
Macrosiphum glawatz Jensen, 2020
|
publication ID |
https://doi.org/10.4289/0013-8797.122.1.81 |
|
DOI |
https://doi.org/10.5281/zenodo.3728803 |
|
persistent identifier |
https://treatment.plazi.org/id/03F88780-AE76-FFFE-FF76-8EDDFB16FD2A |
|
treatment provided by |
Valdenar |
|
scientific name |
Macrosiphum glawatz Jensen |
| status |
sp. nov. |
new species
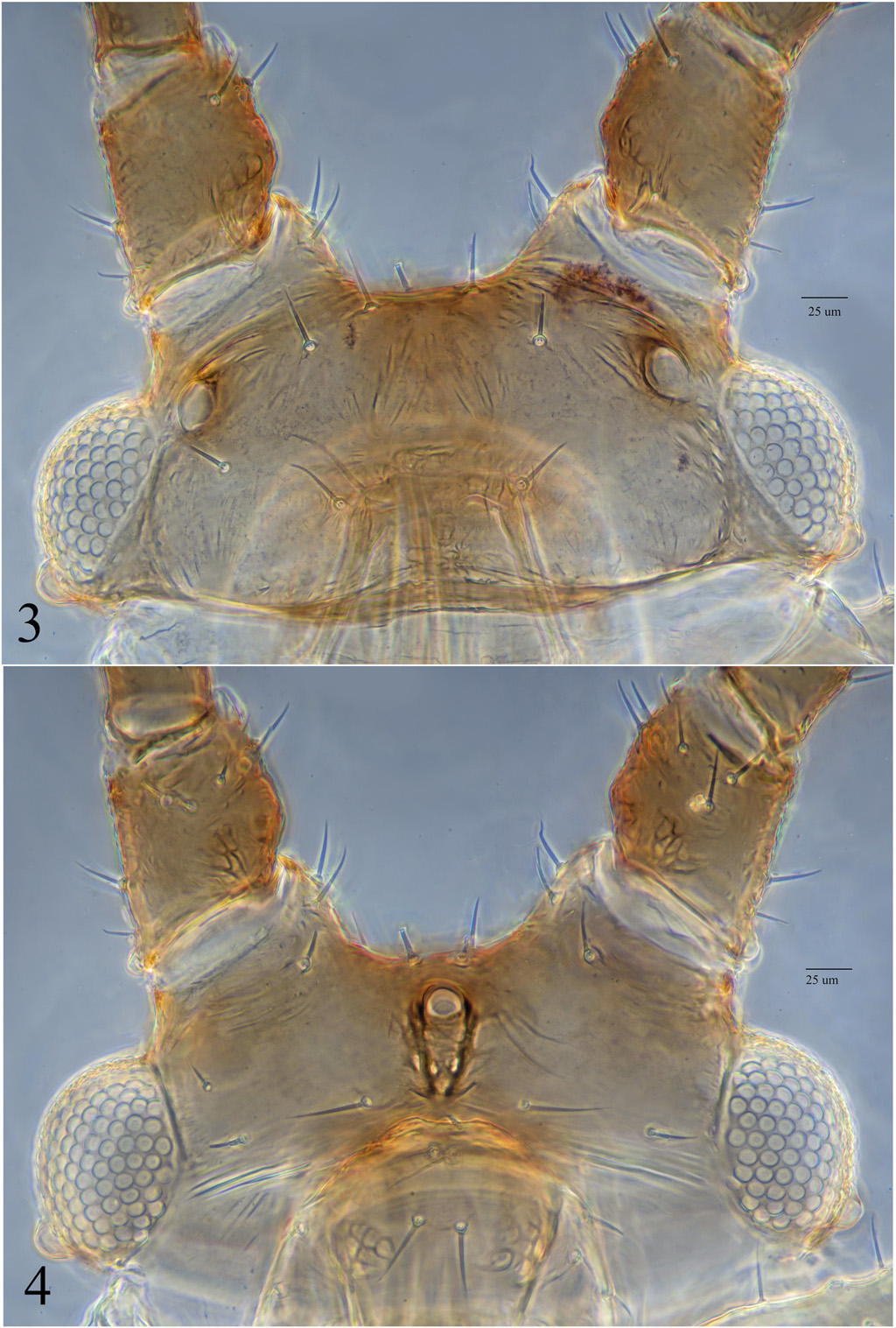
http://zoobank.org/ C8514E8C-7F79- 425F-A87D-7AFE7C4203F8 ( Figs. 1–12 View Figs View Figs View Figs View Figs )
Apterae.— Color: When alive, medium green, pinkish red, or red mottled
with green, antennae, tips of tibiae, tarsi, and apical portions of siphunculi brown. When macerated, antennae with segment III gradually becoming light brown toward apex, extreme apex darker brown ( Fig. 5 View Figs ); a.s. IV dusky to light brown at extreme base, then pale until gradually darkening toward apex, with about apical ¼ brown, but sometimes entire segment brown; a.s. V and VI light brown to brown, except a.s. V sometimes with paler middle section. Head pale to slightly dusky. Rostral segment III dusky to light brown, u.r.s. brown. Femora entirely pale to faintly dusky dorsoapically. Tibiae faintly dusky at base, paler in the middle, then darker to dark brown at apex. Tarsi dark brown. Siphunculi entirely pale to dusky or light brown over apical ½ to 4/5, reticulated part noticeably darker ( Fig. 7 View Figs ). Other body parts pale.
Morphology: Measurements of body, appendages, setae, etc. see Table 1 View Table 1 . Antennae with segment I ( Figs. 1-2 View Figs ) smooth, sometimes with a few broad spinules apically on medial side; a.s. II with strong cluster of ornamentation medially, which extends ventrally and dorsally; a.s. III ( Fig. 5 View Figs ) more or less evenly covered in faint imbrications, sensoria scattered over about ½ to full length of segment; a.s. IV-VI with imbrications becoming more prominent; antennal setae blunt to slightly capitate. Head with antennal tubercles as in Fig. 1 View Figs , small specimens with antennal tubercles lower; 1 or 2 faint spinal tubercles rarely present; dorsum of head sometimes slightly wrinkled, especially area immediately anterio-medial to eyes; dorsal setae blunt, tips slightly less prominent than on antennae; ventral and dorsal setae more or less the same shape; variable patch of spinules present ventrally between mouthparts and antennal tubercles, Fig. 2 a View Figs typical example, some specimens with more spinules near the antennal bases; a small patch of spinules usually present posterio-ventrally on each side of mouthparts. Rostrum reaching metacoxae; segment II with stylet groove spinulated, segments III and u.r.s. without ornamentation; setae on rostrum pointed; u.r.s. long, triangular, pointed apically ( Table 1 View Table 1 , Fig. 11 View Figs ). Prothorax rarely with faint lateral tubercles. Thorax with dorsum slightly wrinkled, setae blunt; setae on coxae and trochanters pointed; femora with setae blunt, no ventral ones dramatically longer than others, ornamentation more or less absent to spinules and small imbrications over apical ~½ anteriorly, this ornamentation more extensive on metafemur. Tibiae with setae blunt, some ventral ones visibly thicker than thickest dorsal ones, metatibia setae not noticeably longer than corresponding setae on pro- and mesotibiae. Tarsi ( Fig. 12 View Figs ) with 3 setae on segment I, protarsal II with 2 or 3 pairs of dorsal setae, imbrications smooth, rarely with faint spinules. Abdomen with tergum membranous, smooth; dorsal setae blunt to slightly capitate, ventral setae pointed; segments III-VI rarely with small indistinct lateral tubercles. Siphunculi ( Fig. 7 View Figs ) with about 5 rows of apical polygonal reticulation, strong ribbed or spinulate imbrications more basally, becoming smaller and more ribbed or spinulate on basal ~¼. Cauda ( Figs. 9-10 View Figs ) with setae more or less paired; dorsal ornamentation of ribbed imbrications about 8-10 across width at middle; ventral ornamentation of spinules in groups of 2-4. Tergites VII and VIII without spinal tubercles. Subgenital plate with row of postero-marginal setae, 1 pair of anterior setae, and 1 to ~15 setae on its disk. Middle pair gonapophyses fused.
Alatae.— Color: When alive, medium green, pinkish red, or yellowish green, thoracic plates and appendages light brown to brown. When macerated, antennae with a.s. I dusky to brown, a.s. II often a little darker, remainder of antenna brown except extreme base of a.s. III. Head pale to dusky, area around each ocellus brown ( Fig. 3 View Figs ). Rostrum with segment III brown, u.r.s. dark brown. Dorsal and ventral thoracic plates light brown. Femora pale at base, gradually darkening toward apex, dark brown over apical ~¼. Tibiae and tarsi brown, tibiae sometimes lighter brown on basal~4/5. Abdomen with lateral sclerites on segments II-IV faintly dusky. Siphunculi brown beyond extreme base ( Fig. 8 View Figs ). Other body parts pale.
Morphology: Measurements of body, appendages, setae, etc. see Table 1 View Table 1 . Antennae with segment III ( Fig. 6 View Figs ) with sensoria scattered over full length of segment; a.s. IV-VI with imbrications more or less uniformly prominent. Head with antennal tubercles more or less as in Fig. 3 View Figs ; spinal tubercles absent; dorsum of head more or less smooth; a few spinules present ventrally between mouthparts and antennal tubercles, Fig. 4 a View Figs typical example, some specimens without visible spinules; a small patch of spinules usually present posterioventrally on each side of mouthparts. Rostrum almost reaching metacoxae. Prothorax sometimes with faint lateral tubercles. Thorax with thoracic plates normal, wings full-sized, wing venation normal with media twice-branched; femora with ornamentation more or less absent to spinules and small imbrications over apical ~½ anteriorly, ventrally, and a little posteriorly this ornamentation more prominent than in aptera. Abdomen with segments II-IV with lateral sclerites with indistinct borders and with more or less uniform spinulation and usually very small lateral tubercles. Siphunculi ( Fig. 8 View Figs ) with about 7 rows of apical polygonal reticulation. Cauda narrower than in aptera and with constrictions between setal pairs. Otherwise more or less as in aptera.
Type material.— Holotype aptera: Slide number AJ8518, U.S.A., Oregon, Lake County, Warner Mts. , Can Spring , ex Potentilla gracilis , 10 June 2016, A.S. Jensen (2 USNM). The holotype slide has the USNM database barcode number: USNMENT 01021470.
Paratype apterae: Slide number AJ663, U.S.A., Oregon, Umatilla County, Bridge Creek wildlife area, ex Potentilla gracilis , 16 May 1992, A.S. Jensen (1 ASJ); Slide number AJ10161, U.S.A., Oregon, Deschutes County, near Black Butte, ex Potentilla gracilis , 30 May 2018, A.S. Jensen (1 ASJ); Slide numbers AJ7234, AJ7235, U.S.A., Oregon, Grant County, Umatilla National Forest, Rd. 10 near Dale , ex Potentilla gracilis , 31 May 2014, A.S. Jensen (4 ASJ, BMNH); Slide numbers AJ9210, AJ9211, U.S.A., Oregon, Union County, Wallowa Whitman National Forest, Fly Creek , ex Potentilla gracilis , 3 June 2017, A.S. Jensen (7 CNC, ASJ); Slide number AJ8518, U.S.A., Oregon, Lake County, Warner Mts., Can Spring , ex Potentilla gracilis , 10 June 2016, A.S. Jensen (2 ASJ); Slide numbers AJ9255, AJ9256, U.S.A., Oregon, Lake County, West side of Lakeview valley, 10 June 2017, A.S. Jensen (4 USNM, CNC); Slide number AJ10222, U.S.A., Oregon, Lake County, West side of Lakeview valley, ex Potentilla gracilis , 14 June 2018, A.S. Jensen (4 ASJ); Slide numbers AJ4852, AJ4853, AJ4854, U.S.A., Washington, Whitman County, Rock Creek – Towel Falls , ex Potentilla gracilis , 11 vi 2011, A.S. Jensen (13 ASJ); Slide number AJ8487, U.S.A., Oregon, Lake County, Rd. 3780 W. of Lakeview valley, ex Potentilla gracilis , 17 vi 2016, A.S. Jensen (3 ASJ); Slide numbers AJ5678, AJ5679, AJ5680, U.S.A., Oregon, Union County, I-84 Frontage Rd. Exit, ex Potentilla gracilis , 17 June 2012, A.S. Jensen (9 ASJ); Slide number AJ8546, U.S.A., Oregon, Lake County, Bauers Creek Rd. north of Lakeview, ex Potentilla gracilis , 24 June 2016, A.S. Jensen (1 BMNH); Slide number AJ10283, U.S.A., Oregon, Lake County, Fremont N.F., Winter Rim area, ex Potentilla gracilis , 24 June 2018, A.S. Jensen (1 ASJ); Slide number AJ8548, U.S.A., Oregon, Lake County, Radio towers area above Lakeview , ex Potentilla gracilis , 26 June 2016, A.S. Jensen (2 ASJ); Slide numbers AJ5687, U.S.A., Oregon, Union County, I-84 Frontage Rd. Exit, ex Potentilla gracilis , 16 July 2012, A.S. Jensen (5 ASJ); Slide numbers AJ9579, AJ9580, AJ9581, U.S.A., Oregon, Lake County, Fremont N.F., Hannan Trail along Sycan River , ex Potentilla gracilis , 29 July 2017, A.S. Jensen (11 ASJ); Slide number AJ8796, U.S.A., Oregon, Lake County, Warner Mts. near Camas Valley, ex Potentilla gracilis , 7 August 2016, A.S. Jensen (2 ASJ); Slide number AJ8790, U.S.A., Oregon, Lake County, Warner Mts. near Rogger Meadow, ex Potentilla gracilis , 7 August 2016, A.S. Jensen (3 ASJ); Slide number AJ8839, U.S.A., Oregon, Lake County, North Warners, Mud Creek Camp , ex Potentilla gracilis , 15 August 2016, A.S. Jensen (1 ASJ); Slide number AJ8926, U.S.A., Oregon, Lake County, North Warners, Can Spring , ex Potentilla gracilis , 27 August 2016, A.S. Jensen (4 ASJ); Slide number AJ6760, U.S.A., Oregon, Union County, I-84 Frontage Rd. near North Powder, ex Potentilla gracilis , 23 August 2013, A.S. Jensen (1 ASJ).
Paratype alatae: Slide number AJ663, U.S.A., Oregon, Umatilla County, Bridge Creek wildlife area, ex Potentilla gracilis , 16 May 1992, A.S. Jensen (1 ASJ); Slide number AJ10161, U.S.A., Oregon, Deschutes County, near Black Butte, ex Potentilla gracilis , 30 May 2018, A.S. Jensen (1 ASJ); Slide number AJ7234, U.S.A., Oregon, Grant County, Umatilla National Forest, Rd. 10 near Dale, ex Potentilla gracilis , 31 May 2014, A.S. Jensen (1 ASJ); Slide numbers AJ9255, AJ9256, U.S.A., Oregon, Lake County, West side of Lakeview valley, 10 June 2017, A.S. Jensen (4 USNM, CNC); Slide number AJ4854, U.S.A., Washington, Whitman County, Rock Creek – Towel Falls , ex Potentilla gracilis , 11 vi 2011, A.S. Jensen (1 ASJ); Slide number AJ5679, U.S.A., Oregon, Union County, I-84 Frontage Rd. Exit, ex Potentilla gracilis , 17 June 2012, A.S. Jensen (1 ASJ); Slide number AJ8546, U.S.A., Oregon, Lake County, Bauers Creek Rd. north of Lakeview, ex Potentilla gracilis , 24 June 2016, A.S. Jensen (1 BMNH); Slide number AJ10283, U.S.A., Oregon, Lake County, Fremont N.F., Winter Rim area, ex Potentilla gracilis , 24 June 2018, A.S. Jensen (1 ASJ).
Slides containing all type specimens are clearly marked and highlighted with red ink.
Etymology.—This species is named in honor of Claus and Helga Glawatz, parents of Gina Rone (see also the Acknowledgments). Despite living in Germany and speaking little English, Claus and Helga have seen more of the United States than almost anyone, including much of the habitat of M. glawatz . For the purposes of the rules of nomenclature, the name should be considered a random collection of letters without gender.
Biology and distribution.—This aphid lives throughout the growing season on Potentilla gracilis in meadows and edges of dry forest. Much collecting effort over the years has not yet yielded any sex- uales, and none of the apterae collected so far can be confidently labeled as fundatrices. Despite these facts, it is extremely likely that M. glawatz is monoecious holocyclic on P. gracilis . Evidence supporting this idea are the collections of apterae of this species in early spring. At the time of these collections, such as 16 May or 30 May, P. gracilis in eastern Oregon is just entering its pre-flowering growth stage and is in perfect condition for aphid reproduction. At the same time, shrubs and trees that might be host to an undiscovered primary host life stage are just beginning growth and would be very unlikely to support two spring generations of an aphid that then migrates to P. gracilis by early May. The lack of success in finding sexuales is related to two primary issues: the phenology and growth habit of P. gracilis and the nearly ubiquitous cattle grazing of relevant habitats. The life cycle of P. gracilis is similar to the hosts of two other recently described Macrosiphum species from similar habitats, Macrosiphum dewsler Jensen on Linum lewisii Pursh and Macrosiphum garyreed Jensen on Geum triflorum Pursh ( Jensen 2017) , in that it overwinters as a basal rosette and begins growing in early spring, flowers during the summer, and then survives the late summer drought as a basal rosette with very small leaves. It is likely that the sexuales of M. glawatz develop on these late summer rosette leaves but searching for them is challenging in part due to heavy cattle grazing and trampling that decimates M. glawatz habitat every summer. If a good site could be located that is free of grazing, M. glawatz could be followed from early spring through its presumed sexual stage in that site, but we have not been lucky enough to find such a place. Macrosiphum glawatz is unusual among Macrosiphum and similar genera in that it is almost always tended by ants. The only other western North American Macrosiphum that is usually ant-tended is Macrosiphum rhamni (Clarke) when it is feeding on its secondary host, Pteridium aquilinum (L.) Kuhn. Only a few samples of the ants tending M. glawatz have been collected, but these all belong to the genus Formica L. (identified to genus using Hölldobler and Wilson 1990).
Macrosiphum glawatz is so far known only from Oregon and Washington east of the Cascade Mountains, but almost certainly occurs in nearby portions of California and Idaho.
Remarks.—Study of Macrosiphum living on Potentilla forced us to examine the identities of two poorly understood nominal species: Macrosiphum potentillae (Oestlund) and Macrosiphum potentillicaulis Miller. Oestlund (1887) described M. potentillae from an undisclosed location and date(s) in Minnesota based on an unknown number of alate viviparous specimens collected from Potentilla anserina L. Oestlund’s description is very brief (less than 8 lines of text), offering almost no useful characters. Cook (1982) reported that there were 24 specimens at UMN collected by Oestlund in 1886 and that he, Cook, designated a lectotype from among them. In February of 2019 we requested a loan of the M. potentillae material on hand at the UMN collection. Among the 28 slides received, 24 were from Oestlund’s collection(s) in 1886 in Minneapolis, Minnesota. There is no specific collection date on the slides, and the host is shown as “ Potentilla .” These 24 slides each contain one specimen, all but one cleared and remounted into balsam from Oestlund’s original material: 5 nymphs, 10 badly over-cleared (almost invisible) alatae, 6 damaged but not terribly overcleared alatae, one aptera in good condi- tion except for the antennae (segments III-VI entirely missing except the basal ¼ of one segment III), and one alate male. One slide containing an intact, well-preserved alata is clearly marked as lectotype. Examination shows that Oestlund’s species is distinct from M. glawatz . In the apterae, the abdominal dorsum of M. potentillae is sclerotized and faintly brownish in color with clear areas around the bases of the siphunculi, the ventral surface of the head has substantially more spinulation than in M. glawatz , and a.s. I and II are heavily ornamented with imbrications and spinulation. Both alatae and apterae have cauda shorter than M. glawatz (about 220 µm vs. 220–440 µm in M. glawatz ) and have setae on a.s. III short, about equal to or less than the basal width of a.s. III. Examination of this type material led the first author to identify as M. potentillae three apterae he collected near the Saguenay River in Quebec, Canada in August of 2013. It is noteworthy that M. potentillae is very similar to, and possibly the same as, Macrosiphum manitobense Robinson , which has been suspected since its original description to be heteroecious with an unknown summer host ( Robinson 1965). The available alate viviparae of our material of M. manitobense and M. potentillae are basically indistinguishable. Our secondgeneration apterous viviparae on Cornus from western Oregon however, differ slightly from the apterae from Potentilla in Minnesota and Quebec. These differences could be due to the specimens having intermediate morphology between fundatrices and summer secondary host apterae, or they could point toward there being two similar species involved. With this close similarity discovered, further research should include study of Potentilla species available in the same areas where M. manitobense is found on Cornus . Also, as is evident from the key presented below, modified from Blackman and Eastop (2019), M. potentillae has some similarity to species currently placed in Anthracosiphon Hille Ris Lambers. This relationship should be explored further.
The situation with M. potentillicaulis , described from Potentilla gracilis (as Potentilla bakeri ) from northern Idaho, was also interesting and challenging to unravel. Looking at the description and figures provided by Miller (1933) it is obvious that his species is not the same as M. glawatz , differences including the very long cauda and completely black siphunculi mentioned in Miller’s description. It is still important, however, to resolve the uncertainty around the name M. potentillicaulis . Jensen (1998) published a key to “North American Macrosiphum whose apterous viviparae have black siphunculi and pale, membranous terga” and in that key M. potentillicaulis was not differentiated from Macrosiphum valerianae (Clarke) (as Macrosiphum kiowanepus (Hottes) and Macrosiphum subarcticum Robinson ) due to the lack of any characters to separate them. Subsequently, Eastop and Blackman (2005) declared M. subarcticum a junior synonym of M. valerianae and Jensen (2012) showed M. kiowanepus was a junior synonym of M. valerianae . During preparation of the latter paper the first author considered deciding on the validity of M. potentillicaulis , but none of the available material from Miller was labeled as type. Given the fact that M. valerianae is commonly found on Potentilla , it was decided that any material not labeled as type of M. potentillicaulis could have been M. valerianae subsequently misidentified as M. potentillicaulis . The name was therefore retained for future study. In winter of 2016 the first author once again examined the Miller material in the USNM and found a slide with the code number referred to by Miller (1933) as being the “ holotype:” Slide “#S 35-32” ( Fig. 25 View Fig ). This slide had no other markings indicating that it was a type slide, and in fact its identity as type had been overlooked in a recent comprehensive inventory of USNM’s type specimens (as a result of this work, this type slide now has the USNM barcode number: USNMENT 01021471). Examination of the specimens on Miller’s holotype slide (2 alate viviparae and one large nymph) makes it clear that they should be considered M. valerianae and that Macrosiphum potentillicaulis Miller is a junior synonym of Macrosiphum valerianae (Clarke) , new synonymy.
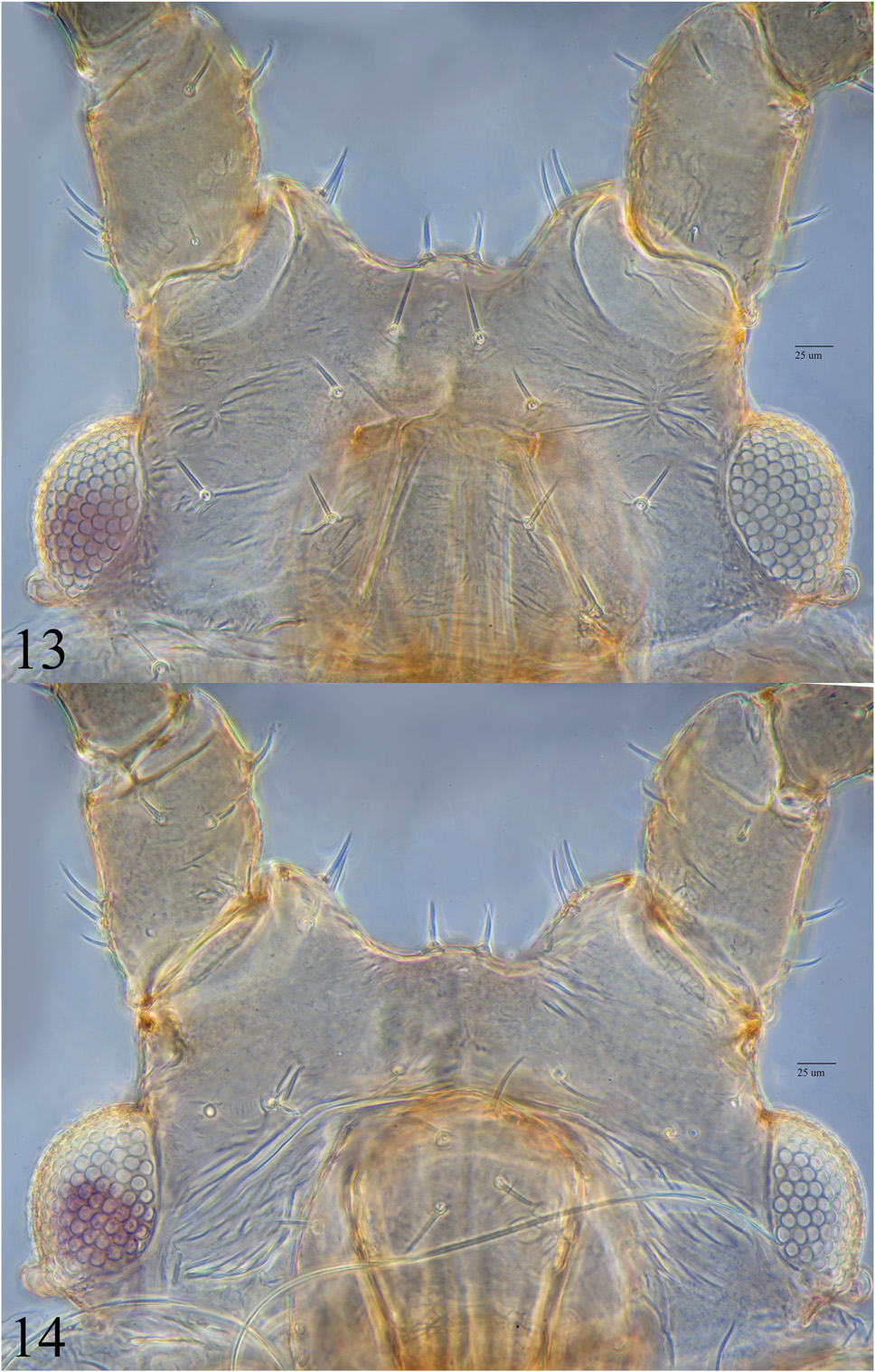
Recognition of M. glawatz from among the Macrosiphum -like species known to live on Potentilla can be accomplished using the key below, modified from Blackman and Eastop (2019). Among North American Macrosiphum , the two species most similar to M. glawatz are M. mentzeliae and Macrosiphum gaurae (Williams) . The species share several features including membranous abdominal tergum in apterae, moderate length antennae (see “Remarks” below under M. mentzeliae ), siphunculi, and cauda (for Macrosiphum ), generally pale body and appendages, long u.r.s., and the same u.r.s./metatarsal II ratios, and alatae with sensoria scattered over full length of a.s. III. However, accumulation of additional material of all three species revealed easy ways to separate them. The first author has studied and measured many specimens of M. gaurae from across North America, and a key feature of this species distinguishing it from M. glawatz and M. mentzeliae is that its siphunculi are almost always shorter than a.s. III (only one specimen has violated this rule, with siphunculus/a.s. 1.03), compared to siphunculi almost always longer than a.s. III in the latter two species (4 of 16 measured alatae of M. glawatz have siphunculi slightly shorter than a.s. III). Presence in apterae of spinules on the ventral surface of the head between antennae and mouthparts in M. glawatz ( Fig. 2 View Figs ) also distinguishes it from M. gaurae and M. mentzeliae (spinules absent or nearly so in both species). Similarly, absence of a cluster of robust spinules ventro-laterally on a.s. I separates M. glawatz from M. mentzeliae ( Fig. 14 View Figs ) and M. gaurae . A species that may be truly closely related is M. garyreed , which feeds on Geum triflorum living in the same dry forest habitat as M. glawatz and which looks similar in the field. These two species share features such as membranous abdominal tergum in apterae, moderate length antennae, siphunculi, and cauda (for Macrosiphum ), generally pale body and appendages, and long u.r.s., but can be easily separated by the following: u.r.s./metatarsal II ratio almost always smaller in M. glawatz (in apterae, 1.03– 1.24 in M. glawatz , 1.20–1.43 in M. garyreed ), spinules on the head ventrally between antennae and mouthparts in M. glawatz (usually absent in M. garyreed ), lateral and spinal tubercles common in M. garyreed , rare in M. glawatz , M. glawatz almost always with more sensoria on a.s. III (in apterae, 3–19 in M. glawatz , 1–10 in M. garyreed ; in alatae, 18-35 in M. glawatz , 15–16 in M. garyreed ).
Below is the section of the key from Blackman and Eastop (2019) for the aphids recorded on Potentilla edited to include M. glawatz , and to include M. valerianae in place of M. potentillicaulis and using their abbreviations and terminology. In constructing the M. potentillae aspects of the key, Blackman and Eastop (2019) seem to have previously used material of M. glawatz . Mariusz Kanturski (and later, Paul Brown) kindly provided a photograph of the two slides in the BMNH and mentioned by Blackman and Eastop (2019) under the discussion of M. potentillae . These specimens were collected by D. Hille Ris Lambers in the Warner Mountains of Lake County, Oregon, the same area in which most of our collections and observations were made. Therefore, the key below was reconstructed using the Oestlund material from UMN and the second author’s specimens of M. potentillae from Quebec. Also, we modified couplet 27 to reflect the range of variation found in our specimens of M. agrimoniellum and to add an important character of the alatae. Finally, our available material identified as Macrosiphum pallidum (Oestlund) does not conform to the separation in couplet 30; this is likely due to uncertainty around the range of variation and/ or species boundaries of what is called M. pallidum across North America. Because we cannot improve this couplet, we have left it as is.
19. SIPH with a subapical zone of reticulation (at least 4-5 rows of closed, polygonal cells) ...................................................... 20
– SIPH without polygonal reticulation (sometimes with a few interconnected subapical striae) .................................... 31
20 SIPH dark and with a greatly inflated, balloon-like middle section ............................ .................... Rhopalosiphoninus latysiphon
– SIPH pale or dark, not inflated ............ 21
21. Dorsal abdomen with extensive light to dark brown sclerotisation. Ventral side of head ornamented with spicules from front of ANT tubercles to rear of head ............................ 22
– Dorsal abdomen without extensive sclerotisation. Head usually without spicules but if spicules present, not covering all of ventral side of head ............................. 24 22. Longest hairs on ANT III 0.6-1.4 X BD
III. Cauda triangular, less than 1.5 X
longer than its basal width in dorsal view
............................ Anthracosiphon hertae
– Longest hairs on ANT III about 0.5 X BD
III. ....................................................... 23
23. Antennae beyond ANT I and II black.
Cauda finger-like, at least 2 X longer than
its basal width in dorsal view (0.25-
0.32 mm) ....... Anthracosiphon crystleae
– Antennae light brown to brown. Cauda
triangular, about 1.5 X longer than its
basal width in dorsal view (about
0.22 mm) ........ Macrosiphum potentillae
24. Longest hairs on ANT III 0.3-0.5 X BD
III. ANT III with 1-4 very small rhinaria.
ANT tubercles rather low, and median
frontal tubercle distinctly developed ......
.................................... Sitobion fragariae
– Longest hairs on ANT III 0.6-1.3 X BD
III. ANT III with 1-35 rhinaria of var-
ious sizes. ANT tubercles well devel-
oped, median frontal tubercle undeveloped
................................................................ 25
25. SIPH pale or dusky only towards apices
......................................................... 26
– SIPH dark, or pale only at bases ....... 28
26. Ventral side of head with a band of spic-
ules between ANT and base of mouthparts
.............................. Macrosiphum glawatz
– Ventral side of head without spicules or
with only a few tiny spicules on each side
of midline between ANT tubercles and
base of mouthparts ............................. 27
27. R IV+V 1.1-1.3 X HT II. ANT III with (8-)
15-20 rhinaria extending over more than
0.6 of its length. (ANT III of al. with
rhinaria extending over full length) ....
.............. Macrosiphum agrimoniellum
– R IV+V 0.8-1.0 X HT II. ANT III with 1-
10 rhinaria confined to basal half. (ANT
III of al. with rhinaria extending over ~0.8
of its length).. Macrosiphum euphorbiae
28. SIPH wholly jet black, and rather
thick; 5-7 X as long as their width at
midlength. ANT III pale except at apex
................ Macrosiphum pachysiphon – SIPH wholly dark, or paler at base, and more
than 10X longer than their width at mid-
length. ANT III dark at least on sensoriated
part ......................................................... 29
29. Head, ANT I and ANT II as dark as SIPH,
which are wholly dark. ANT III with 4-25
rhinaria 77% with 11 or more. (ANT III of al.
with 31-71 rhinaria) ........ Macrosiphum rosae
– Head, ANT I and ANT II paler than darkest
part of SIPH, which may be paler at base.
ANT III with 2-10 rhinaria. (ANT III of al.
with 13-22 rhinaria) ............................... 30
30. SIPH 1.9-2.2 X cauda. R IV+V 0.8-0.95
X HT II ......... Macrosiphum valerianae
– SIPH 1.2-1.7 (-1.8) X cauda. R IV+V 0.9-
1.15 X HT II ......... Macrosiphum pallidum




| USNM |
USA, Washington D.C., National Museum of Natural History, [formerly, United States National Museum] |
| BMNH |
United Kingdom, London, The Natural History Museum [formerly British Museum (Natural History)] |
| CNC |
Canada, Ontario, Ottawa, Canadian National Collection of Insects |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |