
Chiasmocleis carvalhoi ( Nelson, 1975 )
|
publication ID |
https://doi.org/10.1206/834.1 |
|
persistent identifier |
https://treatment.plazi.org/id/03F8878E-6F58-843B-FD75-FC13721EF9F3 |
|
treatment provided by |
Felipe |
|
scientific name |
Chiasmocleis carvalhoi ( Nelson, 1975 ) |
| status |
|
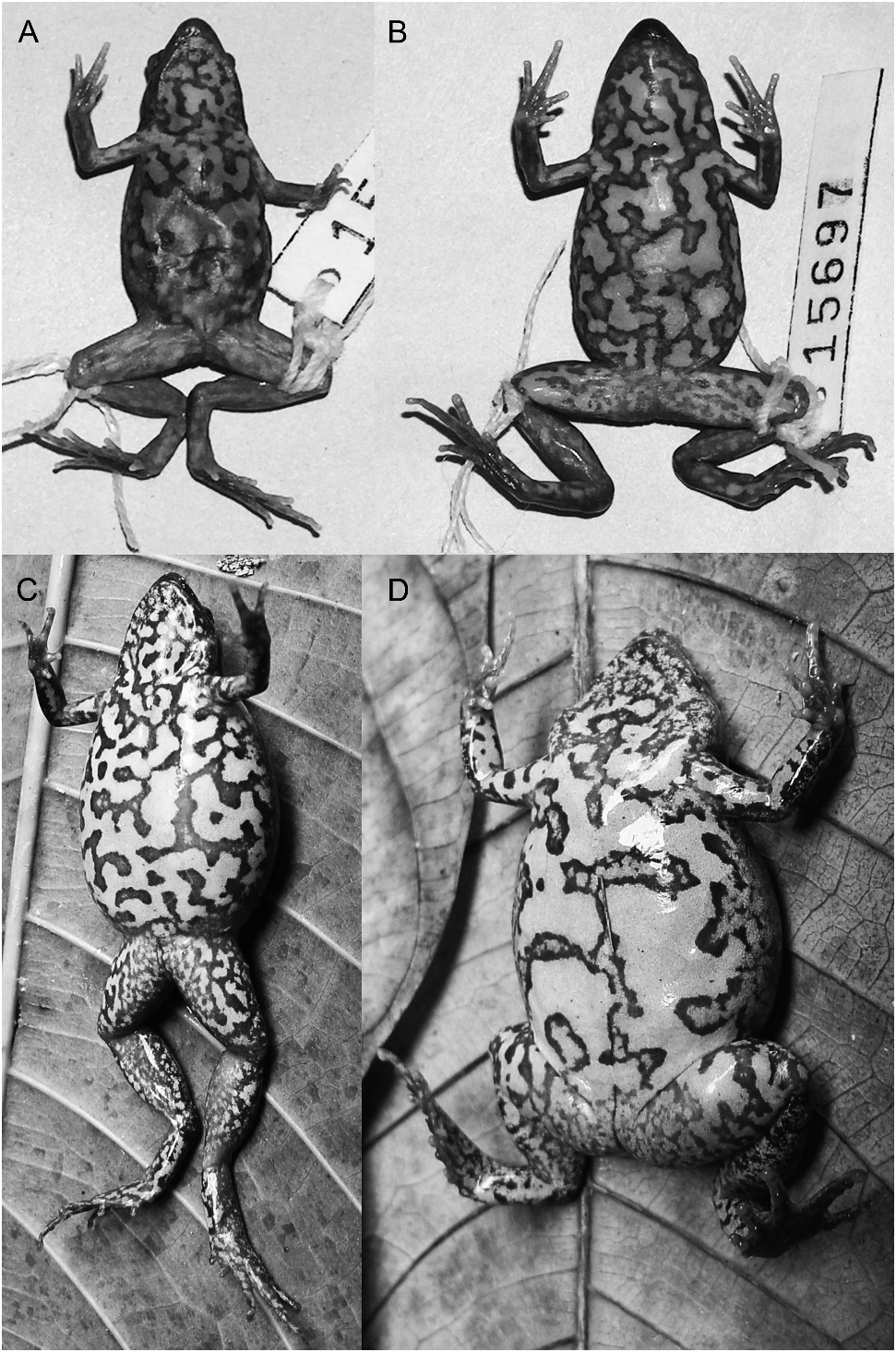
Chiasmocleis carvalhoi ( Nelson, 1975) View in CoL Figures 26 View Fig , plate 5A–F
Syncope carvalhoi: ( Nelson, 1975) View in CoL .
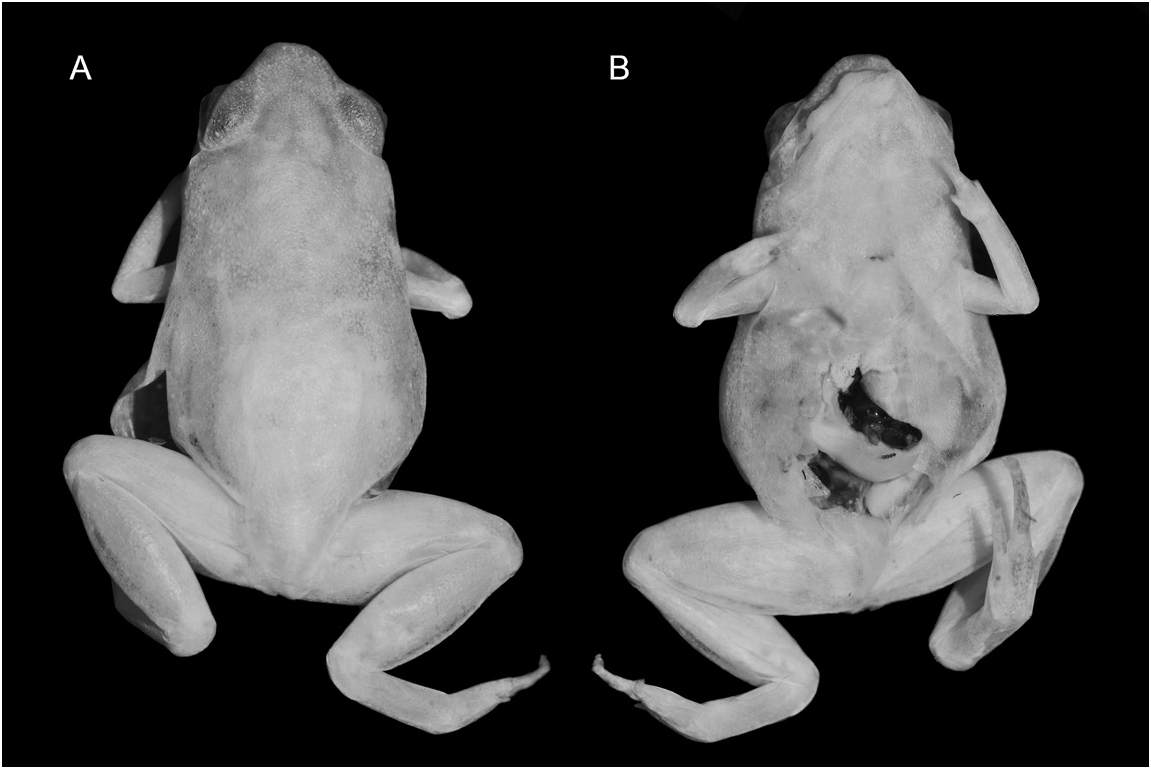
HOLOTYPE: MZUSP 36429; an adult male, in very good state of preservation.
TYPE LOCALITY: Río Ampi-Yacú (5 Ampiyacú), Estirón, Loreto, Peru. No coordinates given in the original. We found at least three different localities in Loreto named Estirón. Because the original explicitly states the specimens were collected along in the Rio Ampiyacú river, we assume the type locality refers to Estirón del Cuzco, for which we provide the following approximate coordinates 03 ° 229S / 72 ° 009W. 10
DIAGNOSIS: A small species for the genus; SVL of the male holotype 9.4 mm ( Nelson, 1975) and 10.9–11.7 mm in the two female paratypes ( N 5 2: Nelson, 1975). Body relatively robust, snout round in dorsal and lateral views. Tympanum is usually not visible. FII–IV much reduced; FI not visible; FIII well developed with subarticular tubercle
10 Coordinates taken from direct observation of satellite images in the interactive software Google Earth, release 6.2.2 for Mac (Google Inc., 2012).
barely visible, tip of FIII pointed; FIV almost not distinguishable; fingers slightly fringed; palmar tubercles not present; relative finger lengths IV,II,III. Four toes visible, TI not visible; toes only slightly fringed, not webbed; tips of toes slightly expanded, especially in TIII–TIV, tips of TIII–TIV pointed. Relative toe lengths II,V,III,IV. Dermal spines absent. No femoral stripe present. In preservative, venter (belly and undersurfaces of thigh) beige with white spots or blotches. In life, dorsum brownish or orangish with darker (brown to dark brown blotches); cream-colored canthal stripe that may end at the shoulder region or extend all the way to the inguinal regions, becoming wider and more irregularly defined posteriorly; belly with large anastomosing cream-colored large blotches (brown area forms an irregularly reticulated pattern).
VARIATION: Number of specimens examined in detail ( N 5 7) is much reduced to present data on variation, but we provide a few remarks on color in life, based on examination of photographs (pl. 5A–F). Two syntopic species, from Umarital, Rio Ampiyacú region, Loreto, Peru (, 30 km from the type locality) vary markedly in dorsal pattern; JMP 1499 (pl. 5C–D) has a more uniform dorsum (brown) and a canthal stripe extending to the shoulder, while JMP 1451 (pl. 5A–B) has dark brown blotches over a brown dorsum and the stripe extends all the way to the inguinal region. Because these two specimens were examined only from photographs, we do not know whether this difference is due to sex or ontogeny or by chance.
CALL AND TADPOLE: Call is unknown. Nelson (1975) reported large unpigmented ovarian eggs for the species suggesting endotrophic larvae (e.g., direct development or nonfeeding tadpoles that develop in water or moist soil). Nothing more is known about its reproductive behavior and, at this point, there is no evidence that a tadpole phase even exists.
12 0.006
11 0.006 0.004
10 0.000 0.006 0.004
9 0.002 0.002 0.007 0.006 bassleri 8 0.044 0.042 0.042 0.044 0.046 Chiasmocleis 7 6 0.000 0.017 0.017 0.029 0.029 0.028 0.028 0.028 0.028 0.029 0.029 0.031 0.031 of
sequences 5 0.035 0.035 0.048 0.035 0.033 0.033 0.035 0.037 7
S
TABLE between 16 4 3 0.037 0.035 0.037 0.026 0.029 0.029 0.026 0.046 0.041 0.007 0.033 0.006 0.031 0.031 0.006 0.007 0.031 0.035 0.002 distances 2 0.007 0.033 0.031 0.029 0.029 0.044 0.004 0.002 0.002 0.007 0.006 pairwise 1 0.039 0.042 0.044 0.042 0.035 0.035 0.041 0.041 0.039 0.039 0.041 0.042 Uncorrected)) 47166 47167)
))
27334) 27764) 27765 17515) 24565)
Locality ALP
14940
(
Amazonas
21576
(
MHNSM
Loreto
MNCNADN
( Loreto
:
Vaupés (MNCNADN (
:
Vaupés MNCNADN
Amazonas
MPEG (
Amazonas
MPEG ()
Pará
28326 MPEG (MTR) (28345
Acre
) MTR (28391
Acre
28402 MTR (
Acre
) ( Orellana
:
QCAZ
:
(QCAZ Orellana
:
Brazil: Peru: Peru
Colombia Colombia: Brazil Brazil:: Brazil: Brazil Brazil:: Brazil Ecuador Ecuador
1 2 3 4 5 6 7 8 9 10 11 12 13
DISTRIBUTION (fig. 18): Known from east Loreto, Peru, and west Colombia (see Acosta-Galvis, 2000, for the Colombian record). Ortega-Andrade and Valencia (2010) provided a record of Chiasmocleis tridactyla (as Syncope tridactyla ) for Lorocachi, Pastaza, Ecuador. We have not examined these specimens, but from the figures in their publication it seems those specimens should be tentatively identified as C. carvalhoi based on reduced number of fingers (three) and toes (four) and ventral color pattern (large white blotches) versus small spots in C. tridactyla . The only ambiguous character is the presence of a tympanum (barely visible) in both specimens reported by the authors (QCAZ 9577, 9520; tympanum absent in the types of C. carvalhoi ).
REMARKS: Two specimens from Rio Nanay, Loreto, Peru, included in de Sá et al. (2012) as Syncope sp. were found nested within our Chiasmocleis carvalhoi clade. These specimens show 0.0 % genetic distance from specimen MNCNADN 27392, collected near Iquitos, Loreto, Peru. Pending examination of a larger series of C. carvalhoi and the vouchers of the specimens sequenced by de Sá et al. (2012), we include those specimens in C. carvalhoi . Genetic distances between all specimens of C. carvalhoi includ- ed in the phylogenetic analysis are given in table 8.
Chiasmocleis devriesi Funk and Cannatella, 2009 View in CoL Figure 15D View Fig , plate 5G–H
HOLOTYPE (figs. 15D, pl. 5G–H): MHNSM 21540; examined only from the photographs in the original publication ( Funk and Cannatella, 2009).
TYPE LOCALITY: Amazon Conservatory for Tropical Studies ( ACTS) Field Station, 65 km NE of Iquitos, 1 km N of the Río Napo, and 1.3 km SSE of the ACTS Field Station (3 ° 159340S, 72 ° 549100W), Departamento Loreto, Peru.
DIAGNOSIS: Since we have not examined the sole specimen of Chiasmocleis devriesi (except from the photographs published in the original description), readers should refer to the original description ( Funk and Cannatella, 2009) for a diagnosis of the species.
VARIATION: Known only from the female holotype.
CALL AND TADPOLES: Call and tadpoles are unknown.
REMARKS: Funk and Cannatella (2009) described the species on the basis of a single female specimen, which renders any evaluation of morphological variation unattainable. Additionally, many diagnostic characters in Chiasmocleis are present only or are more pronounced in males (i.e., webbing, fringes of digits, and dermal spines). The ventral pattern of the holotype (fig. 15D, pl. 5H) is similar to some specimens of C. anatipes , C. avilapiresae , and C. ventrimaculata and cannot be unambiguously set apart from those on the basis of this character. Chiasmocleis devriesi differs from all species by its larger size (SVL 42.4 mm in C. devriesi ; rivaled only by C. avilapiresae , with a maximum recorded SVL, according to our sample, of 37.8 mm) and its moderately pointed snout. The absence of a femoral line further differentiates C. devriesi from C. avilapiresae (femoral line present). From our phylogenetic analysis, it seems clear that the relationship of the sole specimen of C. devriesi lies with populations assigned to C. anatipes and not with C.
TABLE 8 Uncorrected pairwise distances between 16S sequences of Chiasmocleis carvalhoi
avilapiresae or C. ventrimaculata (fig. 8). The holotype is related to a couple of specimens from Orellana, Ecuador, forming a wellsupported sister clade to another three specimens, unambiguously assigned to C. anatipes . Because the specimens from Orellana are juveniles they cannot be unambiguously assigned to C. anatipes nor do they fit precisely the diagnosis of C. devriesi . For the purpose of our phylogenetic analysis, we have labeled the specimens from Orellana as C. anatipes , but they clustered with the holotype of C. devriesi . There is zero genetic distance in the 16S between the sole specimen of C. devriesi and each of the specimens of C. anatipes included in our analysis (see table 4). We have examined only the holotype of C. devriesi from photographs published in the original description ( Funk and Cannatella, 2009; also reproduced here as fig. 15D and pl. 5H). Due to the lack of evidence supporting or falsifying the status of C. devriesi as a valid taxon, we chose not to synonymize it with C. anatipes , but highlight that this issue deserves close attention in the future. For the phylogenetic analysis, for example, only the 16S fragment is present for the holotype of C. devriesi (which shows no genetic differentiation among populations; table 4). Given the phylogenetic proximity of these populations, it would be wise to expand character sampling for these taxa in the future.
DISTRIBUTION (fig. 27): Known only from the type locality.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Chiasmocleis carvalhoi ( Nelson, 1975 )
| Peloso, Pedro L. V., Sturaro, Marcelo José, Forlani, Mauricio C., Gaucher, Philippe, Motta, Ana Paula & Wheeler, Ward C. 2014 |
Chiasmocleis devriesi
| Funk and Cannatella 2009 |
Syncope carvalhoi : ( Nelson, 1975 )
| : (Nelson 1975 |