.0 (mean 23.2) µm (N=8); uncus: ~12.0–14.2 (mean ~12.8) µm (N=16); manubrium: 15.9–19.0 (mean 17.9) µm (N=10); elliptical sclerite: 6.9–7.7 × 2.7–3.5 (mean 7.1 × 3.3) µm (N=4); angle between uncus and manubrium: 110–140° (mean 125°) (N=16).
Distribution and ecology. The species has to date only been found at the type locality among littoral vegetation at a depth of about 1m. It was present during all seasons.
Examination of the gut content revealed frustules of epiphytic diatoms, mostly Cocconeis placentula Ehrenberg, 1838 and to a lesser extent Fragilaria spp., indicating that C. iskenderunensis n. sp. feeds on diatoms.
Several animals showed an epibiotic fungus attached at the distal end of the trunk and at the base of the foot. Fungi have also been reported on the surface of the body in other rotifers, e.g. the bdelloids Anomopus telphusae Piovanelli, 1903 and Embata parasitica (Giglioli, 1863) by Fontaneto et al. (2004). The specific location of the fungus on the new species at the distal end of the body, and thus near the cloacal opening, indicates that it takes advantage of the excretion products of its host.
Bdelloidea indeterm.
Cephalodella forficula (Ehrenberg, 1838) C. gibba (Ehrenberg, 1830)
Colurella adriatica Ehrenberg, 1831 C. uncinata (Müller, 1773)
Dicranophorus grandis (Ehrenberg, 1832) Eothinia elongata (Ehrenberg, 1832) Euchlanis dilatata Ehrenberg, 1830 Itura aurita (Ehrenberg, 1830) Lecane bulla (Gosse, 1886)
L. closterocerca (Schmarda, 1859) L. flexilis (Gosse, 1851)
L. stenroosi (Meissner, 1908) Lepadella acuminata (Ehrenberg, 1834) L. amphitropis Harring, 1916 L. ehrenbergii (Petry, 1850)
L. quadricarinata (Stenroos, 1898) L. rhomboides (Gosse, 1886)
L. triptera (Ehrenberg, 1830) Ptygura sp.
Synchaeta pectinata Ehrenberg, 1832 Testudinella patina (Hermann, 1783) Trichocerca elongata (Gosse, 1886) T. porcellus (Gosse, 1851)
Trichotria pocillum (Müller, 1776)
1Nomenclature according proposal of Segers et al. (2012) and Jersabek et al. (2015)
Comments. The new species shares the diagnostic characters cotyle, non-retractile head shield and virgate trophi with Cotylegaleata , supporting placement in this genus. Cotylegaleata iskenderunensis n. sp. is distinguished from C. perplexa by the following outstanding characteristics of the body (characteristics of the latter in parentheses): cotyle auricular (subcircular cup-shaped), outline of trunk oval (rectangular), foot without spurs (two distal ventro-lateral spurs), total length 293-372 µm (212-251 µm). Noticeable differences between the two species are apparent in trophi morphology as well. On the whole, all trophi elements are more strongly build in C. iskenderunensis n. sp.: unci massive, bearing callosities (tiny, lacking callosities), unci fused to manubria (movable), rami robust with fairly broad alulae (slenderer with more pronounced alulae), manubria stout, partially twisted around axis, cauda club-shaped, head bifurcate (slender, rod-shaped with recurved cauda, head narrow with tubular chambers). The characteristic epipharynx, consisting of two relatively long stiletto-shaped elements, present in C. perplexa is absent in C. iskenderunensis n. sp..
Although differences in trophi morphology are striking, we do not consider the shared character states of body morphology homoplasious. It is well known that trophi morphology of congeners with different external morphology can be remarkably similar (e.g. Brachionus: Segers et al. 1993 ), or on the other hand that species with similar habitus (e.g. Trichocerca siamensis and T. uncinata: Segers & Pholpunthin 1997 ) can show marked differences in trophi structure. These differences may be attributed to a different food regime and/or way of food intake. No information is available about the feeding behaviour of C. perplexa , but the species was suggested to be some kind of ectoparasite using its cotyle as a sucker for attachment and food intake, with the head shield providing support preventing collapse of the sucker cavity and deformation of the head under the negative pressure generated by the pumping action of the virgate trophi. Cotylegaleata iskenderunensis n. sp. was found feeding on diatoms of which Cocconeis placentula in particular is known to strongly adhere to the substrate and difficult to detach due to its domed free upper valve. When browsing the substrate using the pumping action of its trophi, its cotyle and head shield may have the same function as in C. perplexa . The callosities at the outer surface of the dorsal part of the unci, and at the same height of the asymmetrical uncus-manubrium joint, are probably strengthened points of attachment for muscles operating the movement of the mallei.
The trophi of Cotylegaleata iskenderunensis n. sp. show some features which appear unique and were hitherto not reported in ploimid monogonont rotifers: the fusion of the well developed unci and manubria into an immobile joint, the shape of the unci and their specific connection with rami and manubria, as well as the lateral lamellar ramus expansion. In the virgate trophi the unci may be composed of several unci teeth (e.g. many Notommata ), or the number of unci teeth can be reduced (e.g. most Cephalodella and Trichocerca ) to a single strong tooth ( Nogrady et al. 1995). When the uncus is composed of several unci teeth the ventral most tooth is normally most strongly developed, and the proximal uncus margin appears connected over its whole width to the head of the manubrium by ligaments, forming a flexible joint; a joint of the ventral uncus margin with a ventral process of the rami is lacking. This clearly contrasts with the unci of C. iskenderunensis n. sp. which show more or less equally developed stout unci teeth, the unci being only connected by their dorsal proximal corner to the manubria, and with their ventral edge interlocking with a furrow formed by a ventral ramus process.
The differences in some of the features of C. perplexa and C. iskenderunensis n. sp. involve the modification of the previous diagnosis of the family and genus. The amended diagnosis is as follows: external morphology bilaterally symmetrical, with small head, large trunk and long foot bearing two toes. Head with ventral cotyle and dorsal stiffened non-retractile head shield. Corona reduced. Trophi specialized, modified virgate.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |